


-
山苍子(Litsea cubeba (Lour.) Pers.)系樟科(Lauraceae)木姜子属(Litsea Lam.)落叶灌木或小乔木,其果实、根、茎、叶等均含有芳香油,是我国特有的香料植物资源之一[1]。山苍子具有生长快、结实早、耐贫瘠等特点,其精油在香料上可以合成紫罗兰酮类香料和日化香精等,医药上有祛风散寒、消肿止痛、平喘和抗过敏等功效,食用上可用于除膻去腥、提味增鲜,用途广泛[2]。
山苍子主要分布在中国、日本、印度以及印度尼西亚、马来西亚和越南等东南亚国家[3],我国山苍子资源广泛分布于长江以南区域。不同地理生境条件下的山苍子,在经过长期的自然选择和地理隔离后,形成了丰富的遗传变异[4]。本研究组前期试验表明,山苍子的果实产量、精油柠檬醛含量等主要育种目标与一些生长性状存在显著的相关关系[5-6],因此,在山苍子多点造林试验的初期,有必要对其生长表现进行测定并分析基因型与环境的互作关系,对参试家系进行早期适应性和稳定性评价,以期为后期选择经济性状优良的品种提供参考,从而尽早为不同地区选择优良家系,为山苍子遗传改良和良种选育奠定基础。
关于林木的生长适应性和遗传稳定性,已有多位学者在种源或家系选择研究时将其作为评价标准之一[7-8]。如张平安等[9]分析了油茶(Camellia oleifera Abel.)无性系在各地区的生长特性和稳定性,对参试无性系进行了分类;王军辉等[10]对桤木(Alnus cremastogyne Burk.)进行了多点试验研究,根据主效应值和稳定性参数分析结果,预估了各种源的适生地区。稳定性分析模型中,AMMI(Additive main effects and multiplicative interaction,又称为主效可加互作可乘模型)模型源于社会学和物理学领域,最开始由Gauch将之应用于多点试验的数据分析[11]。相较于方差分析、线性回归和主成分分析等区域试验的传统分析方差,AMMI模型通过从加性模型的互作项中进一步分离出若干个乘积项之和,能更准确地分析出基因型和环境的交互作用[12-13]。
目前,关于山苍子的研究大部分集中于山苍子生物学特性、精油提取加工和利用方面[14-16]。此外,山苍子的栽培造林方面也有一些研究[17-19]。然而,关于山苍子选择育种的研究报道却极少,仅有的3篇文献[6, 20-21]只是基于某一区域野生资源或单点试验林进行评价,没有考虑环境因素的效应,难以选择适应不同区域的良种。因此,进一步开展山苍子的良种选择和种质发掘具有重要的意义。本研究组于2009年开始山苍子的相关研究,陆续在多地营建了各种家系试验林,并已在天然种群的表型和经济性状变异[22-23]、单点试验林各家系生长和经济性状评价[6]等方面进行了相关报道。本研究以13个山苍子家系2年生多点试验林为对象,对其树高和地径进行测定,分析其遗传变异,并采用AMMI模型对山苍子家系进行适应性分析,以期为后续山苍子优良家系的选择及推广提供重要参考。
-
2014年8月采集参试的13个山苍子自由授粉家系成熟的种子(表 1),2015年春季于中国林业科学研究院亚热带林业研究所温室大棚播种育苗,2016年春季分别在福建省清流国有林场、湖北省京山县太子山林场、重庆市万州区林木种子站试验林场3个试验点造林(表 2)。试验林按照完全随机区组设计,6株小区,5次重复,株行距3 m×3 m,穴状整地,穴规格30 cm×30 cm×40 cm。2016年12月,林分停止生长时,利用卷尺和游标卡尺对3个地点的试验林的树高和地径进行全林每木调查。
表 1 参试家系及种源地
Table 1. Family code and provenances
家系Family 种源地Provenance 29 福建建瓯Jianou of Fujian GX2 广西百色Baise of Guangxi GX3 广西百色Baise of Guangxi GX4 广西百色Baise of Guangxi HS4 安徽黄山Huangshan of Anhui HS5 安徽黄山Huangshan of Anhui JY2 福建建阳Jianyang of Fujian SC1 浙江遂昌Suichang of Zhejiang SC2 浙江遂昌Suichang of Zhejiang SC3 浙江遂昌Suichang of Zhejiang SC6 浙江遂昌Suichang of Zhejiang SC7 浙江遂昌Suichang of Zhejiang SC9 浙江遂昌Suichang of Zhejiang 表 2 各试验点概况
Table 2. General situation of each test site
试验点Test site 经度E 纬度N 海拔Altitude/m 年平均气温Meanannual temperature/℃ 年平均日照Mean annual sunshine /h 年平均降水量Mean annual precipitation/mm 无霜期Frost-free period/d 土壤类型Soil type 土壤pH Soil pH 重庆万州Wanzhou of Chongqing 108°14′ 30°38′ 817.1 17.7 1 484.4 1 243.0 320~335 黄壤 6.2 湖北京山Jingshan of Hubei 112°52′ 30°56′ 124.6 16.3 1 970.5 1 179.0 223~243 黄壤 5.9 福建清流Qingliu of Fujian 116°49′ 26°11′ 438.7 18.2 1 583.4 1 853.5 256~300 黄壤 5.1 -
利用DPS16.05统计分析软件对数据进行AMMI模型及线性模型分析。
AMMI稳定性分析模型如下[24]:
$ Y_{i j k}=\mu+\alpha_{i}+\beta_{j}+\sum\limits_{r=1}^{n} \lambda_{r} \psi_{i} \sigma_{j r}+\rho_{i j}+\varepsilon_{i j k} $
式中:Yijk是基因型i在环境j的第k次重复的观测值;μ是总体均值;αi是第i个基因型与总体均值的离差(即基因型主效应);βj是第j个环境与总均值的离差(即环境主效应);λr为第r个交互效应主成分轴(IPCA)的特征值或奇异值,代表交互作用平方和中可由该轴解释的部分;ψir为第r轴的基因型特征向量值;σjr为第r轴的环境特征向量值;n为保留在模型中的主成分因子轴的总个数;ρij为提取n个IPCA轴后留下的残差(交互剩余项);εijk为实验误差。
线性回归分析模型[24]为:
$ Y_{i j}=\mu+g_{i}+e_{j}+g e_{i j}+\varepsilon_{i j} $
式中:Yij是第i个家系在j地点的性状观测值;μ为总体均值;gi为i家系基因型主效应;ej为j地点环境主效应;geij为家系与地点的交互作用;εij为随机误差。
基因型(家系)的相对稳定性参数参照吴为人[25]改进的方法进行计算:
$ D_{i}=\sqrt{\sum\limits_{i=1}^{n} \omega_{n} \gamma_{i n}^{2}} $
式中:n为提取的IPCA个数,ωn为权重系数,它表示每个IPCA所解释的平方和占全部IPCA解释的平方和的比例,γin2为第个i基因型在第n个IPCA上的得分。Di的大小度量了基因型的相对稳定性,Di值越小越稳定。
-
表型变异系数(PCV)和遗传变异系数(GCV)计算公式[26]:
$ P C V=\frac{S}{X} \times 100 \% $
$ G C V=\frac{\sigma}{X} \times 100 \% $
式中:S为表型标准差,σ为遗传标准差,X为性状群体均值。
家系遗传力:$h^{2}=1-1 / F$[27]。
-
Bartlett检验结果(表 3)显示:山苍子家系的树高和地径在各试验点的误差均方差异不显著。因此,对3个参试地点各家系的生长性状进行联合方差分析,结果(表 4)表明:树高在家系间差异显著(0.01<P<0.05),在地点间表现出差异极显著(P<0.01),而家系和地点的交互作用未达到显著水平;地径在家系间和地点间均差异极显著(P<0.01),在家系和地点交互作用上差异显著(0.01<P<0.05)。
表 3 Bartlett方差齐性检验
Table 3. Homogeneity test of variance by Bartlett
性状Traits 试验点Test site 卡方值Chi P 重庆万州Wanzhou of Chongqing 20.724 0.054 树高Height 湖北京山Jingshan of Hubei 10.385 0.582 福建清流Qingliu of Fujian 8.399 0.753 地径Grounddiameter 重庆万州Wanzhou of Chongqing 12.194 0.430 湖北京山Jingshan of Hubei 17.332 0.138 福建清流Qingliu of Fujian 20.605 0.056 表 4 3个试验点生长性状联合方差分析
Table 4. Joint analysis of variance for three test sites
生长性状Growth traits 变异来源Source of variation df SS MS F P 树高Height 家系Family 12 10 696.37 891.36 2.259* 0.016 地点Site 2 61 307.69 30 653.85 77.697** <0.01 交互作用Family×Site 24 8 145.70 339.40 0.860 0.651 误差Error 78 30 773.28 394.53 地径Ground diameter 家系Family 12 188.42 15.70 3.302** <0.01 地点Site 2 820.14 410.07 86.247** <0.01 交互作用Family×Site 24 202.11 8.42 1.771* 0.031 误差Error 78 370.86 4.75 注:*差异显著(0.01<P<0.05),**差异极显著(P<0.01),下同。
Note: * Significant difference (0.01<P<0.05), ** extremely significant difference (P<0.01), the same below.地径的家系和地点交互作用达到显著水平(0.01<P<0.05),而且家系、地点以及家系和地点交互作用的平方和分别占总平方和的11.91%、51.86%和12.78%,可见家系和地点交互作用对地径的影响比家系单一作用大,有必要对交互作用进行进一步的分析。因此,确定利用地径进行后续的稳定性分析。
-
不同试验点13个山苍子家系的树高和地径变异参数见表 5。总体看来,地径的表型变异系数(PCV)和遗传变异系数(GCV)都大于树高,树高PCV最大的地点为重庆万州,地径PCV最大的地点为福建清流,而湖北京山试点的树高GCV和地径GCV均最大。2个生长性状在3个地点间具有相同的趋势,福建清流试点的树高和地径最大,湖北京山试点次之,重庆万州试点表现最差。湖北京山试点树高的家系遗传力较大,为0.578,重庆万州和福建清流试点树高的家系遗传力则较小(0.018和0.003);福建清流试点地径的家系遗传力最小(0.455),重庆万州试点地径的家系遗传力最大(0.806)。
表 5 各试验点生长性状变异参数
Table 5. Variation parameters of growth characters of each test site
性状Traits 试验点Test site 均值Mean 变幅Range of variation PCV/% GCV/% 家系遗传力Family heritability 树高Height/cm 重庆万州Wanzhou of Chongqing 64.04 20.00~100.50 23.84 1.97 0.018 湖北京山Jingshan of Hubei 96.12 54.50~147.33 21.30 11.64 0.578 福建清流Qingliu of Fujian 120.58 70.00~175.00 20.98 0.62 0.003 地径Ground diameter /mm 重庆万州Wanzhou of Chongqing 5.56 3.09~8.62 24.17 18.79 0.806 湖北京山Jingshan of Hubei 8.44 4.11~13.52 28.60 20.35 0.763 福建清流Qingliu of Fujian 12.03 5.40~18.95 29.27 13.31 0.455 -
鉴于地径的家系和地点交互作用表现出显著水平(0.01<P<0.05),对地径进行了线性回归模型和AMMI模型分析(表 6)。依据线性回归模型分析结果,联合回归仅能解释交互作用的13.39%,信息遗漏程度极高,因此,普通线性回归模型不适于对参试家系进行交互作用的分析。从AMMI模型的分析结果(表 6)可看出:仅第1项主成分分量就能解释交互作用的66.22%,且达显著性水平(0.01<P<0.05),表明应用AMMI解释参试家系与地点间的交互效应是可行的。进一步统计显示,2项主成分分量可以解释全部的互作项总平方和,具有极高的准确性。
表 6 地径线性模型和AMMI模型分析
Table 6. Analysis of variance, linear model and AMMI model of ground diameter
分析方法Method 变异来源Source of variation df SS SS% MS F P 线性回归Linear regression 联合回归Joint regression 1 27.07 13.39 27.07 5.69* 0.02 基因回归Gene regression 11 70.40 34.83 6.40 1.35 0.22 环境回归Environment regression 1 0.38 0.19 0.38 0.08 0.78 残差Residual 11 104.26 9.48 1.99* 0.04 误差Error 78 370.86 4.75 AMMI模型AMMI model IPC1 13 133.83 66.22 10.29 2.17* 0.02 IPC2 11 68.27 33.78 6.21 1.31 0.24 误差Error 78 370.86 4.75 注:表中SS%表示各项平方和占地径的家系和地点交互作用平方和的比例。
Note:SS% in the table shows the proportion of the sum of square to the tatal sun of square of Family×Site interaction. -
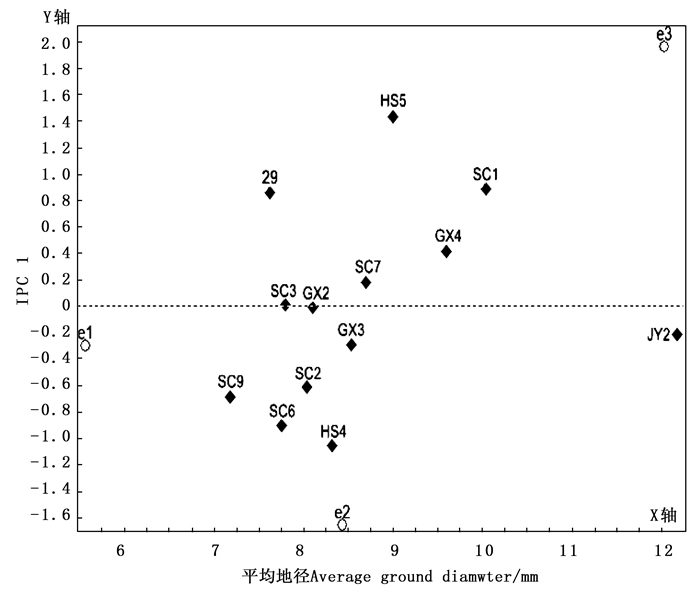
图 1给出了以平均地径为x轴,以家系和地点交互效应的主成分分量IPC1值为y轴的AMMI 1双标图。AMMI 1双标图中,越远离y轴的家系地径越大,越靠近x轴的家系越稳定;家系图标越接近某些试点,即该家系对这些试点具有特殊适应性。由图 1可见:在横轴方向上试点的分散程度大于家系的分散程度,说明试点的变异大于家系的变异,即同一家系在不同试点的地径表现差异,比同一试点不同家系间的地径表现差异大。在横轴方向上,家系JY2的地径均值最大,家系SC9的地径均值最小;福建清流试点的地径均值最大,湖北京山试点居中,重庆万州试点最小。在纵轴方向上,SC3和GX2 2个家系地径的IPC1值最小,其次是JY2、GX3和SC7 3个家系,说明这5个家系与地点的互作效应较小,受环境影响小,稳定性较强;相应的HS5和HS4等家系地径的IPC1值较大,稳定性较差。
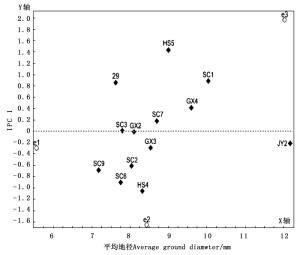
图 1 AMMI 1双标图
Figure 1. AMMI 1 Biplot
在以IPC1值和IPC2值为横纵坐标的AMMI 2双标图中(图 2),IPC1和IPC2可共同解释全部的交互作用。AMMI 2双标图中,家系到原点的距离越近,交互作用越小,家系稳定性越高。可以看出,SC3、SC7和GX2 3个家系距离原点最近,稳定性较好,而家系HS5的稳定性最低。
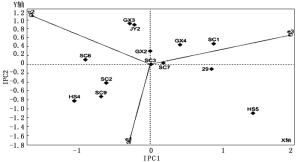
图 2 AMMI 2双标图
Figure 2. AMMI 1Biplot
双标图的优点是可以直观地了解山苍子各家系的地径表现以及其和地点间的交互作用,缺点则是不能进行定量描述,因此,对各家系的稳定性参数Dg进行了计算。从表 7可看出:山苍子各家系地径的稳定性排名为:SC3>SC7>GX2>GX4>JY2>SC2>GX3>29>SC9>SC6>SC1>HS4>HS5。
表 7 7各家系稳定性参数
Table 7. Stability parameter of each family
家系Family 地径均值Mean /mm 离差Deviation IPC1 IPC2 Dg 位次Precedence 29 7.621 7 -1.056 8 0.852 1 -0.103 8 0.696 0 8 GX2 8.097 4 -0.581 1 -0.005 8 0.296 6 0.172 5 3 GX3 8.535 0 -0.143 5 -0.288 9 0.925 0 0.586 8 7 GX4 9.591 1 0.912 6 0.414 8 0.444 3 0.425 0 4 HS4 8.311 1 -0.367 4 -1.056 3 -0.821 8 0.983 4 12 HS5 9.003 0 0.324 5 1.430 5 -1.100 3 1.328 2 13 JY2 12.173 3 3.494 9 -0.219 3 0.895 2 0.550 0 5 SC1 10.042 0 1.363 6 0.886 3 0.468 5 0.770 9 11 SC2 8.028 7 -0.649 8 -0.615 4 -0.414 8 0.555 8 6 SC3 7.799 6 -0.878 8 0.011 2 -0.001 5 0.009 2 1 SC6 7.746 7 -0.931 8 -0.906 8 0.107 6 0.740 6 10 SC7 8.692 2 0.013 7 0.181 3 0.031 4 0.148 7 2 SC9 7.178 3 -1.500 1 -0.683 9 -0.726 2 0.698 5 9 对图 1、2和表 7进行综合分析,参试的13个山苍子家系中,地径表现好且稳定性高的家系为JY2和GX4,地径表现好但稳定性差的家系为SC1和HS5,地径表现差但稳定性高的家系为SC3和GX2,地径表现和稳定性都较差的家系为SC2、HS4和GX3。
-
从图 1可以看出:福建清流试点与HS5、29、SC1、GX4和SC7 5个家系位于y轴的同一侧,福建清流试点对这些家系具有正向积极的交互作用,即这些家系在福建清流试点地径表现较好;相应的重庆万州试点和湖北京山试点对HS4、SC6、SC9、SC2、GX3和JY2 6个家系具有正向积极的交互作用,说明这些家系在这2个试验点地径表现较好。
AMMI 2双标图中,家系在地点与原点连线上的投影点到原点的距离表示该家系在此地点的交互作用的大小,距离越长,交互作用越大,若投影点落在连线上则交互作用为正向,若落在连线的反向延长线上,则交互作用为负向[28]。由图 2可以看出:家系HS5和家系HS4分别在湖北京山试点和福建清流试点的反向延长线上有较长的反向投影,说明湖北京山试点对家系HS5、福建清流试点对家系HS4有较强的负向交互作用,家系HS5不适宜在湖北京山试点种植,家系HS4不适宜在福建清流试点种植。
-
3个试验点联合方差分析表明,参试山苍子家系的树高和地径在家系和地点间的差异均达到显著(0.01 < P<0.05)或极显著水平(P<0.01);家系和地点的交互作用树高未达到显著水平(0.01 < P<0.05),地径则达到显著水平(0.01 < P<0.05),且交互作用的影响大于家系的影响,这为山苍子的稳定性分析提供了可信的遗传背景,因此,确定利用地径进行后续的稳定性分析。家系JY2在3个试验点生长表现均较好,具有普遍适应性,而其它家系则对某些试点具有特殊适应性。与此类似,颜昌兰等[30]在对青稞(Hordeum vulgare Linn. var. nudum Hook.f.)的品种稳定性和适应性研究中,也通过AMMI模型分析筛选出了具有普遍适应性和特殊适应性的家系。家系29在福建清流试点生长表现较好,而在湖北京山和重庆万州试点的树高和地径则排名较靠后,这可能是由于家系29的种源地福建建瓯与福建清流试点距离较近,立地环境较相似。
本研究中,山苍子地径的家系和地点交互作用效应大于家系效应,所以家系选择时应考虑存在的互作效应。与本研究不同,赵兴堂等[26]在水曲柳(Fraxinus mandshurica Rupr.)种源选择时,由于各性状种源效应值均大于种源与环境互作效应值,故选择时以种源效应为准。在家系和地点交互作用显著的基础上,对参试家系的地径表现进行AMMI模型分析,2种双标图和稳定性参数各自所呈现出来的信息都有局限性,结果也会有出入,如家系29在AMMI 1双标图中稳定性较差,稳定性参数排名也较靠后,而在AMMI 2双标图中则稳定性较好。可见,不同的分析手段虽然各有其优势,但由于模型不同也势必造成结果有细微差异。本研究综合AMMI双标图和稳定性参数对各家系进行评定,分别选出了地径表现好且稳定性高的家系JY2和GX4,地径表现好但稳定性差的家系SC1和HS5,地径表现差但稳定性高的家系SC3和GX2,以及地径表现和稳定性均较差的家系SC2、HS4和GX3。
由于育种目标不同,对于后续山苍子的推广方向也应有所侧重。对于生长良好且稳定性高的家系如JY2和GX4,可在多个地区造林,并重点关注其经济性状表现;对于生长良好但稳定性不高的家系如SC1和HS5,建议在其生长适宜区域造林,如和福建清流生境条件类似的地区;而对于生长较差但稳定性高的家系,其生长适应性强,对不良环境抵御能力高,例如家系SC3生长表现较差,但稳定性极高,建议在环境相对恶劣的地区造林。
限于林木生长周期较长,现阶段试验林仍处于幼林时期,只能对其生长性状进行初步评价和稳定性分析,为后续优良品种的多区域推广种植提供一定参考。山苍子为香料植物,良种选育应以经济指标为主要选择方向,下阶段需要对其产量和含油率等经济性状进行测定和分析,从而对山苍子优良家系进行综合评价和选择。
-
(1) 山苍子家系的树高和地径在家系和地点效应上均达显著(0.01 < P<0.05)或极显著(P<0.01)水平;3个试验点树高和地径的表型变异系数为20.98%~29.27%,遗传变异系数为0.62%~20.35%,其中,地径的表型和遗传变异系数普遍大于树高;3个试验点地径的家系遗传力(0.455~0.806)均大于树高的家系遗传力(0.003~0.578),树高受低等到中等强度的遗传控制,地径受中等到高等强度的遗传控制;山苍子各家系的树高和地径在福建清流试点生长表现最好,湖北京山试点居次,重庆万州试点最差。
(2) 参试山苍子家系中,地径表现好且稳定性高的家系为JY2和GX4;地径表现好但稳定性差的家系为SC1和HS5;地径表现差但稳定性高的家系为SC3和GX2;地径表现和稳定性都较差的家系为SC2、HS4和GX3。
山苍子家系幼林生长性状遗传变异及稳定性分析
Analysis of the Genetic Variation and Stability of Growth Traits of Litsea cubeba Young Plantation
-
摘要:
目的 对山苍子不同家系幼林生长性状进行遗传变异及稳定性分析,为山苍子优良家系选择及推广提供参考。 方法 以13个半同胞山苍子家系为材料,采用随机区组设计,分别在重庆万州、湖北京山和福建清流3个试验点营建家系试验林。对23年生山苍子幼林的树高和地径性状进行测定和多点联合分析,并采用AMMI模型对山苍子各家系的生长稳定性进行评价。 结果 (1)山苍子家系的树高和地径在家系和地点效应上均达到显著(0.01 < P < 0.05)或极显著(P < 0.01)水平,地径的家系和地点交互作用达到显著水平(0.01 < P < 0.05);3个试验点树高和地径的表型变异系数为20.98%~29.27%,遗传变异系数为0.62%~20.35%,其中,地径的表型和遗传变异系数普遍大于树高;树高的家系遗传力为0.003~0.578,地径的家系遗传力为0.455~0.806。(2)综合双标图和稳定性参数得出:家系JY2和GX4地径表现较好且稳定性较高。 结论 山苍子家系的树高和地径在家系和地点间的差异均达到极显著水平(P < 0.01),存在丰富的遗传变异。在家系水平上,树高受低等到中等强度的遗传控制,地径受中等到高等强度的遗传控制。各家系在福建清流试点生长表现最好,湖北京山试点居次,重庆万州试点最差,其中家系JY2和GX4在3个试验点地径均表现较好且稳定性较高。 Abstract:Objective To provide reference for the follow-up family selection and variety extension by genetic variation analysis and stability evaluation of the growth traits of Litsea cubeba families. Method 13 L. cubeba families were used to establish pedigree test forests in Wanzhou of Chongqing Municipality, Jingshan of Hubei Province and Qingliu of Fujian Province, which were designed in randomized complete blocks. The height and ground diameter were measured and analyzed when the forests were 2-years-old, then the growth performance and stability of different L. cubeba families were evaluated by AMMI model. Result (1) Both the family and site effect of height and ground diameter were significant(0.01 < P < 0.05) or highly significant (P < 0.01), the interaction effect of family and site of ground diameter was significant (0.01 < P < 0.05); the phenotypic variance coefficient of height and ground diameter ranged from 20.98% to 29.27%; the genetic variation coefficient of which ranged from 0.62% to 20.35%; and both the phenotypic and genetic variance coefficient of ground diameter were generally larger than that of height; the family heritability of tree height ranged from 0.003 to 0.578, and the family heritability of ground diameter ranged from 0.455 to 0.806. (2) Both the families JY2 and GX4 performed well in ground diameter and stability. Conclusion The family and site effect of height and ground diameter are both significant(0.01 < P < 0.05) or highly significant (P < 0.01), there is rich genetic variation. In the family level, the height of tree is genetically controlled by low to medium intensity, and the ground diameter is genetically controlled by medium to high intensity. The growth of different families performs best in Qingliu, followed by Jingshan, and the worst in Wanzhou. The families JY2 and GX4 show good ground diameter performance and high stability. -
Key words:
- Litsea cubeba
- / family
- / multipoint test
- / growth traits
- / genetic variation
- / stability
-
表 1 参试家系及种源地
Table 1. Family code and provenances
家系Family 种源地Provenance 29 福建建瓯Jianou of Fujian GX2 广西百色Baise of Guangxi GX3 广西百色Baise of Guangxi GX4 广西百色Baise of Guangxi HS4 安徽黄山Huangshan of Anhui HS5 安徽黄山Huangshan of Anhui JY2 福建建阳Jianyang of Fujian SC1 浙江遂昌Suichang of Zhejiang SC2 浙江遂昌Suichang of Zhejiang SC3 浙江遂昌Suichang of Zhejiang SC6 浙江遂昌Suichang of Zhejiang SC7 浙江遂昌Suichang of Zhejiang SC9 浙江遂昌Suichang of Zhejiang  下载: 导出CSV
下载: 导出CSV
表 2 各试验点概况
Table 2. General situation of each test site
试验点Test site 经度E 纬度N 海拔Altitude/m 年平均气温Meanannual temperature/℃ 年平均日照Mean annual sunshine /h 年平均降水量Mean annual precipitation/mm 无霜期Frost-free period/d 土壤类型Soil type 土壤pH Soil pH 重庆万州Wanzhou of Chongqing 108°14′ 30°38′ 817.1 17.7 1 484.4 1 243.0 320~335 黄壤 6.2 湖北京山Jingshan of Hubei 112°52′ 30°56′ 124.6 16.3 1 970.5 1 179.0 223~243 黄壤 5.9 福建清流Qingliu of Fujian 116°49′ 26°11′ 438.7 18.2 1 583.4 1 853.5 256~300 黄壤 5.1
下载: 导出CSV
表 3 Bartlett方差齐性检验
Table 3. Homogeneity test of variance by Bartlett
性状Traits 试验点Test site 卡方值Chi P 重庆万州Wanzhou of Chongqing 20.724 0.054 树高Height 湖北京山Jingshan of Hubei 10.385 0.582 福建清流Qingliu of Fujian 8.399 0.753 地径Grounddiameter 重庆万州Wanzhou of Chongqing 12.194 0.430 湖北京山Jingshan of Hubei 17.332 0.138 福建清流Qingliu of Fujian 20.605 0.056
下载: 导出CSV
表 4 3个试验点生长性状联合方差分析
Table 4. Joint analysis of variance for three test sites
生长性状Growth traits 变异来源Source of variation df SS MS F P 树高Height 家系Family 12 10 696.37 891.36 2.259* 0.016 地点Site 2 61 307.69 30 653.85 77.697** <0.01 交互作用Family×Site 24 8 145.70 339.40 0.860 0.651 误差Error 78 30 773.28 394.53 地径Ground diameter 家系Family 12 188.42 15.70 3.302** <0.01 地点Site 2 820.14 410.07 86.247** <0.01 交互作用Family×Site 24 202.11 8.42 1.771* 0.031 误差Error 78 370.86 4.75 注:*差异显著(0.01<P<0.05),**差异极显著(P<0.01),下同。
Note: * Significant difference (0.01<P<0.05), ** extremely significant difference (P<0.01), the same below.
下载: 导出CSV
表 5 各试验点生长性状变异参数
Table 5. Variation parameters of growth characters of each test site
性状Traits 试验点Test site 均值Mean 变幅Range of variation PCV/% GCV/% 家系遗传力Family heritability 树高Height/cm 重庆万州Wanzhou of Chongqing 64.04 20.00~100.50 23.84 1.97 0.018 湖北京山Jingshan of Hubei 96.12 54.50~147.33 21.30 11.64 0.578 福建清流Qingliu of Fujian 120.58 70.00~175.00 20.98 0.62 0.003 地径Ground diameter /mm 重庆万州Wanzhou of Chongqing 5.56 3.09~8.62 24.17 18.79 0.806 湖北京山Jingshan of Hubei 8.44 4.11~13.52 28.60 20.35 0.763 福建清流Qingliu of Fujian 12.03 5.40~18.95 29.27 13.31 0.455
下载: 导出CSV
表 6 地径线性模型和AMMI模型分析
Table 6. Analysis of variance, linear model and AMMI model of ground diameter
分析方法Method 变异来源Source of variation df SS SS% MS F P 线性回归Linear regression 联合回归Joint regression 1 27.07 13.39 27.07 5.69* 0.02 基因回归Gene regression 11 70.40 34.83 6.40 1.35 0.22 环境回归Environment regression 1 0.38 0.19 0.38 0.08 0.78 残差Residual 11 104.26 9.48 1.99* 0.04 误差Error 78 370.86 4.75 AMMI模型AMMI model IPC1 13 133.83 66.22 10.29 2.17* 0.02 IPC2 11 68.27 33.78 6.21 1.31 0.24 误差Error 78 370.86 4.75 注:表中SS%表示各项平方和占地径的家系和地点交互作用平方和的比例。
Note:SS% in the table shows the proportion of the sum of square to the tatal sun of square of Family×Site interaction.
下载: 导出CSV
表 7 7各家系稳定性参数
Table 7. Stability parameter of each family
家系Family 地径均值Mean /mm 离差Deviation IPC1 IPC2 Dg 位次Precedence 29 7.621 7 -1.056 8 0.852 1 -0.103 8 0.696 0 8 GX2 8.097 4 -0.581 1 -0.005 8 0.296 6 0.172 5 3 GX3 8.535 0 -0.143 5 -0.288 9 0.925 0 0.586 8 7 GX4 9.591 1 0.912 6 0.414 8 0.444 3 0.425 0 4 HS4 8.311 1 -0.367 4 -1.056 3 -0.821 8 0.983 4 12 HS5 9.003 0 0.324 5 1.430 5 -1.100 3 1.328 2 13 JY2 12.173 3 3.494 9 -0.219 3 0.895 2 0.550 0 5 SC1 10.042 0 1.363 6 0.886 3 0.468 5 0.770 9 11 SC2 8.028 7 -0.649 8 -0.615 4 -0.414 8 0.555 8 6 SC3 7.799 6 -0.878 8 0.011 2 -0.001 5 0.009 2 1 SC6 7.746 7 -0.931 8 -0.906 8 0.107 6 0.740 6 10 SC7 8.692 2 0.013 7 0.181 3 0.031 4 0.148 7 2 SC9 7.178 3 -1.500 1 -0.683 9 -0.726 2 0.698 5 9
下载: 导出CSV
-
[1] 方学军.山苍子油的功能及应用[J].湖南林业科技, 2007, 34(3):82-84. doi: 10.3969/j.issn.1003-5710.2007.03.032 [2] 崔永忠, 廖声熙, 崔凯, 等.贵州山区山苍子苗年生长规律[J].林业科学研究, 2013, 26(4):501-505. doi: 10.3969/j.issn.1001-1498.2013.04.017 [3] 王旭, 杨关锋.我国山苍子开发利用的现状与发展对策[J].经济林研究, 2010, 28(3):136-139. doi: 10.3969/j.issn.1003-8981.2010.03.028 [4] 田胜平, 汪阳东, 陈益存, 等.不同居群山苍子果实精油和柠檬醛含量及其与地理-气候因子的相关性[J].植物资源与环境学报, 2012, 21(3):57-62. doi: 10.3969/j.issn.1674-7895.2012.03.009 [5] 李红盛, 汪阳东, 陈益存, 等.山苍子生长经济性状遗传变异分析及优良家系选择[J].经济林研究, 2017, 35(4):64-71. [6] Gao M, Chen Y, Wang Y. Evaluation of the yields and chemical compositions of the essential oils of different Litsea cubeba varieties[J]. Journal of Essential Oil Bearing Plants, 2016, 19(8):1888-1902. doi: 10.1080/0972060X.2016.1252695 [7] 沈熙环.油松、华北落叶松良种选育实践与理论[M].北京:科学出版社, 2015:56-58. [8] 徐化成.油松地理变异与种源选择[M].北京:中国林业出版社, 1992:112-118. [9] 张平安, 孙凡, 姚小华, 等.油茶高产无性系生长特性及稳定性分析[J].西南大学学报:自然科学版, 2011, 33(4):12-17. [10] 王军辉, 顾万春, 李斌, 等.桤木优良种源/家系的选择研究——生长的适应性和遗传稳定性分析[J].林业科学, 2000, 36(3):59-66. doi: 10.3321/j.issn:1001-7488.2000.03.010 [11] 张泽, 鲁成.基于AMMI模型的品种稳定性分析[J].作物学报, 1998, 24(3):304-309. doi: 10.3321/j.issn:0496-3490.1998.03.007 [12] Zobel R W, Wright M J, Gauch H G. Statistical analysis of a yield trial[J]. Agronomy Journal, 1988, 80(3):388-393. doi: 10.2134/agronj1988.00021962008000030002x [13] 刘丽华, 胡远富, 陈乔, 等.利用AMMI模型分析寒地水稻3个品质性状的基因型与环境互作[J].作物学报, 2013, 39(10):1849-1855. [14] 张德权, 吕飞杰, 台建祥.超临界CO2流体技术萃取山苍子油的研究[J].食品与发酵工业, 2000, 26(2):54-57. doi: 10.3321/j.issn:0253-990X.2000.02.014 [15] Yang Y, Jiang J, Qimei L, et al. The fungicidal terpenoids and essential oil from Litsea cubeba in Tibet[J]. Molecules, 2010, 15(10):7075-7082. doi: 10.3390/molecules15107075 [16] Hu L, Wang Y, Du M, et al. Characterization of the volatiles and active components in ethanol extracts of fruits of Litsea cubeba (Lour.) by gas chromatography-mass spectrometry (GC-MS) and gas chromatography-olfactometry (GC-O)[J]. Journal of Medicinal Plants Research, 2011, 5(14):3298-3303. [17] 吴松成, 樊光毅, 谢宗仁.山苍子的开发利用及栽培技术[J].江西林业科技, 2003(1):15-16. doi: 10.3969/j.issn.1006-2505.2003.01.008 [18] 朱才熙.山苍子的生长特性与栽培技术[J].云南科技管理, 2000, 13(3):63. [19] 张国庆, 詹克敏.山苍子栽培与加工[J].安徽林业科技, 2003(1):16. [20] 岳季林, 徐文彬, 罗运春.贵州省山苍子品种(类型)及良种选择初步研究[J].经济林研究, 1997, 15(3):9-11. [21] 徐佑明, 唐忠平, 周仁飞.山苍子优良种质资源选择及丰产栽培技术[J].衡阳师范学院学报, 2016, 37(6):112-114. doi: 10.3969/j.issn.1673-0313.2016.06.022 [22] 田胜平, 汪阳东, 陈益存, 等.山苍子天然种群叶片和种实性状的表型多样性[J].生态学杂志, 2012, 31(7):1665-1672. [23] Si L, Chen Y, Han X, et al. Chemical composition of essential oils of Litsea cubeba harvested from its distribution areas in China[J]. Molecules, 2012, 17(6):7057-7066. doi: 10.3390/molecules17067057 [24] 唐启义. DPS数据处理系统[M].北京:科学出版社, 2013:1276-1318. [25] 吴为人.对基于AMMI模型的品种稳定性分析方法的一点改进[J].遗传, 2000, 22(1):31-65. [26] 赵兴堂, 夏德安, 曾凡锁, 等.水曲柳生长性状种源与地点互作及优良种源选择[J].林业科学, 2015, 51(3):140-147. [27] 顾万春, 刘德安, 田玉林.白榆种源与家系的选种研究[J].林业科学, 1987, 23(4):415-424. [28] 袁爱梅, 张敏, 陈惠敏, 等.利用AMMI模型分析冬小麦品种区试数据的稳定性[J].种子, 2005, 24(5):59-61. doi: 10.3969/j.issn.1001-4705.2005.05.017 [29] 严威凯.双标图分析在农作物品种多点试验中的应用[J].作物学报, 2010, 36(11):1805-1819. [30] 颜昌兰, 白文琴, 郭超, 等.青稞品种稳定性及适应性的AMMI模型分析[J].干旱地区农业研究, 2016, 34(2):157-162. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 4006
- HTML全文浏览量: 1627
- PDF下载量: 367
- 被引次数: 0
