


-
我国油茶(Camellia oleifera Abel)主产区大多为典型的亚热带高温气候,土壤长期的脱硅富铝化作用致使钾素流失较多,钾成为了油茶林地的土壤养分限制因子之一[1-2]。我国从60年代起开始油茶施肥试验,研究领域多集中于氮、磷、钾的施肥量和施肥方法对油茶地上部分的影响[1, 3],对油茶根系影响的研究并不多见。微根管技术自20世纪90年代以来,已较广泛地运用于农林业[4-6]。张永清等[7]研究表明,低钾胁迫下小麦的根尖数、总根长、总表面积均明显降低;张志勇等[8]在缺钾对棉花幼苗根系生长的相关研究中表明,缺钾显著抑制了根系伸长;Shin[9]等认为缺钾不会抑制根系的伸长生长,但显著降低了侧根数和侧根分布密度。目前有关施钾对油茶细根时空分布动态的相关研究还较少。本研究以初果期的高产油茶林为试验材料,在测土的基础上,进行了不同钾肥用量试验,对该油茶林细根的形态指标及净生长速率进行了探讨。
-
试验地位于江西省宜春市袁州区西村镇,属典型亚热带季风气候,年降水量1 595.8 mm,年平均温度16.4℃。试验地为红壤,缓坡,是油茶的适生区和主产区。土壤平均养分采用ASI法测定,其中土壤pH为3.51,有机质含量12.65 g·kg-1,硝态氮含量4.37 mg·kg-1,铵态氮含量69.81 mg·kg-1,速效磷含量7.02 mg·kg-1,速效钾含量93.73 mg·kg-1。供试材料为2011年造林的高产油茶林,平均株高、地径、冠幅依次为:186 cm、4.8 cm、139 cm。
2015年11月在油茶林地随机选择长势中等的油茶树,参照Johnson [10]等介绍的方法,于其根部按同一方向与地面呈45°角埋设微根管并编号,共埋设27根微根管。微根管外径5.5 cm,内径5.0 cm,长90 cm,露出地面33 cm,垂直深度达40 cm左右。试验采用完全随机化设计,在其它养分水平和管理措施一致的前提下设置了3个钾素水平(K0、K1、K2)处理,9次重复。3个处理的有效养分含量依次为N92P48K0、N92P48K135、N92P48K270。其中,P肥(钙镁磷)每年冬季一次施入,K肥(氯化钾)于每年5月和12月分两次施入,N肥每年3月和5月分两次施入。
-
为减少管的安装造成对根生长的影响,使管与土壤达到良好的接触,在微根管安装半年后进行数据采集[11-13]。从2016年6月26日开始每月采集一次图像,到2017年6月止, 期间共采集图像12次(2017年1月冬季低温,油茶处于休眠期状态,未采集数据)。采用BTC图像采集系统(美国Bartz技术公司生产)采集图像,观测窗面积为1.8 cm×1.4 cm,每个微根管收集40张图片,可观测020 cm与2040 cm土层的油茶细根。用WinRHIZOTron图像分析软件对所采集的图像进行处理,以获取细根长度、平均直径、表面积等数据。依据微根管号、图像采集时间、观测框位置、细根编号等建立细根数据库[14-16]。
本研究所采用的细根指标为总根长(TRL)、总根尖数(TRT)、总表面积(TSA)、平均直径(ARD)、平均根长密度(ARLD)以及净生长速率(ARLDNGR)。以单位土壤体积的根长密度(RLD)作为研究细根生长动态的基本参数,其计算公式如下:
$ RLD = RL/A \times DOF $
其中,RL为观测窗中观测到的细根根长;A为观测窗面积;DOF为田间观测深度或田间深度,一般为0.20.3 cm,本研究中取0.2 cm[17-18]。
细根净生长速率(RLDNGR)为单位时间的细根根长密度净增加量,按下式计算:
$ RL{D_{NGR}} = (RL{D_{n + 1}} - RL{D_n})/T $
RLDn+1与RLDn分别表示第n+1次与第n次观测到的细根根长密度,T为相邻两次观测的间隔天数。
-
用Microsoft Excel 2013整理获得的细根数据,采用IBM Statistcis SPSS 20.0软件对数据进行统计分析,用单因素方差分析对不同钾素水平以及不同土层的油茶细根生长指标是否具有显著差异进行检验, 并对其进行重复测量方差分析,从而比较不同钾水平处理下油茶细根的生长差异及其空间分布规律。
-
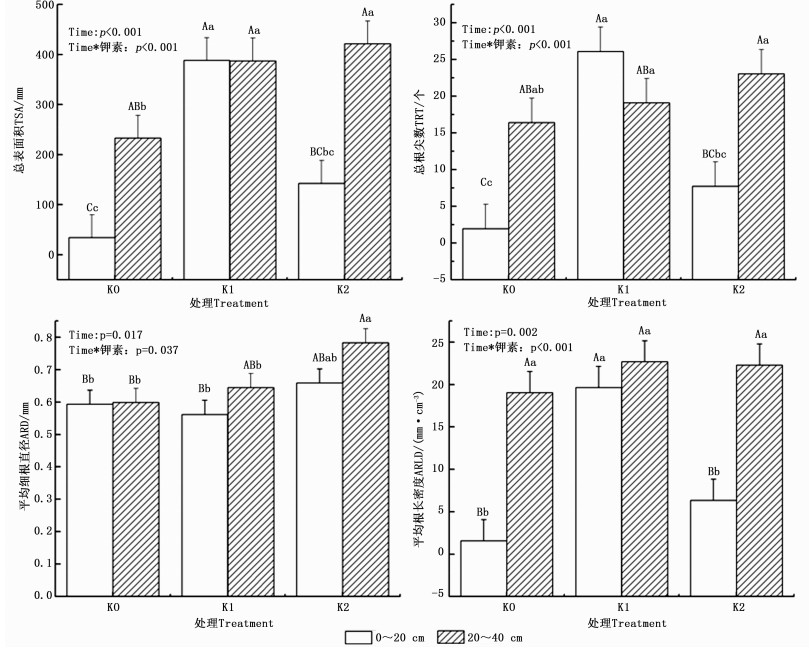
对2016年6月—2017年6月共12次观测得到的油茶细根形态指标数据进行分析(表 1)表明,TRL在不同观测时期存在显著差异(P < 0.05),时间与钾素处理间无显著交互作用;其它3个形态指标不同观测时期之间均存在极显著差异(P < 0.01),ARD与TRT分别存在时间与钾素处理的显著与极显著交互作用,而TSA无时间与钾素处理的交互作用;除TRT外,其它3个形态指标都在钾素水平间均存在显著差异。
表 1 油茶细根形态指标重复测量方差分析
Table 1. Repeated measurement variance analysis results of morphology indicators of Camellia oleifera fine root
变异来源
Source of variation平均细根直径ARD 总根尖数TRT 总根长TRL 总表面积TSA P F P F P F P F 时间Time 0.002 2.834 < 0.001 3.279 0.011 2.293 0.002 2.775 时间*钾素Time*Potassium 0.017 1.803 0.001 2.305 0.073 1.501 0.351 1.095 钾素Potassium 0.029 4.132 0.194 1.758 0.042 3.358 0.012 5.290 多重比较结果(表 2)表明,K1、K2处理下的ARD都显著大于K0,K1与K2间无显著差异;K1处理下的TRL显著大于K0,K2与K0差异不显著;就TSA而言,K1极显著大于K0,而K2、K0处理之间有显著差异;施钾肥有利于油茶细根总根长、总表面积以及平均直径的增加,基于油茶细根形态指标的重复测量方差分析,总体上以K1处理效果最佳。
表 2 油茶细根形态指标多重比较
Table 2. Multiple comparisons result of morphology indicators of Camellia oleifera fine root
钾素水平Potassium level 平均细根直径ARD/mm 总根尖数TRT/个 总根长TRL/ mm 总表面积TSA/ mm2 K0 0.576 6Ab 14.453 7Aa 118.874 5Ab 214.189 1Bb K1 0.688 6Aa 29.157 4Aa 261.478 7Aa 573.618 6Aa K2 0.668 7Aa 21.648 1Aa 183.487 1Aab 419.432 ABab -
重复测量方差分析结果(表 3)表明,油茶的ARLD在不同的观测时期内差异显著(P < 0.05),时间与钾素处理之间无显著交互作用,不同钾素水平下的ARLD存在显著差异。多重比较结果(表 4)表明:K1处理下的ARLD显著大于K0处理,K2处理下的ARLD虽然也大于K0处理,但两者差异不显著。
表 3 平均根长密度及细根净生长速率的重复测量方差分析
Table 3. Repeated measurement variance analysis results of ARLD and ARLDNGR
变异来源
Source of variation平均根长密度
ARLD净生长速率
ARLDNGRP F P F 时间Time 0.011 2.293 0.001 3.255 时间*钾素Time*Potassium 0.073 1.501 0.202 1.278 钾素Potassium 0.042 3.358 < 0.001 31.758 表 4 平均根长密度及细根净生长速率的多重比较
Table 4. Multiple comparisons result of ARLD and ARLDNGR
钾素水平
Potassium level平均根长密度
ARLD/ (mm·cm-3)净生长速率
ARLDNGR /(mm·cm-3·d-1)K0 5.896 6Ab -0.031 1Bb K1 12.970 2Aa 0.060 6Aa K2 9.101 5Aab 0.048 1Aa 从表 3可知,不同观测时期的ARLDNRG差异极显著(P < 0.05),时间与钾素处理之间无显著交互作用,不同钾素水平下的ARLDNRG存在极显著差异。多重比较结果(表 4)表明:K1、K2处理下的ARLDNRG显著高于K0处理,K1、K2处理间差异不显著。这说明施钾有利于提高油茶细根的净生产力。
-
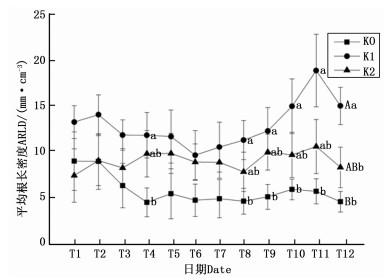
图 1所示为2016年6月—2017年6月不同钾水平下油茶细根形态指标的年生长动态。T1T12表示从第1次到第12次的观测日期。由图 1可知,油茶的TRL和TRT在T1T3(2016年6月—2016年8月)时期都呈现为先增加后减小的趋势,在T4T8时期(2016年9月—2017年2月)变化较为稳定;在T10T12(2017年4月—2017年6月)时期,油茶的TRL和TRT再次出现先增加后减小的趋势,并且K1处理下的TRL和TRT在T11(2017年5月)出现明显峰值。与不施钾肥的K0处理相比,在施了钾肥的K1与K2处理下,油茶细根的TRL在T4(2016年9月)、T8(2017年2月)、T9(2017年3月)、T10(2017年4月)表现为K1>K2>K0,且K0、K1间差异显著;在观测末期(2017年5月、6月),K1处理下的TRL和TRT极显著大于对照;与对照相比,K1处理下的TRT于T10、T11(2017年4月、5月)与T12(2017年6月)分别出现显著与极显著差异。
图 1 油茶细根形态指标的动态变化
Figure 1. Dynamic changes of morphology indicators of Camellia oleifera fine root
油茶的ARD在整个生育期内变化幅度较大,各处理都在T9(2017年3月)出现峰值,K0、K1、K2三个处理下的ARD峰值分别为0.631、0.750、0.788 mm;在T1、T6、T9、T12(2016年6月、2016年12月、2017年3月、2017年6月)时期,K2处理与K0处理的ARD之间差异显著,K1与K2处理无显著差异。就TSA而言,观测前期及观测末期各处理的变幅较大,观测中期的TSA变化较稳定;K2处理下的TSA于T9(2017年3月)先于K1处理出现峰值,而K0处理的TSA在2017年无明显峰值;在整个观测期内油茶的TSA都表现为K1>K2>K0,并且K0、K1处理在T3、T4以及T7T12时期(2016年8月、2016年9月以及2016年12月—2017年6月)出现显著或极显著差异,K1与K2处理间差异不显著。
-
从图 2可知,K0处理的ARLD在T2T4(2016年7月—2016年9月)时期急剧下降,之后变化平缓;这可能是由于油茶果实油脂转化期消耗掉地下根系过多养分而致根系大量死亡[19];而施了钾肥的K1与K2处理减缓了油茶根系的死亡,其ARLD在T2T4(2016年7月—2016年9月)时期表现为先减小后增大;在T6T11(2016年11月—2017年5月),K1处理下的ARLD持续大幅增大,并于T11(2017年5月)出现峰值。油茶的ARLD在首次观测时期表现为K1>K0>K2,而在T1(2016年6月)之后的观测期内都表现为K1>K2>K0,并且K1处理下的ARLD在T4和T8T11(2016年9月和2017年2月—2017年5月)时期显著大于K0处理,K2与对照无显著差异;K1处理的ARLD在T12(2017年6月)极显著大于K0处理,显著大于K2处理。

图 2 油茶细根平均根长密度的动态变化
Figure 2. Dynamic changes of ARLD of Camellia oleifera fine root
图 3所示为1年观测期内油茶细根净生长速率在不同钾水平下的的变化规律。油茶的ARLDNRG在整个生育期内波动幅度较大,相比K0处理而言,K1、K2处理下的油茶ARLDNRG变化较剧烈,尤其体现在DF时间段(2016年10月—2016年12月),其ARLDNRG剧烈下降至负值后又大幅上升到正值,说明施钾肥促进了油茶细根的周转。K1处理在IJ阶段(2017年3月—2017年5月)的ARLDNRG达到峰值,为0.2 mm·cm-3·d-1,说明K1处理下油茶细根在春季的生产量远大于死亡量。在整个观测期内,油茶的ARLDNRG总体表现出K1、K2处理大于K0处理,并且K2、K1处理分别在B、J(2016年7月—2016年8月、2017年4—5月)时间段显著高于K0处理,分别在C、A时间段(2016年6月—2016年7月、2016年8月—2016年9月)极显著高于K0处理。
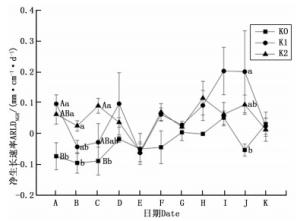
图 3 油茶细根净生长速率的动态变化
Figure 3. Dynamic changes of ARLDNRG of Camellia oleifera fine root
-
剔除掉异常数据,选择了6个重复对上(020 cm)、下(2040 cm)土层的油茶细根形态指标及根长密度进行重复测量方差分析,得到不同钾水平下油茶细根的土层分布规律(图 4)。由图 4可知,K0处理下层土中TSA、和ARLD极显著大于上层土;这说明K0处理下的油茶细根主要分布在2040 cm土层;K1处理上层土中TSA、TRT和ARLD极显著大于K0处理, 说明低钾(K1)处理极显著提高了020 cm土层细根的TSA、TRT和ARLD,而高钾(K2)处理对上层土的细根无显著促进作用,K2处理提高TSA、TRT和ARLD的作用主要体现在下层土中。就ARD而言,K0处理的上下土层ARD无显著差异,施钾之后对下层土的细根作用较明显,尤其体现在K2处理的下层土中。相比于K0处理,K2处理极显著增大了下层土的ARD。

图 4 油茶细根在不同土层中的分布规律
Figure 4. The distribution regularity of Camellia oleifera fine root in different soil layers
-
Sánchez-Calderón等[20]研究表明,钾素通过控制IAA(吲哚乙酸)氧化酶的活性,可提高IAA的含量,促进光合产物向根系转运,所以钾的富集可促进植物根系的生长。张志勇[13]、宋美珍[21]发现,钾素对棉花根系发育有促进效应,尤其是对侧根数和侧根总长的促进效应更加明显,并且钾还可以增加棉花的主根长和根系体积。潘艳花[22]等认为施钾影响西瓜根系的形态变化,不施钾条件下会明显抑制根系的生长。施用一定量的钾肥对根系的生长发育有促进作用,但过量钾肥对根系生长有一定抑制作用。赵永平等在不同供钾水平对金盏花根性形态的影响研究中亦认为适量施钾对植物根系的生长有促进作用,而缺钾在一定程度上会抑制植物根系的生长发育。本研究亦发现,施钾肥有利于油茶细根总根长、总表面积以及平均直径的增加,与对照(K0)处理相比,差异显著,这与前人研究结果一致。
本试验结果表明,在其它养分条件一致的前提下,施钾肥有利于油茶各生育时期根尖数、直径、长度以及表面积的增加。增施钾肥不仅改善了油茶细根的根系形态指标,增加了其吸收面积,并且增大了油茶细根的根长密度及净生长速率。K0处理下的油茶细根净生长速率在生育期内多为负值,K1、K2处理下的净生长速率显著高于K0处理,这说明K0处理下的油茶细根死亡速率高于生产速率,施钾肥有助于降低油茶细根的死亡速率,或者提高油茶细根的生产速率,尤其体现在K1处理下油茶细根在春季的生产量远大于其死亡量,其净生长速率在春季达到峰值。
已有研究表明,细根生长的季节动态表现为3种形态,无明显季节格局[23],单峰曲线[24]和双峰曲线[23]。本研究发现,油茶细根的季节生长节律为春季较高的单峰型。细根的死亡是一个复杂的生理生态过程[25]。从生理学角度分析,细根的死亡与光合产物的分配有关[26]。秋季为油茶果实膨大期,油茶的油脂转化需要消耗大量养分,来自地上部分的碳供应降低,根系通过死亡来抵御降低的碳供应[27-29],从而导致在油茶油脂转化时期根系较高的死亡量,油茶各形态指标在此期间降低。本研究所观察到的K0处理的油茶细根根尖数、长度、表面积以及根长密度随季节变化现象较好的印证了这一说法。而施了钾肥的K1、K2处理明显减缓了秋季油茶细根形态指标降低的趋势,说明施钾肥能够提高细根活力,促进光合产物向根系分配。根系是地下生物的重要组成部分,根系形态具有很强的可塑性,易受土壤水分、养分的影响,同时根系形态指标的变化也改变根系对养分和水分的吸收,进而影响植物物地上部分的生长[30-31]。它在植物生长过程中起到关键性作用,是植物与土壤环境接触的重要界面,它能够直接通过自身的调节,增强植物的生存能力[32-36],因此适量施钾有助于增强油茶的生存能力,促进养分高效循环利用,从而提高油茶的品质与产量。
许多对林木细根的研究结果认为林木细根生物量会随着土层深度的增加而减少[37-39],本研究却发现油茶的细根主要是分布在下层土,这与油茶是轴状深根系树种紧密相关[40]。土壤养分的差异是影响细根分布的重要原因[41-42],本研究发现无钾处理(K0)下的油茶细根主要分布在2040 cm土层,低钾(K1)处理极显著提高了020 cm土层细根的根尖数、根表面积及根长密度,而高钾(K2)处理虽然总体上提高了细根的形态指标,但对上层土的细根无显著促进作用。细根生长具有一定的趋水性和趋肥性[43-45],这可能是由于施钾改变了不同土层的土壤养分环境进而导致油茶细根在空间上的分布特征。本研究仅对油茶细根的生长动态以及空间分布进行了分析,今后会从生理生态的角度对影响油茶细根生长发育以及空间分布的机制进行进一步研究。
-
施钾处理的油茶细根根尖数、平均直径、总根长以及总表面积都有所增加,并且施钾有利于油茶根长密度及净生长速率的增大。无钾处理(K0)下的油茶细根主要分布在2040 cm土层,低钾(K1)处理极显著提高了020 cm土层细根的根尖数、根表面积及根长密度,而高钾(K2)处理虽然总体上提高了细根的形态指标,但对上层土的细根无显著促进作用。基于对油茶细根形态指标的分析,综合考虑经济因素,K1处理的钾肥施用量对油茶细根的生长与其根系形态建成效果最明显。合理适量施钾肥有助于油茶根系的生长发育,提高根系活力,增大细根的吸收面积。本研究通过观测油茶细根生长的年动态发现油茶细根的季节生长节律为春季较高的单峰型,油茶细根的直径范围为0.50.8 mm,在2017年3月出现直径峰值。而总根长、总根尖数、总表面积以及根长密度都在2017年5月出现峰值,说明油茶细根的伸长、增加以及变粗并不是同时进行,但总体上春季为油茶细根的生长旺盛期。K0处理的总根长、总根尖数、总表面积以及根长密度在2016年秋季出现下降趋势,而K1、K2处理明显减缓了其秋季细根形态指标降低的趋势,说明施钾肥能够促进光合产物向根系分配,有助于实现养分的高效循环利用。
油茶细根时空分布动态对施钾水平的响应
Response of Fine Root Growth of Camellia oleifera to Potassium Application Level
-
摘要:
目的 为了解不同施钾水平对油茶(Camellia oleifera Abel)细根时空分布动态的影响。 方法 采用微根管(minirhizotron)动态监测技术,以2011年种植的油茶林为试材,在等养分条件下,2015年设置不施钾K0(N92P48K0)、低钾K1(N92P48K135)、高钾K2(N92P48K270)3个处理,2016年6月起对林地0~40 cm土壤剖面的油茶细根的时空分布动态进行了观测。 结果 表明:K1、K2处理增大了油茶细根根尖数、平均直径、总根长以及总表面积,并且施钾有利于油茶根长密度及净生长速率的增大。不施钾处理(K0)下的油茶细根主要分布在20~40 cm土层,低钾(K1)处理极显著提高了0~20 cm土层细根的根尖数、根表面积及根长密度,而高钾(K2)处理对上层土的细根无显著促进作用。K0处理的总根长、总根尖数、总表面积以及根长密度在2016年秋季出现下降趋势,而K1、K2处理明显减缓了其秋季细根形态指标降低的趋势;油茶的平均直径在整个生育期内变化幅度较大,各处理都在2017年3月出现峰值,K0、K1、K2三个处理下的平均直径峰值分别为0.631、0.750、0.788 mm;总根长、总根尖数、总表面积以及根长密度都在2017年5月出现峰值;K1处理下的净生长速率在2017年3月-2017年5月达到峰值,并显著高于对照; 结论 油茶细根的季节生长节律为春季较高的单峰型,细根直径范围为0.5~0.8 mm;合理适量施钾肥有助于油茶根系的生长发育,提高根系活力,增大细根的吸收面积;施钾肥能够促进光合产物向根系分配,有助于实现养分的高效循环利用。 Abstract:Objective To study the effect of different potassium levels on spatial and temporal distribution dynamics of Camellia oleifera Abel fine root. Method The dynamic monitoring technique of minirhizotron was used in this study. The C. oleifera plantation planted in 2011 was used as experimental material under the condition of equal nutrient, K0(N92P48K0), K1(N92P48K135) and K2(N92P48K270) were applied to investigate the growth dynamics of fine roots of C. oleifera in 0~40 cm soil profile in the forestland. Result The results showed that the treatments K1 and K2 increased the root tip number, average diameter, total root length, and total surface area of C. oleifera, and the application of potassium was beneficial to the increase of root length and net growth rate of C. oleifera. The fine roots of C. oleifera under control treatment were mainly distributed in the 20~40 cm soil layer. The treatment with low K (K1) significantly increased the number of root tips, root surface area, and root length of fine roots in 0~20 cm soil layers. Potassium (K2) treatment had no significant effect on the fine roots of the upper layer soil. The total root length, total root tip number, total surface area and root length density of K0 decreased in autumn 2016, while the treatments K1 and K2 slowed down the decline of fine root morphology in autumn. The ARD of C. oleifera varied greatly during the whole growth period, and the peak value of ARD appeared in March 2017. The ARD peak values were 0.631mm, 0.750 mm and 0.788 mm under K0, K1 and K2 treatments respectively. The total root length, total root tips, total surface area and root length density all peaked in May 2017. The net growth rate under K1 peaked from March 2017 to May 2017 and was significantly higher than that of the control. Conclusion The seasonal growth rhythm of fine root of C. oleifera follows a single peak with a high spring in the range of 0.5~0.8 mm. Appropriate amount of K fertilizer is helpful to the growth and development of root system of C. oleifera, increase the root activity and the absorption area of fine root. Potassium fertilizer can promote the distribution of photosynthetic products to the root system, which helps to achieve efficient recycling of nutrients. -
图 1 油茶细根形态指标的动态变化
Figure 1. Dynamic changes of morphology indicators of Camellia oleifera fine root
图 3 油茶细根净生长速率的动态变化
Figure 3. Dynamic changes of ARLDNRG of Camellia oleifera fine root
图 4 油茶细根在不同土层中的分布规律
Figure 4. The distribution regularity of Camellia oleifera fine root in different soil layers
表 1 油茶细根形态指标重复测量方差分析
Table 1. Repeated measurement variance analysis results of morphology indicators of Camellia oleifera fine root
变异来源
Source of variation平均细根直径ARD 总根尖数TRT 总根长TRL 总表面积TSA P F P F P F P F 时间Time 0.002 2.834 < 0.001 3.279 0.011 2.293 0.002 2.775 时间*钾素Time*Potassium 0.017 1.803 0.001 2.305 0.073 1.501 0.351 1.095 钾素Potassium 0.029 4.132 0.194 1.758 0.042 3.358 0.012 5.290  下载: 导出CSV
下载: 导出CSV
表 2 油茶细根形态指标多重比较
Table 2. Multiple comparisons result of morphology indicators of Camellia oleifera fine root
钾素水平Potassium level 平均细根直径ARD/mm 总根尖数TRT/个 总根长TRL/ mm 总表面积TSA/ mm2 K0 0.576 6Ab 14.453 7Aa 118.874 5Ab 214.189 1Bb K1 0.688 6Aa 29.157 4Aa 261.478 7Aa 573.618 6Aa K2 0.668 7Aa 21.648 1Aa 183.487 1Aab 419.432 ABab
下载: 导出CSV
表 3 平均根长密度及细根净生长速率的重复测量方差分析
Table 3. Repeated measurement variance analysis results of ARLD and ARLDNGR
变异来源
Source of variation平均根长密度
ARLD净生长速率
ARLDNGRP F P F 时间Time 0.011 2.293 0.001 3.255 时间*钾素Time*Potassium 0.073 1.501 0.202 1.278 钾素Potassium 0.042 3.358 < 0.001 31.758
下载: 导出CSV
表 4 平均根长密度及细根净生长速率的多重比较
Table 4. Multiple comparisons result of ARLD and ARLDNGR
钾素水平
Potassium level平均根长密度
ARLD/ (mm·cm-3)净生长速率
ARLDNGR /(mm·cm-3·d-1)K0 5.896 6Ab -0.031 1Bb K1 12.970 2Aa 0.060 6Aa K2 9.101 5Aab 0.048 1Aa
下载: 导出CSV
-
[1] 胡冬南, 牛德奎, 张文元, 等.钾肥水平对油茶果实性状及产量的影响[J].林业科学研究, 2015, 28(2): 243-248. [2] 张文元, 牛德奎, 郭晓敏, 等.施钾水平对油茶养分积累和产油量的影响[J].植物营养与肥料学报, 2016, 22(3): 863-868. [3] 俞元春, 白玉杰, 俞小鹏, 等.油茶林施肥效应研究概述[J].林业科技开发, 2013, 27(2): 1-4. doi: 10.3969/j.issn.1000-8101.2013.02.001 [4] 牛学礼, 南志标.运用微根管技术研究草地植物细根的进展[J].草业学报, 2017, 26(11): 205-215. doi: 10.11686/cyxb2017041 [5] Yuan Y D, Yang Y S, Chen G S, et al. Fine root longevity of a Cunninghamia lanceolata plantation estimated by minirhizotrons[J]. Journal of Subtropical Resources and Environment, 2009, 4(2): 47-52. [6] 陈展, 于浩, 尚鹤, 等.臭氧胁迫对树木根系影响研究进展[J].林业科学研究, 2016, 29(3): 455-463. doi: 10.3969/j.issn.1001-1498.2016.03.023 [7] 张永清, 毕润成, 庞春花, 等.不同品种春小麦根系对低钾胁迫的生物学响应[J].西北植物学报, 2006, 26(6): 1190-1194. doi: 10.3321/j.issn:1000-4025.2006.06.016 [8] 张志勇, 王清连, 李召虎, 等.缺钾对棉花幼苗根系生长的影响及其生理机制[J].作物学报, 2009, 35(4): 718-723. [9] Shin R, Schachtman D P. Hydrogen peroxide mediates plant root cell response to nutrient deprivation[J]. Proc Natl Acad, 2004, 101(23): 8827-8832. doi: 10.1073/pnas.0401707101 [10] Johnson M G, Tingey D T, Philips D L, et al. Advancing fine root research with minirhizotrons[J]. Environmental and Experimental Botany, 2001, 45(3): 263-289. doi: 10.1016/S0098-8472(01)00077-6 [11] Shibata H, Hiura T, Tanaka Y, et al. Carbon cycling and budget in a forested basin of southwestern Hokkaido, northern Japan[J]. Ecological Research, 2005, 20(3): 325-331. doi: 10.1007/s11284-005-0048-7 [12] Mei L, Wang Z Q, Cheng Y H, et al. A review: factors influencing fine root longevity in forest ecosystems[J]. Acta Phytoecologica Sinica, 2004, 28(4): 704-710. [13] Chen X Y, Eamus D, Hutley L B. Seasonal patterns of fine-root productivity and turnover in a tropical savanna of northern Australia[J]. Journal of Tropic Ecology, 2004, 20(2): 221-224. doi: 10.1017/S0266467403001135 [14] 张志山, 李新荣, 张景光, 等.用Minirhizotrons观测柠条根系生长动态[J].植物生态学报, 2006, 30(3): 457-464. doi: 10.3321/j.issn:1005-264X.2006.03.012 [15] Gordon W S, Jackson R B. Nutrient concentrations in fine roots[J]. Ecology, 2000, 81(1): 275-280. doi: 10.1890/0012-9658(2000)081[0275:NCIFR]2.0.CO;2 [16] Burton A J, Pregitzer K S, Hendrick R L. Relationships between fine root dynamics and nitrogen availability in Michigan northern hardwood forests[J]. Oecologia, 2000, 125(3): 389-399. doi: 10.1007/s004420000455 [17] Zhang X Q, Wu K H, Murach D. A review of methods for fine-root production and turnover of trees[J]. Acta Ecologica Sinica, 2000, 20(5): 815-883. [18] 张亚雄.蓄水坑灌下苹果树细根动态及其影响因素的研究[D].太原: 太原理工大学, 2017. [19] 周长富, 姚小华, 林萍, 等.油茶果实发育特性及水分、油脂含量动态分析[J].扬州大学学报, 2013, 34(3): 49-53+82. [20] 王立梅, 刘奕清, 阮玉娟, 等.植物钾素研究进展[J].中国园艺文摘, 2015(5): 71-72. doi: 10.3969/j.issn.1672-0873.2015.05.029 [21] Sánchez-Calderón L, López-Bucio J, Chacón-López A, et al. Phosphate starvation induces a determinate developmental program in the roots of Arabidopsis thaliana[J]. Plant Cell Physiol, 2005, 46(1): 174-184. [22] 宋美珍, 杨惠元, 蒋国柱.黄淮海棉区钾肥效应研究[J].棉花学报, 1993, 5(1): 73-78. [23] 潘艳花, 马忠明, 吕晓东, 等.不同供钾水平对西瓜幼苗生长和根系形态的影响[J].中国生态农业学报, 2012, 20(5): 536-541. [24] Persson H.Root dynamics in a young Scots pine stand in Central Sweden[J]. Oikos.1978, 30(3): 508-519. doi: 10.2307/3543346 [25] Mc Claugherty C A, Aber J D, Melillo J M, The role of fine roots in the organic matter and nitrogen budgets of two forested ecosystems[J]. Ecology, 1982, 63(5): 1481-1490. doi: 10.2307/1938874 [26] 陈建文, 王孟本, 史建伟.柠条人工林幼林与成林细根动态比较研究[J].生态学报, 2011, 31(22): 6978-6988. [27] Farrar J F, Jones D L. The control of carbon acquisition by roots[J]. New Phytologist, 2000, 147(1): 43-53. doi: 10.1046/j.1469-8137.2000.00688.x [28] Mou P, Robert H J, Tan Z Q, et al. Morphological and physiological plasticity of plant roots when nutrients are both spatially and temporally heterogeneous[J]. Plant and Soil, 2013, 364: 373-384. doi: 10.1007/s11104-012-1336-y [29] 梅莉, 王政权, 程云环, 等.林木细根寿命及其影响因子研究进展[J].植物生态学报, 2004, 28(4): 704-710. [30] Zou X H, Wu P F, Chen N L, et al. Chinese fir root response to spatial and temporal heterogeneity of phosphorus availability in the soil[J]. Canadian Journal of Forest Research, 2014, 45: 402-410. [31] 汤丹, 龚榜初, 江锡兵, 等.不同甜柿砧穗组合根系差异性研究[J].林业科学研究, 2016, 29(1): 85-92. doi: 10.3969/j.issn.1001-1498.2016.01.013 [32] 沈仁芳, 孙波, 施卫明, 等.地上-地下生物协同调控与养分高效利用[J].中国科学院院刊, 2017, 32(6): 566-574. [33] 赵其国, 沈仁芳, 滕应.中国土壤安全"一带一路"发展战略的思考[J].生态环境学报, 2016, 25(3): 365-371. [34] 杨林章, 孙波.中国农田生态系统养分循环和平衡及其管理.[M].北京:科学出版社, 2008. [35] 于飞, 施卫明.近10年中国大陆主要粮食作物氮肥利用率分析[J].土壤学报, 2015, 52(6): 1311-1324. [36] 张福锁, 王激清, 张卫峰, 等.中国主要粮食作物肥料利用率现状与提高途径[J].土壤学报, 2008, 45(5): 915-924. doi: 10.3321/j.issn:0564-3929.2008.05.018 [37] Wardle D A, Bardgett R D, Klironomos J N, et al. Ecological linkages between aboveground and below ground biota[J]. Science, 2004, 304(5677): 1629-1633. doi: 10.1126/science.1094875 [38] 秘洪雷, 兰再平, 孙尚伟, 等, 滴灌栽培杨树人工林细根空间分布特征[J].林业科学研究, 2017, 30(6): 946-953. [39] Wang H Y, Dai W, Yang X J, et al. Spatial heterogeneity of soil organic carbon and nutrients in low mountain area of Changbai Mountains[J]. Chinese Journal of Applied Ecology, 2014, 25(9): 2460-2468. [40] Mou P, Jones R H, Mitchell R J, et al. Spatial distribution of roots in Sweetgum and Loblolly pine monocultures and relations with a-bove-ground biomass and soil nutrients[J]. Functional Ecology, 1995, 9(4): 689-698. doi: 10.2307/2390162 [41] 周政贤.油茶生态习性、根系发育及垦覆效果调查研究[J].林业科学, 1963, 8(4): 336-346. [42] 王娜, 程瑞梅, 肖文发, 等.三峡库区马尾松细根分解及其养分释放[J].林业科学研究, 2017, 30(1): 18-24. [43] 闫小莉, 戴腾飞, 邢长山, 等.水肥耦合对欧美108杨幼林表土层细根形态及分布的影响[J].生态学报, 2015, 35(11): 3692-3701. [44] 张小全.环境因子对树木细根生物量、生产与周转的影响[J].林业科学研究, 2001, 14(5): 566-573. doi: 10.3321/j.issn:1001-1498.2001.05.017 [45] 程云环, 韩有志, 王庆成, 等.落叶松人工林细根动态与土壤资源有效性的研究[J].植物生态学报, 2005, 29(3): 403-410. doi: 10.3321/j.issn:1005-264X.2005.03.009 [46] 李帅锋, 贾呈鑫卓, 杨利华, 等.林龄对思茅松人工林根系生物量的影响[J].林业科学研究, 2018, 31(2): 26-33. -

 点击查看大图
点击查看大图

计量
- 文章访问数: 3836
- HTML全文浏览量: 1977
- PDF下载量: 165
- 被引次数: 0
