



-
被子植物的叶通常呈表面积巨大的扁平状,能够同外界环境进行广泛的物质、能量与信息交流,是个体碳同化和气体交换的主要场所[1]。叶的形态多样性极为丰富,被普遍应用于分类学领域的同时,也受到遗传育种工作者的广泛关注。研究表明,叶的形态特征同植物的生存与生长紧密相关[2-4],可作为农作物或林木良种选育的早期指标,因而,解析叶形性状的遗传机理可以为育种实践提供理论支持。目前,应用分子生物学手段对叶形态建成的遗传控制机制展开了大量研究,发现多种植物激素、转录因子和小RNA均参与其中[5-7];在家系群体中,利用遗传图谱对水稻(Oryza sativa L.)[8]、玉米(Zea mays L.)[9]、小麦(Triticum aestivum L.)[10]等农作物控制叶形性状的数量性状位点(Quantitative Trait LQTLs)进行定位并已据此获得多个关键基因;对杜仲(Eucommia ulmoides O.)[11]、茶树(Camellia sinensis L.)[12]及杨树(Populus L.)[4, 13]等木本植物也开展了相关研究,然而,主效QTLs及相关候选基因的定位仍较困难。随着测序技术的进步,图谱密度不再是制约林木性状基因组定位的瓶颈,建立相应规模的作图群体并结合高通量测序手段,便可对控制目标性状的遗传位点进行精细定位[14-16],从而为进一步深入解析叶形性状的遗传机理提供良好契机。叶的形态属于多元复合型性状[15, 17],在大群体尺度对其进行全面、准确的表型测定是该领域研究的关键所在[18]。目前,叶形性状的测定主要有传统手工测量、叶面积仪扫描和计算机图像分析3种方法[19]。传统手工测量目的性强,但效率和精度难以保证;计算机图像处理仍处于探索阶段,相关软件开发有待进一步完善[20],因而,现阶段相关研究主要采用叶面积仪扫描法。便携式叶面积仪操作简易,适宜对高通量样本进行处理,但获得的信息量有限,多数仪器仅可读取叶片的面积、周长、长和宽4项指标[19],而漏掉如叶脉夹角、最大叶宽位置等重要的叶形态学信息。
杨树是我国主要造林树种之一,在阔叶树遗传改良研究中占据重要地位,解析其叶形性状的遗传变异规律意义重大。杨属五大派叶形性状差异较大,通过派间杂交获得的子代叶的形态多样性非常丰富[16, 21]。丹红杨(P. deltoides CL. ‘Danhong’)是由人工选育的具有速生、抗蛀干害虫等优良特性的南方型美洲黑杨新品种,其叶片巨大、呈三角形,叶柄长,叶基呈截形[22]。小叶杨(P. simonii)为我国北方地区青杨派主要乡土树种之一,具备广泛的适应性和优良的抗逆性,其叶片小,呈菱状卵形、椭圆形或倒卵形,叶柄极短,叶基楔形、宽楔形或窄圆形,叶缘平整,具细锯齿[23]。以丹红杨为母本,以从内蒙古通辽地区小叶杨天然林选出的优树通辽1号杨为父本,通过室内切枝水培人工控制授粉获得的F1代,观察发现,子代叶片大小与形状的变异极为丰富[24]。本研究以丹红杨×通辽1号杨亲本及F1代为试验材料,采用叶面积仪扫描辅以传统手工测量的方式对叶面积、叶柄长等7个叶表型性状进行测定,经计算获得4个复合指标,较为全面地提取了叶的形状与大小信息,利用多种统计方法对所测指标进行综合分析,揭示了杨树叶形性状的遗传变异规律,为进一步探索其分子遗传机制奠定基础。
-
2016年2月,以丹红杨为母本,通辽1号杨为父本,采用室内切枝水培的方法,经人工控制授粉杂交获得F1代材料,并分别在温室和北京玉泉山苗圃中完成育苗工作。2017年3月,将材料分3个随机区组扦插于中国林业科学研究院温室内,并进行统一管理。
-
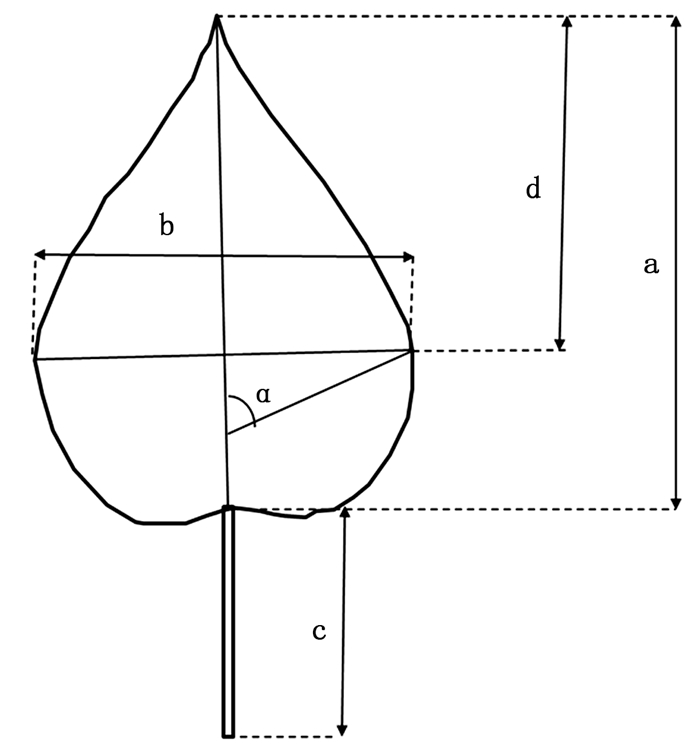
2017年8月上旬,对全部422个杂交子代及其亲本材料的第9、10片叶(从茎尖向下完全展开的叶片记为第1片叶)进行取样,每系号3株,共计6片叶。使用Yaxin-1241便携式叶面积仪,扫描并记录叶片的面积、长、宽及周长4个指标,并借助刻度尺、量角器等工具手工测量叶柄长、最大叶宽位置以及主叶脉与最大叶宽处侧脉的夹角,总计获取7个度量指标。叶片长度与夹角测定的详细情况见图 1。
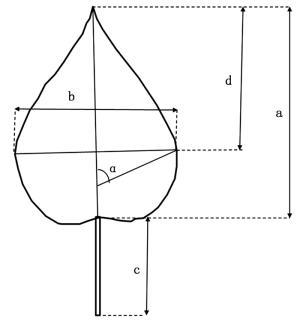
图 1 叶片度量示意图
Figure 1. Schematic of leaf blade measurement
-
使用Excel 2013软件录入数据并利用公式(1)、(2)、(3)和(4)计算获得叶长宽比、最大叶宽位置、叶柄相对长和叶缘因子4个指标。使用SPSS 19.0软件进行均值比较、描述统计、正态检验、相关分析以及主成分因子分析。
$叶长宽比 = 叶长/叶宽 = b/a$
(1) $最大叶宽位置 = 叶最宽处距叶尖距离/叶长 = d/a$
(2) $叶柄相对长 = 叶柄长/叶长 = c/a$
(3) $叶缘因子 = 4 \times 叶面积/{叶周长^2}$
(4) 以叶长、叶宽、侧脉夹角和最大叶宽距叶尖长度为自变量X,叶面积为因变量Y进行通径分析[25]并计算决定系数。式(5)中:PY·Xi是Xi对Y的直接通径系数,SXi是自变量Xi的标准差;SY是因变量Y的标准差;式(6)中:PXi·Yj是Xi通过Xj对Y的间接通径系数,rij是相关系数,Pj·Y是Xj对Y的通径系数;式(7)中:di是直接决定系数;式(8)中:dij是间接决定系数。
${P_{Y \cdot Xi}} = {b_i} \times {S_{Xi}}/{S_Y}$
(5) ${P_{Xi \cdot Yj}} = {r_{ij}} \times {P_{j \cdot Y}}\left( {i \ne j} \right)$
(6) ${d_i} = {P_{Y \cdot Xi}}^2$
(7) ${d_{ij}} = 2 \times {r_{ij}} \times {P_{Y \cdot Xi}} \times {P_{Y \cdot Xj}}$
(8) -
本研究的杂交母本丹红杨与父本通辽1号杨分别属于杨树中黑杨派(Aigeiros)和青杨派(Tacamahaca),其叶片形状存在较大的种间差异。图 2A、2B表明:丹红杨的叶片呈巨大的三角形,叶柄长,叶基截形;通辽1号杨的叶片呈较小的倒卵形,叶柄极短,叶基楔形。表 1显示:两亲本除了叶缘因子外其余9个叶形指标差异均达极显著水平(P<0.01)。丹红杨的叶面积5 608.4 mm2,是通辽1号杨的5.01倍;叶柄长62.1 mm,为通辽1号杨的11.29倍;另外叶片的周长、长、宽和侧脉夹角等指标均明显大于通辽1号杨。对于表征叶片形状的指标而言,丹红杨的叶长与叶宽较接近,最大宽度位于靠近叶片基部位置,而通辽1号杨的叶长是其宽度的1.405倍,最大叶宽所处位置距叶尖仅18.7 mm,占叶片总长度的38.8%。

图 2 亲本和子代叶形变异情况
Figure 2. Leaf shape variation among parents and F1 hybrid progenies
表 1 丹红杨和通辽1号杨叶形性状分析
Table 1. The analysis of leaf morphological traits of P. deltoides CL. 'Danhong' and P. simonii CL. 'Tongliao1'
性状
Traits丹红杨
P. deltoides CL.‘Danhong’通辽1号杨
P. simonii CL.‘Tongliao1’T检验(t值)
T-test (t value)显著性(P值)
Significance (P value)叶面积Leaf area (LA)/mm2 5 608.4±492.7 1 118.9±100.1 26.79 1.24×10-9 叶长Leaf length (LL)/mm 94.2±5.7 48.2±3.8 20.21 1.09×10-11 叶宽Leaf width (LW)/mm 86.2±2.7 34.3±0.6 57.24 1.20×10-12 叶柄长Petiole length (PL) /mm 62.1±6.2 5.5±1.8 26.22 4.98×10-10 叶周长Leaf perimeter (LP)/mm 282.1±16.7 125.7±7.0 25.87 5.11×10-11 侧脉夹角Lateral vein angle (LVA)/° 66.1±5.3 50.6±3.0 7.61 4.70×10-6 叶长宽比Ratio of length and width (LL/LW) 1.092±0.043 1.407±0.088 -9.59 7.26×10-7 最大叶宽位置Position of leaf width (LWP) 0.794±0.081 0.388±0.056 12.38 4.81×10-9 叶柄相对长Relative length of petiole (RPL) 0.661±0.077 0.113±0.032 19.76 8.29×10-10 叶缘因子Leaf margin factor (LMF) 0.282±0.019 0.283±0.008 -0.154 0.88 -
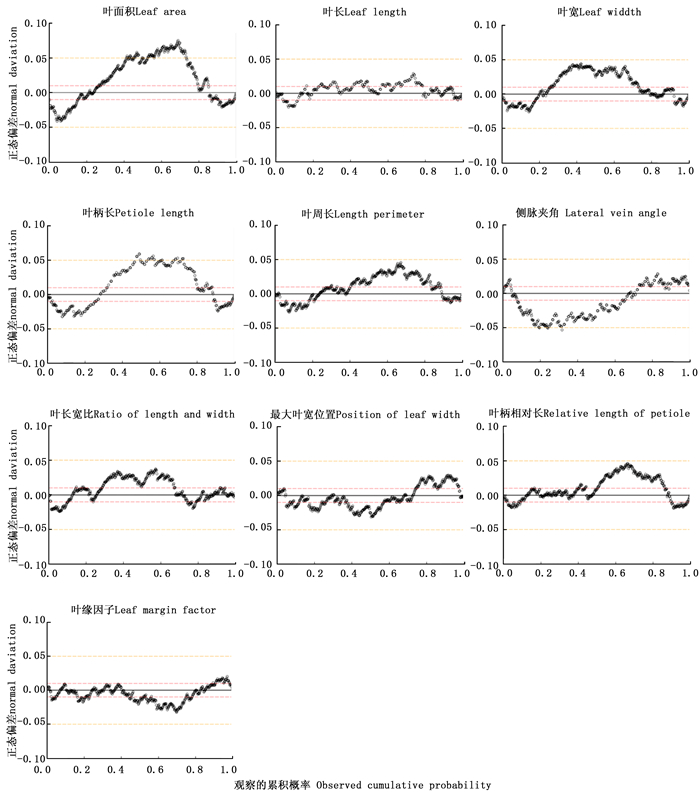
杂交子代叶形性状的变异非常丰富(图 2C~F)。表 2表明:10个指标的峰度均为正值,表明子代叶形性状的分布相对于标准正态分布更为集中。使用Blom比例估算公式对子代叶形性状的分布进行标准正态检验,结果显示:其正态偏差总体介于-0.05~0.05之间,符合正态分布(图 3)。子代叶面积的极小值与极大值分别为1 071.233 mm2和6 063.233 mm2,均值为2 780.291 mm2。叶面积、叶长、叶宽、叶柄长和叶周长是表征大小的指标,其变异系数均超过0.1,其中,叶面积和叶柄长的变异系数大于0.2,分别达0.272和0.215,说明这2个性状在子代中的变异最为丰富。侧脉夹角、叶长宽比、最大叶宽位置、叶柄相对长和叶缘因子5个指标描述了叶的形状,其中,仅叶长宽比和叶柄相对长2个指标的变异系数大于0.1。叶形性状的频率分布见图 4,全部10个指标在子代中均呈连续分布,其中,侧脉夹角和叶缘因子的偏度为负值,相对于标准正态分布稍左偏,其余8个指标的分布则相对略右偏。
表 2 F1代叶形性状分布统计量
Table 2. The statistical analysis of leaf morphological traits in F1 progenies
性状
Traits均值
Mean value极小值
Minimal value极大值
Maximal value标准差
Standard deviation变异系数
Variable coefficient偏度
Skewness峰度
Kurtosis叶面积LA/mm2 2 780.291 1 071.233 6 063.233 757.538 0.272 0.814 1.080 叶长LL/mm 71.077 46.333 103.333 8.741 0.123 0.251 0.348 叶宽LW/mm 57.882 35.000 88.617 9.189 0.159 0.458 0.286 叶柄长PL/mm 31.681 12.667 57.500 6.825 0.215 0.545 0.698 叶周长LP/mm 195.330 120.133 291.617 25.593 0.131 0.412 0.797 侧脉夹角LVA/° 59.090 32.400 72.600 5.344 0.090 -0.890 2.436 叶长宽比LL/LW 1.252 0.767 2.084 0.174 0.139 0.496 1.261 最大叶宽位置LWP 0.641 0.412 0.923 0.055 0.086 0.012 1.925 叶柄相对长RPL 0.446 0.247 0.684 0.078 0.174 0.377 0.237 叶缘因子LMF 0.287 0.168 0.346 0.021 0.075 -0.507 1.729 注:变异系数=标准差/均值。
Note: Variable Coefficient = Standard Deviation / Mean Value.
图 3 F1代叶形性状的正态检验
Figure 3. Normality test of leaf morphological traits in F1 progenies

图 4 F1代叶形性状的频率分布
Figure 4. Frequency distribution of leaf morphological traits in F1 progenies
-
为阐明10个叶形指标间的相互关系,采用Pearson参数进行相关分析,结果(表 3)表明:在10个叶形指标间45对两两相关关系中,多达31对呈极显著水平(P<0.01),另有5对为显著水平(P<0.05),其中,叶面积、叶周长、叶长、叶宽和叶柄长5个表征大小的指标相互间均呈极显著的正相关关系。另外5个表征形状的指标中,叶长宽比与叶面积的关系最密切,相关系数为-0.316。叶缘因子是描述叶缘相对生长情况的重要指标,它与叶长宽比的相关系数为-0.641,达到极显著水平。侧脉夹角、最大叶宽位置和叶柄相对长反映了叶的侧向生长情况,其中,最大叶宽位置与叶柄相对长之间存在极显著的正相关关系,相关系数为0.268,而这2个指标同侧脉夹角间则未发现显著性相关关系。
表 3 F1代叶形性状的相关分析
Table 3. Correlation analysis of leaf morphological traits in F1 hybrid progenies
性状
Traits叶面积
LA叶周长
LP叶长
LL叶宽
LW叶柄长
PL侧脉夹
角LVA叶长宽比
LL/LW叶柄相对
长RPL最大叶宽
位置LWP叶缘因子
LMF叶面积LA 1.000 0.945** 0.785** 0.889** 0.523** 0.173** -0.316** 0.103* -0.188** 0.265** 叶周长LP 1.000 0.874** 0.806** 0.587** 0.107* -0.146** 0.117* -0.096* -0.038 叶长LL 1.000 0.553** 0.585** -0.060 0.256** 0.028 0.001 -0.224** 叶宽LW 1.000 0.428** 0.241** -0.649** 0.144** -0.162** 0.378** 叶柄长PL 1.000 -0.004 0.031 0.820** 0.204** -0.122* 侧脉夹角LVA 1.000 -0.328** 0.038 0.089 0.274** 叶长宽比LL/LW 1.000 -0.138** 0.191** -0.641** 叶柄相对长RPL 1.000 0.268** 0.013 最大叶宽位置LWP 1.000 -0.325** 叶缘因子LMF 1.000 注:*表示在0.05水平(双侧)显著相关;**表示在0.01水平(双侧)显著相关。
Note: *Indicates when P<0.05, there is significant correlation; **indicates when P<0.01, there is extremely significant correlation.以叶长(LL)、叶宽(LW)、侧脉夹角(LVA)和最大叶宽距叶尖长度(LWD)为自变量,叶面积(LA)为因变量进行通径分析,结果(表 4)表明:。在4个指标中,叶长和叶宽对叶面积的通径系数绝对值最大(0.611和0.604),表明二者对叶面积的直接作用最大,其次是最大叶宽距叶尖长度(-0.189),侧脉夹角作用最小(0.064)。通过间接通径系数计算发现,其它指标通过叶长、叶宽以及最大叶宽距叶尖长度对叶面积的间接作用总和均大于直接作用,其中,最大叶宽距叶尖长度通过叶长对叶面积施加的间接影响最大(0.508)。
表 4 F1代4个叶片性状对叶面积的通径分析
Table 4. Path coefficient analysis of four leaf blade traits on leaf area in F1 progenies
性状
Traits相关系数
Correlation coefficient直接通径系数
Direct path coefficient间接通径系数Indirect path coefficient LL LW LVA LWD 总和(SUM) LL 0.785** 0.611 0.337 -0.037 0.508 0.808 LW 0.889** 0.604 0.334 0.146 0.218 0.697 LVA 0.173** 0.064 -0.004 0.015 2.555×10-4 0.011 LWD 0.537** -0.189 -0.157 -0.068 0.001 -0.224 注:*表示在0.05水平(双侧)显著相关;**表示在0.01水平(双侧)显著相关。
Note: *Indicates when P<0.05, there is significant correlation; **indicates when P<0.01, there is extremely significant correlation.各性状对叶面积的决定程度分析见表 5。叶长和叶宽是影响叶面积最重要的因素,其对叶面积的决定系数分居前两位(0.374和0.365),对R2的贡献分别高达0.479和0.537,最大叶宽距叶尖长度的影响居第三位,而侧脉夹角的影响则最小。剩余通径系数(0.272)虽然超过最大叶宽距叶尖长度(0.189)和侧脉夹角(0.064)2个指标对叶面积的直接通径系数,但却远小于叶长(0.611)和叶宽(0.604)的贡献,表明叶长和叶宽是决定叶面积的主要因素。
表 5 F1代4个叶片性状对叶面积的决定系数
Table 5. Determination coefficients of 4 leaf blade traits on leaf area in F1 progenies
性状
TraitsLL LW LVA LWD R2 剩余通径系数
Remanent path coefficient各性状对R2的贡献
Contribution to R2LL 0.374 0.408 -0.005 -0.192 0.926 0.272 0.479 LW 0.365 0.019 -0.082 0.537 LVA 0.004 9.647×10-5 0.011 LWD 0.036 -0.101 -
为阐明该杨树杂交组合叶形性状的亲、子代遗传规律,比较了其亲本与子代的均值,并计算各指标在亲子代间的遗传偏离程度,结果(表 6)表明:。叶面积、最大叶宽位置、叶柄相对长以及叶缘因子4个指标亲子代偏离度的绝对值均大于0.2,表明这些性状的遗传具有一定的偏向性,其中,最大叶宽位置与叶柄相对长的偏离度为正值,表明其在亲、子代间的遗传偏向于母本,另2个指标与之相反;而对于叶长、叶宽、叶柄长、叶周长、侧脉夹角以及叶宽长比6个指标,其亲子代偏离度的绝对值均小于0.2,可以认为这些性状在总体上遵循加性遗传规律,不存在明显的遗传偏向性。杂交子代最大叶宽位置和叶柄相对长分别有多达83.18%和77.73%大于中亲值,表明这2个叶形指标的遗传可能受母本影响较大;而对于叶面积,则仅20.62%的子代大于中亲值,推测其父本效应更明显。另外,还统计了子代中超亲无性系所占比例,从表 6可以看到:叶长宽比和侧脉夹角2个指标的超亲子代数目较多,分别占全部子代的36.73%和13.27%。
表 6 亲、子代叶形性状的遗传分析
Table 6. Genetic analysis of leaf morphological traits between F1 progenies and their parents
性状
Traits丹红杨
P.deltoides CL. ‘Danhong’通辽1号杨
P.simonii CL. ‘Tongliao1’中亲值
Mean value of parents子代均值
Mean value of progenies亲子代偏离度b
Degree of deviation between parents
and progenies大于中亲值子代比例
Percentage of progenies exceeded the mean value of parents/%超亲子代比例
Percentage of transgressive progenies/%叶面积LA/mm2 5 608.4 1 118.9 3 363.7 2 780.3 -0.260 20.62 0.47 叶长LL/mm 94.2 48.2 71.2 71.1 -0.004 49.05 1.66 叶宽LW/mm 86.2 34.3 60.3 57.9 -0.091 37.20 0.71 叶柄长PL/mm 62.1 5.5 33.8 31.7 -0.074 32.94 0.00 叶周长LP/mm 282.1 125.7 203.9 195.3 -0.110 32.70 0.71 侧脉夹角LVA/° 66.1 50.6 58.4 59.1 0.097 58.53 13.27 叶宽长比LW/LL a 0.916 0.711 0.813 0.799 -0.142 48.82 36.73 最大叶宽位置LWP 0.794 0.388 0.591 0.641 0.246 83.18 0.24 叶柄相对长RPL 0.661 0.113 0.387 0.446 0.215 77.73 0.95 叶缘因子LMF 0.282 0.283 0.283 0.287 -9.000 57.58 100.00 注:a:为便于比较,采用叶长宽比的倒数来表示其相对值;b:亲子代偏离度=(子代均值-中亲值)/(较大亲本值-中亲值)。
Note: a:To make the comparison more convenience, we use the reciprocal of length-width ratio for its relative value.
b:Divergence between F1 progenies and their parents = (Mean value of F1 progenies-Mid-parents value) /(Larger value of parents-Mid-parents value).由于所测指标间存在广泛且显著的相关关系(表 3),为直观描述杂交子代各叶形性状的总体分布及遗传变异情况,对叶面积、周长、长、宽、侧脉夹角、最大叶宽到叶尖距离以及叶柄长7个直接测量的指标进行主成分分析(PCA)。表 7表明:共提取到2个特征值大于1的公因子,累计能够解释80.19%的整体叶形变异信息;其中,第一主成分贡献率达63.71%,主要载荷叶面积、长、宽、周长、最大叶宽到叶尖距离以及叶柄长;第二主成分贡献率为16.48%,主要载荷侧脉夹角,并携带有较多叶面积和叶宽的信息。
表 7 F1代叶形性状的主成分特征向量
Table 7. Principal component eigenvectors of leaf morphological traits in F1 progenies
性状
Traits第一主成分
The first principal component第二主成分
The second principal component特征值Eigenvalue 4.459 1.154 累计贡献率/%
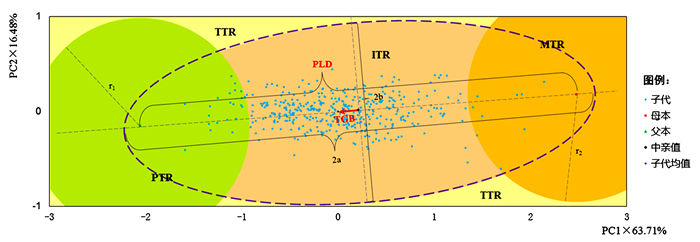
Accumulated contribution rate63.71 80.19 叶面积LA 0.859 0.413 叶周长LP 0.926 0.279 叶长LL 0.942 -0.029 叶宽LW 0.700 0.554 叶柄长PL 0.761 -0.043 侧脉夹角LVA -0.070 0.880 最大叶宽距叶尖长度LWD 0.842 -0.123 根据2个主成分(PC)的贡献率进行统一转化,并利用转化后的变量作2D点图(图 5),以展示叶的整体形态特征在该杂交群体中的遗传结构,各无性系在图中的物理距离表征其叶形性状的总差异度。第一主成分全部落在两亲本之间,第二主成分有127个(30.09%)子代分布于父母本之外,表现出明显的超亲分离现象(图 5)。子代均值位于两亲本连线上,表明子代叶形总体上受两亲本较强的遗传控制。以所有子代各自与中亲值叶形差异的标准差(SD)加上两亲本叶形总差异(PLD)为接受阈(2a),则与父母本所在位置图距之和小于2a(紫色椭圆虚线内)的子代均可以认为受到了亲本强有力的遗传控制,图 5中的子代全部落于这一范围内,且其均值相对于中亲值的遗传偏距(TGB)远小于两亲本间遗传宽度(PLD),推测叶形性状在该群体中主要遵循加性遗传模式。将与通辽1号杨相距小于1/4PLD的区域划定为父本类叶形分布区(PTR),同理可得到母本类型分布区(MTR),以亲本加性遗传控制区的边界将剩余平面划分为中间类叶形分布区(ITR)与超亲类叶形分布区(TTR)。结果显示:绝大部分子代表现为中间类型(92.66%),父本及母本类型子代数量较少,分别仅占5.21%和2.13%,这再次表明该杂交组合中亲本对子代叶形的整体遗传控制模式应当是以加性效应为主,不具备明显的遗传偏向性。

图 5 杂交群体叶形主成分2D点云图
Figure 5. Two-dimensional (2D) distributional scatter with principal component of leaf morphological traits in hybrid population
-
叶的形态学特征属于多元复合性状,包含叶片形状与大小、叶脉分布、叶柄以及叶表面附属物等信息[15, 17]。天然种群叶形变异由自然选择主导,而在人工群体中则与其杂交亲本密切相关[27]。Drost等[16]对美洲黑杨与毛果杨回交群体(BC1)的研究发现,叶宽及叶长宽比代表了BC1中叶形变异的绝大部分信息,而这也是其亲本叶形的主要差异;张福敏[27]对响叶杨与银白杨杂交群体的研究同样表明,子代叶形变异幅度取决于其亲本间差异。本研究的亲本丹红杨和通辽1号杨叶面积、叶柄长和最大叶宽位置等指标差异极显著,在F1代中这些指标的变异幅度同样较大;而对于像叶缘因子这样亲本间差别不明显的指标,在F1代中的变异系数则较小;值得注意的是,丹红杨与通辽1号杨叶长宽比的差异不如最大叶宽位置明显,但在F1中其变异幅度却远大于后者,其中,“超亲类型子代”所占比例高达36.73%,推测在该杂交组合中控制叶长宽比的等位及非等位基因间可能存在广泛的互作。通过对叶形指标的主成分分析发现,子代叶的整体形态特征可能受两亲本较强的加性遗传控制,未发现明显的遗传偏向,但通过对具体特征指标的遗传分析则观察到较明显的偏向性,这是由于在叶形整体特征的提取中可能漏掉部分重要的遗传偏向信息,因而,在对叶形性状进行整体遗传解析时应当充分考虑其特征变量发挥的作用。
叶片在平面内不同方向不对称性生长(即叶的异速生长)是其形态多样性产生的主因[15],由于杨树的叶为典型的轴对称结构,拥有明显的主叶脉[23],因而,叶长宽比成为表征其异速生长最关键的指标。解剖学相关理论表明,叶长和叶宽分别由叶细胞在顶端和侧向的增殖决定,并受相对独立的遗传控制[28-29]。本研究叶长与叶宽在子代中的变异系数都大于0.1,且与长宽比间均呈极显著相关,但叶宽与长宽比的相关系数远大于叶长,表明叶长宽比在该杂交群体中的表型变异由叶片在顶端和侧向的生长共同决定,其中侧向生长(叶宽)的变异发挥主导作用。多项研究显示,当植物在遭受诸如干旱、饥饿等胁迫下,往往是叶宽而非叶长会成为决定其长宽比的主因,叶片选择牺牲部分侧向生长,令叶肉组织尽可能收缩于主叶脉两侧,从而有效降低物质运输中能量损耗和水分散失[29-30]。基于此,在本研究的杂交群体中利用变异幅度较大的叶宽或叶长宽比作为抗旱性状的辅助性选择指标,有望提升育种工作效率。
叶面积是表征植物光合与蒸腾能力的重要参数,可作为林木生长性状的早期选择指标[2, 31]。本研究的杂交子代叶面积变异系数高达0.272,具备较强的选择潜力,利用通径分析手段,发现杨树的叶长和叶宽是决定其叶面积的主要因素。有研究表明,叶宽而非叶长是决定叶面积的主要因素[15-16],但这些结果往往是通过叶长性状遗传基础较窄的研究对象得出的;本研究两亲本叶片长度与宽度的差异均较大,在整个群体中变异丰富,因而得出二者共同决定叶面积大小的结果同样较为合理。此外,本研究422个子代的叶长和长宽比大体平均分布于中亲值两侧,而叶宽和叶面积则有超过60%的子代与父本分布同侧,可以认为该杂交子代叶片宽度的遗传偏向性是影响叶面积分布的主要因素,这表明相对于叶片长度,其宽度方向的生长在叶面积的遗传控制中更能起到决定性的作用。
最大叶宽位置对叶片形态影响巨大,是物种分类的重要依据,但由于其无法通过叶面积仪直接读出,因而目前在群体水平上对该性状的研究仍较有限。研究表明,青杨和藏川杨的最大叶宽位置在不同地理分布区存在显著差异[32-33]。本研究杂交父本通辽1号杨最大叶宽处于叶片中部以上,在整个杨属中较罕见[23];母本丹红杨作为典型的美洲黑杨,其叶片最大宽接近叶基位置[22];由于两亲本该性状表现为杨属内两个极端类型,因而其F1代家系群体是研究该性状的绝佳材料。本研究中有超过83%的子代最大叶宽位置大于中亲值,具有明显的母本效应,但其遗传偏向的机理目前仍不清楚。研究表明,不同叶形指标往往存在相同的遗传控制区域,表型性状的相关性往往能反映其遗传位点的连锁程度[34],通过对各叶形指标与最大叶宽位置进行相关性分析,有助于该性状的遗传解析。在本研究表征叶片形状的5个指标中,同最大叶宽位置相关性最强的是叶缘因子,其次为叶柄相对长和叶柄长,与侧脉夹角间则无显著性相关关系,这表明利用叶缘相对生长、叶柄分化以及叶片异速生长等生物学过程来解释最大叶宽位置的遗传机理具备一定的可行性,但该性状独有的调控位点仍需利用QTL手段进行深入挖掘。
-
亲本丹红杨和通辽1号杨叶形性状差异显著,子代叶形多数为介于父母本的中间类型,变异丰富且呈连续的正态分布。10个表征叶片形状与大小的指标间存在广泛而显著的相关关系,其中,叶长和叶宽是决定叶面积的主要因素。杂交子代叶片形状与丹红杨更为接近,呈较强的母本效应,而叶片大小则表现出偏向父本小叶杨的遗传效应。部分子代(约30.09%)的综合叶形指标具有超亲现象,说明该杂交群体叶形性状的选择潜力较大。本研究利用422个杨属派间杂交子代无性系构成的家系内大群体对叶形性状的遗传变异规律进行了系统分析,丰富了杨树叶表型研究,为利用QTL手段挖掘相关候选基因、解析叶形性状的遗传控制机制奠定基础,也可为日后育种实践提供理论参考。
丹红杨×通辽1号杨杂交子代叶形性状的遗传变异分析
Genetic Variation Analysis of Leaf Morphological Traits in Populus deltoides cl. 'Danhong'×P. simonii cl. 'Tongliao 1' Hybrid Progenies
-
摘要:
目的 对丹红杨(美洲黑杨)×通辽1号杨(小叶杨)422个F1代无性系叶的形态学指标进行分析,揭示杨树派间杂交群体叶形性状的遗传变异规律,为进一步解析其叶形性状的遗传机制奠定基础。 方法 使用Yaxin-1241便携式叶面积仪结合手工测量的方式,对杂交子代当年扦插材料的叶面积、叶长、叶宽、叶周长、叶柄长、侧脉夹角和最大叶宽距叶尖长度等7个指标进行测量,计算叶长宽比、叶柄相对长、最大叶宽位置和叶缘因子,利用SPSS软件完成相关、通径、遗传以及主成分分析。 结果 丹红杨和通辽1号杨两亲本叶面积、叶长、叶宽、叶周长、叶柄长、侧脉夹角、叶长宽比、叶柄相对长以及最大叶宽位置等9个指标差异显著,子代变异丰富且呈连续的正态分布。在10个叶形指标间45对相关关系中,36对呈显著或极显著水平。通径分析结果表明,叶长和叶宽是决定叶面积的主要因素。通过主成分分析得到2个主因子,累计解释80.19%的叶形变异,子代第一主成分的标准化值全部介于两亲本之间,并有64.69%与父本分布在同侧,第二主成分则均匀分布在中亲值两侧。对于表征叶片大小的2个指标叶面积和叶柄长,多数子代小于中亲值且其均值更接近父本,而表征叶片形状的叶柄相对长和最大叶宽位置与之相反,其余指标接近中亲值。除叶长宽比、侧脉夹角以及叶缘因子外,子代叶形指标大多介于两亲本之间。 结论 丹红杨和通辽1号杨叶形性状差异显著,杂交子代叶形性状存在丰富的连续性变异,且性状间关联紧密,叶长和叶宽是决定叶面积的主要因素。叶面积、叶柄长以及最大叶宽位置等性状存在明显的遗传偏向性,杂交子代叶片形状与丹红杨更为接近,呈较强的母本效应,而叶片大小则表现出偏向父本小叶杨的遗传效应。30.09%子代的综合叶形指标具有超亲现象,选择潜力较大,在今后的育种实践中可加以利用。 Abstract:Objective To reveal the genetic variation pattern of leaf morphological traits in poplar intersectional hybrid population, and to lay the foundation for studying their genetic mechanisms by analyzing the leaf morphological indexes in 422 F1 progenies from P. deltoides cl. 'Danhong'×P. simonii cl. 'Tongliao 1'. Method Seven indexes (leaf area, length, width, perimeter, petiole length, vein angle and the distance between largest width and blade tip) were measured through Yaxin-1241 portable area meter and manual measurement. Four indexes (the ratio of leaf length and width, relative length of petiole, position of the largest width and leaf margin factor) were calculated. Correlation, path coefficient, genetic and principal component analysis were accomplished by SPSS software. Result Significant differences were found between two parents for all morphology indexes except leaf margin factor. Plentiful phenotypic variations with normal and continuous distribution existed in hybrid population. As many as 36 of 45 pairs of correlations which were up to significant or extremely significant level. The result of path coefficient analysis indicated that the leaf area was determined mainly by leaf length and width. Two main factors were obtained through principal component analysis, which explained 80.19% of total variations. Standardized value of first principal component in progenies was fallen completely between two parents, with 64.69% were distributed on the same side of male parent, while the second principal component was evenly distributed on both sides of the median parents. Two indexes of leaf size (leaf area and petiole length) in progenies were lower than the median of parents, and the mean was closer to male parent. In contrast, the leaf shape indexes (relative length of petiole and the position of maximum blade width) were closer to the female parent. Other indexes were closer to the median of parents. Most indexes of leaf morphology were between two parents except the ratio of length and width, the leaf margin factor and the lateral vein angle. Conclusion There are significant differences in leaf morphological traits between two parents. The variation of leaf morphological traits in their progenies is continuous and plentiful, and these traits are closely associated, among which leaf length and leaf width are the main factors determining leaf area. Leaf area, petiole length and position of the largest width have a evident genetic bias, the leaf shape of hybrid offspring which is more similar to P. deltoides cl 'Danhong' poplar shows a strong maternal effect, while the leaf size shows a genetic bias toward the male P. simonii cl. 'Tongliao 1' parental effect. The comprehensive leaf shape index of 30.09% progenies has the transgressive segregation phenomenon, which has great potential for selection, and could be utilized in tree breeding in the future. -
Key words:
- Populus
- / progenies
- / leaf morphological traits
- / genetic variation
-
图 3 F1代叶形性状的正态检验
Figure 3. Normality test of leaf morphological traits in F1 progenies
图 4 F1代叶形性状的频率分布
Figure 4. Frequency distribution of leaf morphological traits in F1 progenies
图 5 杂交群体叶形主成分2D点云图
Figure 5. Two-dimensional (2D) distributional scatter with principal component of leaf morphological traits in hybrid population
表 1 丹红杨和通辽1号杨叶形性状分析
Table 1. The analysis of leaf morphological traits of P. deltoides CL. 'Danhong' and P. simonii CL. 'Tongliao1'
性状
Traits丹红杨
P. deltoides CL.‘Danhong’通辽1号杨
P. simonii CL.‘Tongliao1’T检验(t值)
T-test (t value)显著性(P值)
Significance (P value)叶面积Leaf area (LA)/mm2 5 608.4±492.7 1 118.9±100.1 26.79 1.24×10-9 叶长Leaf length (LL)/mm 94.2±5.7 48.2±3.8 20.21 1.09×10-11 叶宽Leaf width (LW)/mm 86.2±2.7 34.3±0.6 57.24 1.20×10-12 叶柄长Petiole length (PL) /mm 62.1±6.2 5.5±1.8 26.22 4.98×10-10 叶周长Leaf perimeter (LP)/mm 282.1±16.7 125.7±7.0 25.87 5.11×10-11 侧脉夹角Lateral vein angle (LVA)/° 66.1±5.3 50.6±3.0 7.61 4.70×10-6 叶长宽比Ratio of length and width (LL/LW) 1.092±0.043 1.407±0.088 -9.59 7.26×10-7 最大叶宽位置Position of leaf width (LWP) 0.794±0.081 0.388±0.056 12.38 4.81×10-9 叶柄相对长Relative length of petiole (RPL) 0.661±0.077 0.113±0.032 19.76 8.29×10-10 叶缘因子Leaf margin factor (LMF) 0.282±0.019 0.283±0.008 -0.154 0.88  下载: 导出CSV
下载: 导出CSV
表 2 F1代叶形性状分布统计量
Table 2. The statistical analysis of leaf morphological traits in F1 progenies
性状
Traits均值
Mean value极小值
Minimal value极大值
Maximal value标准差
Standard deviation变异系数
Variable coefficient偏度
Skewness峰度
Kurtosis叶面积LA/mm2 2 780.291 1 071.233 6 063.233 757.538 0.272 0.814 1.080 叶长LL/mm 71.077 46.333 103.333 8.741 0.123 0.251 0.348 叶宽LW/mm 57.882 35.000 88.617 9.189 0.159 0.458 0.286 叶柄长PL/mm 31.681 12.667 57.500 6.825 0.215 0.545 0.698 叶周长LP/mm 195.330 120.133 291.617 25.593 0.131 0.412 0.797 侧脉夹角LVA/° 59.090 32.400 72.600 5.344 0.090 -0.890 2.436 叶长宽比LL/LW 1.252 0.767 2.084 0.174 0.139 0.496 1.261 最大叶宽位置LWP 0.641 0.412 0.923 0.055 0.086 0.012 1.925 叶柄相对长RPL 0.446 0.247 0.684 0.078 0.174 0.377 0.237 叶缘因子LMF 0.287 0.168 0.346 0.021 0.075 -0.507 1.729 注:变异系数=标准差/均值。
Note: Variable Coefficient = Standard Deviation / Mean Value.
下载: 导出CSV
表 3 F1代叶形性状的相关分析
Table 3. Correlation analysis of leaf morphological traits in F1 hybrid progenies
性状
Traits叶面积
LA叶周长
LP叶长
LL叶宽
LW叶柄长
PL侧脉夹
角LVA叶长宽比
LL/LW叶柄相对
长RPL最大叶宽
位置LWP叶缘因子
LMF叶面积LA 1.000 0.945** 0.785** 0.889** 0.523** 0.173** -0.316** 0.103* -0.188** 0.265** 叶周长LP 1.000 0.874** 0.806** 0.587** 0.107* -0.146** 0.117* -0.096* -0.038 叶长LL 1.000 0.553** 0.585** -0.060 0.256** 0.028 0.001 -0.224** 叶宽LW 1.000 0.428** 0.241** -0.649** 0.144** -0.162** 0.378** 叶柄长PL 1.000 -0.004 0.031 0.820** 0.204** -0.122* 侧脉夹角LVA 1.000 -0.328** 0.038 0.089 0.274** 叶长宽比LL/LW 1.000 -0.138** 0.191** -0.641** 叶柄相对长RPL 1.000 0.268** 0.013 最大叶宽位置LWP 1.000 -0.325** 叶缘因子LMF 1.000 注:*表示在0.05水平(双侧)显著相关;**表示在0.01水平(双侧)显著相关。
Note: *Indicates when P<0.05, there is significant correlation; **indicates when P<0.01, there is extremely significant correlation.
下载: 导出CSV
表 4 F1代4个叶片性状对叶面积的通径分析
Table 4. Path coefficient analysis of four leaf blade traits on leaf area in F1 progenies
性状
Traits相关系数
Correlation coefficient直接通径系数
Direct path coefficient间接通径系数Indirect path coefficient LL LW LVA LWD 总和(SUM) LL 0.785** 0.611 0.337 -0.037 0.508 0.808 LW 0.889** 0.604 0.334 0.146 0.218 0.697 LVA 0.173** 0.064 -0.004 0.015 2.555×10-4 0.011 LWD 0.537** -0.189 -0.157 -0.068 0.001 -0.224 注:*表示在0.05水平(双侧)显著相关;**表示在0.01水平(双侧)显著相关。
Note: *Indicates when P<0.05, there is significant correlation; **indicates when P<0.01, there is extremely significant correlation.
下载: 导出CSV
表 5 F1代4个叶片性状对叶面积的决定系数
Table 5. Determination coefficients of 4 leaf blade traits on leaf area in F1 progenies
性状
TraitsLL LW LVA LWD R2 剩余通径系数
Remanent path coefficient各性状对R2的贡献
Contribution to R2LL 0.374 0.408 -0.005 -0.192 0.926 0.272 0.479 LW 0.365 0.019 -0.082 0.537 LVA 0.004 9.647×10-5 0.011 LWD 0.036 -0.101
下载: 导出CSV
表 6 亲、子代叶形性状的遗传分析
Table 6. Genetic analysis of leaf morphological traits between F1 progenies and their parents
性状
Traits丹红杨
P.deltoides CL. ‘Danhong’通辽1号杨
P.simonii CL. ‘Tongliao1’中亲值
Mean value of parents子代均值
Mean value of progenies亲子代偏离度b
Degree of deviation between parents
and progenies大于中亲值子代比例
Percentage of progenies exceeded the mean value of parents/%超亲子代比例
Percentage of transgressive progenies/%叶面积LA/mm2 5 608.4 1 118.9 3 363.7 2 780.3 -0.260 20.62 0.47 叶长LL/mm 94.2 48.2 71.2 71.1 -0.004 49.05 1.66 叶宽LW/mm 86.2 34.3 60.3 57.9 -0.091 37.20 0.71 叶柄长PL/mm 62.1 5.5 33.8 31.7 -0.074 32.94 0.00 叶周长LP/mm 282.1 125.7 203.9 195.3 -0.110 32.70 0.71 侧脉夹角LVA/° 66.1 50.6 58.4 59.1 0.097 58.53 13.27 叶宽长比LW/LL a 0.916 0.711 0.813 0.799 -0.142 48.82 36.73 最大叶宽位置LWP 0.794 0.388 0.591 0.641 0.246 83.18 0.24 叶柄相对长RPL 0.661 0.113 0.387 0.446 0.215 77.73 0.95 叶缘因子LMF 0.282 0.283 0.283 0.287 -9.000 57.58 100.00 注:a:为便于比较,采用叶长宽比的倒数来表示其相对值;b:亲子代偏离度=(子代均值-中亲值)/(较大亲本值-中亲值)。
Note: a:To make the comparison more convenience, we use the reciprocal of length-width ratio for its relative value.
b:Divergence between F1 progenies and their parents = (Mean value of F1 progenies-Mid-parents value) /(Larger value of parents-Mid-parents value).
下载: 导出CSV
表 7 F1代叶形性状的主成分特征向量
Table 7. Principal component eigenvectors of leaf morphological traits in F1 progenies
性状
Traits第一主成分
The first principal component第二主成分
The second principal component特征值Eigenvalue 4.459 1.154 累计贡献率/%
Accumulated contribution rate63.71 80.19 叶面积LA 0.859 0.413 叶周长LP 0.926 0.279 叶长LL 0.942 -0.029 叶宽LW 0.700 0.554 叶柄长PL 0.761 -0.043 侧脉夹角LVA -0.070 0.880 最大叶宽距叶尖长度LWD 0.842 -0.123
下载: 导出CSV
-
[1] Nicotra A B, Leigh A, Boyce C K, et al. The evolution and functional significance of leaf shape in the angiosperms[J]. Functional Plant Biology, 2011, 38(7):535-552. doi: 10.1071/FP11057 [2] Du Q, Xu B, Gong C, et al. Variation in growth, leaf, and wood property traits of Chinese white poplar (Populus tomentosa), a major industrial tree species in Northern China[J]. Canadian Journal of Forest Research, 2014, 44(4):326-339. doi: 10.1139/cjfr-2013-0416 [3] Wang H, Niu L, Fu C, et al. Overexpression of the WOX gene STENOFOLIA improves biomass yield and sugar release in transgenic grasses and display altered cytokinin homeostasis[J]. Plos Genetics, 2017, 13(3):e1006649. doi: 10.1371/journal.pgen.1006649 [4] 胡建军.美洲黑杨叶面积、生长量、酚类物质及抗虫性状基因定位[D].北京: 北京林业大学, 2002. [5] Bresso E G, Chorostecki U, Rodriguez R E, et al. Spatial control of gene expression by miR319-regulated TCP transcription factors in leaf development[J]. Plant Physiol, 2018, 176:1694-1708.. doi: 10.1104/pp.17.00823 [6] Feng G, Qin Z, Yan J, et al. Arabidopsis ORGAN SIZE RELATED1 regulates organ growth and final organ size in orchestration with ARGOS and ARL[J]. New Phytologist, 2011, 191(3):635-646. doi: 10.1111/nph.2011.191.issue-3 [7] Zhang J, Wei B, Yuan R, et al. The Arabidopsis RING-type E3 ligase TEAR1 controls leaf development by targeting the TIE1 transcriptional repressor for degradation[J]. Plant Cell, 2017, 29(2):243-259. doi: 10.1105/tpc.16.00771 [8] Farooq M, Tagle A G, Santos R E, et al. Quantitative trait loci mapping for leaf length and leaf width in rice cv.IR64 derived lines[J]. Journal of Integrative Plant Biology, 2010, 52(6):578-584. doi: 10.1111/jipb.2010.52.issue-6 [9] Wassom J J. Quantitative trait loci for leaf angle, leaf width, leaf length, and plant height in a maize (Zea mays L) B73×Mo17 population[J]. Maydica, 2013, 58(3):318-321. [10] 闫雪, 史雨刚, 梁增浩, 等.小麦旗叶形态相关性状的QTL定位[J].核农学报, 2015, 29(7):1253-1259. [11] 魏永成, 李周岐, 李煜, 等.杜仲杂交子代苗期表型性状的遗传分析[J].西北农林科技大学学报:自然科学版, 2012, 40(8):137-143. [12] Tan L Q, Wang L Y, Xu L Y, et al. SSR-based genetic mapping and QTL analysis for timing of spring bud flush, young shoot color, and mature leaf size in tea plant (Camellia sinensis)[J]. Tree Genetics & Genomes, 2016, 12(3):52. [13] 苏晓华, 李金花, 陈伯望, 等.杨树叶片数量性状相关联标记及其图谱定位研究[J].林业科学, 2000, 36(1):33-40. [14] Trewin H. Linking transcript, QTL and association mapping to understand the genetic control of leaf size and shape in Populus[D]. University of Southampton, 2008. [15] Migicovsky Z, Li M, Chitwood D H, et al. Morphometrics reveals complex and heritable apple leaf shapes[J]. Frontiers in Plant Science, 2018, 8, No.2185: 1-16. [16] Drost D R, Puranik S, Novaes E, et al. Genetical genomics of Populus leaf shape variation[J]. BMC Plant Biology, 2015, 15(1), No.166: 1-10. [17] Thomas H, Ougham H. On plant growth and form.[J]. New Phytologist, 2017, 216(2):337-338. doi: 10.1111/nph.14807 [18] 李慧慧, 张鲁燕, 王建康.数量性状基因定位研究中若干常见问题的分析与解答[J].作物学报, 2010, 36(6):918-931. [19] 曾燕蓉, 朱方容, 林强, 等.几种桑树叶面积测量方法及叶片大小与叶片质量的相关性分析[J].蚕业科学, 2013(5):868-876. [20] Perez-Sanz F, Navarro P J, Egea-Cortines M. Plant phenomics:an overview of image acquisition technologies and image data analysis algorithms[J]. Gigascience, 2017, 6(11):1-18. [21] 李金花, 张绮纹, 苏晓华, 等.美洲黑杨与不同种源青杨杂种苗叶片和生长性状多水平变异研究[J].林业科学研究, 2002, 15(1):76-82. doi: 10.3321/j.issn:1001-1498.2002.01.012 [22] 张春玲, 李淑梅, 赵自成, 等.杨树新品种'丹红杨[J].林业科学, 2008, 44(1):169. doi: 10.3321/j.issn:1001-7488.2008.01.027 [23] 徐纬英.杨树[M].哈尔滨:黑龙江人民出版社, 1988. [24] 姬慧娟.丹红杨与小叶杨杂交子代苗期抗旱相关性状遗传分析[D].北京: 中国林业科学研究院, 2015. [25] Dewey D R, Lu K H. A Correlation and Path-Coefficient Analysis of Components of Crested Wheatgrass Seed Production[J]. Problemy Tuberkuleza, 1959, 51(9):70-74. [26] El-Kassaby Y A, Ritland K, Ritland C, et al. Forest Genetics[M]//Watts S B, Tolland L. Faculty of Forestry. Vancouver, BC: The university of British Columbia, 2005: 474-486. [27] 张福敏.响叶杨×银白杨遗传图谱加密与相关QTL定位[D].南京: 南京林业大学, 2011. [28] Baker R L, Leong W F, Brock M T, et al. Modeling development and quantitative trait mapping reveal independent genetic modules for leaf size and shape[J]. New Phytologist, 2015, 208(1):257-268. doi: 10.1111/nph.2015.208.issue-1 [29] Tsukaya H. Leaf shape:genetic controls and environmental factors[J]. International Journal of Developmental Biology, 2004, 49(5-6):547-555. [30] Kozuka T, Horiguchi G, Kim G T, et al. The different growth responses of the Arabidopsis thaliana Leaf Blade and the Petiole during Shade Avoidance are Regulated by Photoreceptors and Sugar[J]. Plant & Cell Physiology, 2005, 46(1):213-223. [31] Guet J, Fabbrini F, Fichot R, et al. Genetic variation for leaf morphology, leaf structure and leaf carbon isotope discrimination in European populations of black poplar (Populus nigra L.)[J]. Tree Physiology, 2015, 35(8):850. doi: 10.1093/treephys/tpv056 [32] 徐红, 韩玉兰, 杨自湘, 等.用叶片特征区别不同产地不同单株青杨的研究[J].林业科技通讯, 1995(2):17-18. [33] 薄文浩.藏川杨遗传多样性及杂交子代遗传变异研究[D].北京: 北京林业大学, 2012. [34] 张德强, 张志毅, 杨凯, 等.毛新杨×毛白杨叶片表型和春季萌芽时间QTL分析(英文版)[J].林业科学, 2005, 41(1):42-48. doi: 10.3321/j.issn:1001-7488.2005.01.009 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 4772
- HTML全文浏览量: 1683
- PDF下载量: 155
- 被引次数: 0
