


-
植物体内含有多种微生物,其中许多微生物与植物之间的相互作用对植物的生存、适应性和生态系统功能等方面都有有益的影响[1-2],大量研究表明内生菌在提高植物免疫防御[3]、病害控制[4]、营养获取[5]和植物对非生物胁迫的耐受方面[6]有明显的促进作用。比如Wu等人[7]从野生蓝莓的内生菌中分离到一株深色有隔真菌,将该菌接种到蓝莓上可促进其根部生长和分枝,Carro-Huerga等人[8]从葡萄树体里分离到一株木霉,该菌可减少葡萄上病原菌的定殖从而限制病害的发展。
松材线虫(Bursaphelenchus xylophilus)是全球森林生态系统中最具危险性、毁灭性的有害生物之一,具有极强的扩散性和破坏性,自1982年入侵我国后,先后在亚热带、热带、暖温带、中温带以及秦岭等高海拔地区造成松林大面积枯死[9],造成了巨大的经济和生态损失,而且疫区仍在不断增加。松材线虫侵入寄主后,首先取食松树的薄壁细胞,当松树死亡后,开始取食树体内的真菌,使得其种群得以维持和扩大[10];松树内生微生物群落具有提高树木防御能力的潜力,但也可能随着病害的发展而转换角色,最终导致树木腐烂[11]。由此可见,松树内生微生物在松材线虫病的发展过程中发挥重要作用。充分了解松树体内微生物的群落结构有助于了解松材线虫病的发病过程,也为认识寄主和微生物的共生关系以及微生物资源的开发利用提供参考和科学依据。
许多研究结果表明松材线虫入侵对寄主松树的内生微生物群落多样性具有重要影响,而且松树的许多内生菌也具有很大的生防潜力。曾凡勇等[12]对不同健康状态黑松(Pinus thunbergii Parl.)树干内真菌进行了分离,发现不同健康状态下真菌种群没有显著差异,但是优势真菌及其比率不同。鲁国华[13]分析了不同健康状态马尾松(P. massoniana Lamb.)和黑松树干内真菌的种类变化,认为松材线虫病影响了寄主松树真菌种群的变化。徐风美[14]通过对松材线虫病发生区和未发生区油松(P. tabuliformis Carr.)根部真菌进行分离和鉴定,发现根部外生菌根真菌和深色有隔内生真菌的相对丰度不同,这可能与松树对松材线虫病的抗性有关。Ma等人[15]利用高通量测序技术研究了松材线虫侵染前后黑松不同部位的细菌群落结构,发现健康树和受害树针叶和根部细菌的群落结构不同,表明松材线虫可以影响寄主的细菌群落结构。Ponpandian等人[16]从赤松(P. densiflora Sieb. et Zucc.)、刚松(P. rigida Mill.)、黑松和红松(P. koraiensis Sieb. et Zucc.)不同组织部位中分离得到1622株内生细菌,从中筛选到44株具有杀线活性的细菌,其中窄食单胞菌属(Stenotrophomonas)和芽孢杆菌属(Bacillus)细菌的杀线作用最突出。Kim等人[17]从分离到的92株松树内生细菌中筛选出3株可诱导松树产生抗性的细菌,通过叶面喷洒这3株细菌的菌悬液均可显著降低松材线虫病的发病程度。
马尾松作为我国南方重要的绿化造林树种,近年来松材线虫病在马尾松林普遍发生,但关于马尾松不同组织部位微生物群落多样性的认识还不全面,对马尾松内生微生物的种类与功能缺乏深入的研究,这就阻碍了防控研究的进一步开展。本研究通过对松材线虫病疫区马尾松不同部位微生物的分离,有助于了解松材线虫入侵前后马尾松体内微生物的群落结构,同时可构建马尾松上可培养的微生物种质资源库,为利用内生微生物防治松材线虫病,探索松材线虫病的新型防控技术奠定基础。
-
于2019年9月在浙江省富阳区(30°3′N, 119°58′E)和淳安县(29°36′N, 118°58′E)采集健康马尾松和由松材线虫侵染引起的枯死马尾松各4棵,枯死马尾松均为当年枯死树,经贝尔曼漏斗法分离后每克木材中松材线虫数量大于100头,健康马尾松中没有分离到松材线虫,枯死树中均分离到松材线虫,其中3棵健康马尾松和2棵枯死马尾松来自富阳区,1棵健康马尾松和2棵枯死马尾松来自淳安县。所采集的马尾松均为25~30年生,胸径为17~22 cm,树高为16~20 m。分别采集马尾松地下部分10 cm深处的侧根、地上部分主干的上、中、下3段和2年生针叶,编号后封存于保鲜袋。
-
植物基因组DNA提取试剂盒、细菌基因组DNA提取试剂盒、2 × Taq PCR MasterMix,D2000 DNA Marker购自天根生化科技(北京)有限公司;PCR引物对ITS1/ITS4和27F/1492R由北京六合华大基因科技有限公司合成。
-
采用常规组织分离法分离马尾松不同部位的真菌和细菌。侧根处理前先将根表面的土用无菌水冲洗掉,再用灭菌滤纸擦干后备用。主干处理前先将粗糙的表皮刮掉。所有样品用75%的乙醇浸泡1分钟后用无菌水冲洗3次,取最后1次的冲洗液涂布于PDA培养基(200 g去皮马铃薯,20 g葡萄糖,15 g琼脂粉,去离子水1000 mL)和牛肉膏蛋白胨培养基(NA)(蛋白胨10 g,氯化钠5 g,牛肉浸出粉3 g,琼脂15 g,去离子水1 000 mL)平板做表面消毒效果检测。用无菌滤纸擦干组织表面的水分,用无菌枝剪剪去主干和根经酒精处理过的表面组织,再将剩余的主干和根剪碎成约3 mm × 3 mm × 2 mm的木块,将针叶剪成约6 mm长的片段,用于后续分离。
-
用无菌镊子将剪碎的组织块置于90 mm的PDA培养基上,每个样品接种25个组织块,健康和枯死马尾松各4棵,根、干和针叶各200块,共600个组织块,置于25℃培养箱中黑暗培养,待其长出菌落后用无菌接种针挑取菌落边缘气生菌丝置于PDA培养基上纯化培养。将菌落特征相同的菌株进行归类统计,将分离纯化后的不同菌落形态的菌株接种于PDA培养基上,并对菌株进行编号,保存备用。
-
用无菌镊子夹取1 g剪碎的组织块,置于5 mL的无菌水中,28℃,200 r· min−1振荡培养1 h,混匀,吸取100 μL培养液涂布于NA培养基上,28℃培养24 h,挑取单菌落至NA培养基上纯化,挑取纯化后的细菌单菌落至牛肉膏蛋白胨液体培养基(NB)(蛋白胨10 g,氯化钠5 g,牛肉浸出粉3 g,去离子水1000 mL)中,28 ℃,200 r· min−1培养24 h,4℃保存备用。
-
将1.4.1保存的菌株重新接种于PDA培养基,25℃培养7 d,挑取菌丝于2 mL离心管中,采用植物基因组DNA提取试剂盒(DP320)提取菌株DNA。对于菌液PCR无条带的细菌菌株进行DNA提取。取1.4.2中细菌菌液1 mL于1.5 mL离心管中,6000 r· min−1离心1 min,弃去上清液,采用细菌基因组DNA提取试剂盒(DP302)提取菌株的总DNA。
-
采用真菌核糖体rDNA区通用引物ITS1/ITS4(5′-TCCGTAGGTGAACCTGCGG-3′/5′-TCCTCCGCTTATTGATATGC-3′)进行PCR扩增。PCR反应体系25 μL包含:基因组DNA 1 μL,2 × Taq PCR MasterMix(KT201) 12.5 μL,引物ITS1 0.5 μL,引物ITS4 0.5 μL,ddH2O 10.5 μL。扩增程序:94℃预变性3 min;94℃变性30 s,55℃退火30 s,72℃延伸45 s,35个循环;72℃延伸10 min。
采用细菌16S rDNA区通用引物27F/1492R(5′-AGAGTTTGATCCTGGCTCAG-3′/5′-TACGGTTACCTTGTTACGACTT-3′)进行PCR扩增。利用4℃保存的菌液或基因组DNA作为PCR模板,PCR反应体系50 μL包含:菌液或基因组DNA2 μL,2 × Taq PCR MasterMix(KT201)25 μL,引物27F 1 μL,引物1492R 1 μL,ddH2O 21 μL。扩增程序:94℃预变性5 min;94℃变性30 s,55℃退火30 s,72℃延伸90 s,35个循环;72℃延伸10 min。
-
取5 μL PCR产物用1.0%的琼脂糖凝胶电泳进行检测,将剩余的PCR产物送至北京六合华大基因科技有限公司测序,对拼接后的序列利用BLAST软件进行序列相似性比对。结合本研究分离到的木霉菌株,选取部分代表菌株的ITS序列,利用MEGA 6.0软件的邻接法构建系统发育进化树,自展值(Bootstrap replications)为1000次。
-
将本研究分离到的6种木霉属(Trichoderma)真菌分别接种至PDA平板上,于25℃恒温培养7 d。将实验室保存的松材线虫虫株Nxy61接至培养7 d的灰葡萄孢平板并置于25℃恒温培养,7 d后用无菌水将松材线虫从平板洗下,用无菌水重复换洗3遍后计数。分别吸取约3000条松材线虫接种于培养7 的木霉和灰葡萄孢(Botrytis cinerea Pers.)平板上,于25℃恒温培养7 d,7 d后用无菌水完全冲洗平板,通过光学显微镜统计活体松材线虫的种群数量。每个处理5个重复,以灰葡萄孢培养松材线虫作为对照。
-
采用分离率和分离频率分析马尾松不同部位分离的真菌丰富程度及优势菌群,采用Shannon-Wiener指数(H)、Simpson指数(D)、均匀度指数(J)分析分离真菌的多样性特征,用Sorenson指数(Cs)和Jaccard指数(Cj)分析不同组织和不同健康状态马尾松的真菌种类组成的相似程度[18]。
$ H=-\sum _{i=1}^{s}{P}_{i}\mathrm{ln}{P}_{i}, $
$ D=\sum {{P}_{i}}^{2}, $
$ J=-\sum _{i=1}^{s}{P}_{i}\mathrm{ln}{P}_{i}/\mathrm{ln}S, $
$ {C}_{s}=\frac{2j}{a + b}, $
$ {C}_{j}=\frac{j}{a + b-j}。 $
式中:Pi为第i种的个体数占总物种数S的比例。
j为两个组织或样本共有的真菌种数,a和b分别为每个组织或样本中的真菌种数。
利用SPSS 22.0对松材线虫取食不同真菌后的种群数量进行单因素方差分析及Duncan’s多重比较(p<0.05)。
-
对浙江马尾松进行内生真菌分离,经纯化后分离得到健康马尾松内的真菌106株,其中干部的分离率最高,为55%(55株),其次为针叶和根部,分离率分别为46%(46株)和5%(5株)。经过ITS序列比对将分离菌株划分为19个属,木霉属(Trichoderma)为优势类群,占菌株总数的25.47%,而且不同部位的优势类群不同,根部优势属为毛色二孢属(Lasiodiolodia),干部为木霉属(Trichoderma),针叶为拟盾壳霉属(Paraconiothyrium)(表1)。枯死马尾松中分离得到真菌86株,其中针叶的分离率最高,为44%(44株),其次是干部和根部,分离率分别为32%(32株)和10%(10株)。经过ITS序列比对将分离菌株划分为10个属,木霉属(Trichoderma)也为优势类群,占菌株总数的34.88%,分不同部位来看,根部和干部的优势属为木霉属(Trichoderma),针叶为散斑壳属(Lophodermium)(表2)。
表 1 健康马尾松不同部位内生真菌的组成及分离频率
Table 1. The fungal composition and isolation frequency of different tissues in healthy Masson pine
编号
No.属 Genus 根 Root 干 Stem 叶 Needle 菌株总数
Total number of
strains相对频率
Relative
frequency/%菌株数量
Number of
strains分离频率
Frequency/
%菌株数量
Number of
strains分离频率
Frequency/
%菌株数量
Number of
strains分离频率
Frequency/
%1 木霉属
Trichoderma0 0 23 21.70 1 0.94 24 22.64 2 链格孢属
Alternaria0 0 2 1.89 5 4.72 7 6.60 3 小枝孢属
Brachysporiella0 0 2 1.89 0 0 2 1.89 4 青霉属
Penicillium1 0.94 7 6.60 0 0 8 7.55 5 散斑壳属
Lophodermium0 0 0 0 7 6.60 7 6.60 6 拟盘多毛孢属
Pestalotiopsis0 0 6 5.66 3 2.83 9 8.49 7 短柄霉属
Aureobasidium0 0 1 0.94 0 0 1 0.94 8 炭疽菌属
Colletotrichum0 0 0 0 8 7.55 8 7.55 9 拟盾壳霉属
Paraconiothyrium0 0 0 0 19 17.92 19 17.92 10 毛色二孢属
Lasiodiolodia4 3.78 0 0 0 0 4 3.77 11 被孢霉属
Mortierella0 0 3 2.83 0 0 3 2.83 12 伞状霉属
Umbelopsis0 0 5 4.72 0 0 5 4.72 13 炭团菌属
Hypoxylon0 0 1 0.94 0 0 1 0.94 14 亚罗酵母属
Yarrowia0 0 1 0.94 0 0 1 0.94 15 皮司霉属
Pithomyces0 0 2 1.89 0 0 2 1.89 16 黑孢属
Nigrospora0 0 0 0 2 1.89 2 1.89 17
Kirschsteiniothelia0 0 1 0.94 0 0 1 0.94 18 Eurotiomycetes 0 0 1 0.94 0 0 1 0.94 19
Atractidochium0 0 0 0 1 0.94 1 0.94 合计 Total 5 4.72 55 51.88 46 43.40 106 1 香农-威纳指数/H Shannon-Wiener index/H 0.50 1.81 1.68 2.37 辛普森指数/D Simpson index/D 0.32 0.78 0.76 0.87 均匀度指数/J Evenness index/J 0.31 0.45 0.44 0.51 表 2 枯死马尾松不同部位内生真菌的组成及分离频率
Table 2. The fungal composition and isolation frequency of different tissues in dead Masson pine
编号
No.属
Genus根 Root 干 Stem 叶 Needle 菌株总数
Total number
of strains相对频率
Relative
frequency/%菌株数量
Number of
strains分离频率
Frequency/
%菌株数量
Number of
strains分离频率
Frequency/
%菌株数量
Number of
strains分离频率
Frequency/
%1 木霉属
Trichoderma8 9.30 22 25.58 0 0 30 34.88 2 链格孢属
Alternaria0 0 1 1.16 1 1.16 2 2.33 3 半帚孢属
Leptographium0 0 4 4.65 0 0 4 4.65 4 小枝孢属
Brachysporiella0 0 1 1.16 0 0 1 1.16 5 青霉属
Penicillium2 2.33 0 0 0 0 2 2.33 6 长喙壳属
Ophiostoma0 0 4 4.65 0 0 4 4.65 7 散斑壳属
Lophodermium0 0 0 0 27 31.40 27 31.40 8 拟盘多毛孢属
Pestalotiopsis0 0 0 0 10 11.63 10 11.63 9 微色二孢属
Microdiplodia0 0 0 0 3 3.49 3 3.49 10 短柄霉属
Aureobasidium0 0 0 0 3 3.49 3 3.49 合计 Total 10 11.63 32 37.21 44 51.16 86 1 香农-威纳指数/H Shannon-Wiener index/H 0.50 0.99 1.09 1.73 辛普森指数/D Simpson index/D 0.32 0.49 0.56 0.76 均匀度指数/J Evenness index/J 0.22 0.29 0.29 0.39 -
对浙江马尾松进行内生细菌的分离,经纯化后分离得到健康马尾松内的细菌59株,经过序列比对将分离菌株划分为13个属,其中伯克氏菌属(Burkholderia)18株为优势菌群,占分离细菌总数的30.51%;沙雷氏菌属(Serratia)和肠杆菌属(Enterobacter)各10株,占16.95%;泛菌属(Pantoea)4株,占6.78%;其它细菌占28.81%。根据不同部位的分离情况,根部、干部和针叶的优势菌群分别是伯克氏菌属(Burkholderia)、肠杆菌属(Enterobacter)和沙雷氏菌属(Serratia)(表3)。从枯死马尾松中分离得到细菌56株,经过序列比对将分离菌株划分为12个属,其中伯克氏菌属(Burkholderia)22株为优势菌群,占分离细菌总数的39.29%;肠杆菌属(Enterobacter)8株,占14.29%;沙雷氏菌属(Serratia)6株,占10.71%;芽孢杆菌属(Bacillus)5株,占8.93%;其它细菌占26.78%。根据不同部位的分离情况,根部和干部的优势菌群分别是伯克氏菌属(Burkholderia)和肠杆菌属(Enterobacter)(表4)。
表 3 健康马尾松不同部位内生细菌的组成及分离频率
Table 3. The bacterial composition and isolation frequency of different tissues in healthy Masson pine
编号
No.属 Genus 根 Root 干 Stem 叶
Needle菌株总数
Total number
of strains相对频率
Relative
frequency/%菌株数量
Number of
strains分离频率
Frequency/
%菌株数量
Number of
strains分离频率
Frequency/
%菌株数量
Number of
strains分离频率
Frequency/
%1 伯克氏菌属 Burkholderia 15 25.42 3 5.08 0 0 18 30.51 2 沙雷氏菌属 Serratia 1 1.69 7 11.86 2 3.39 10 16.95 3 肠杆菌属 Enterobacter 0 0 9 15.25 1 1.69 10 16.95 4 拉乌尔菌属 Raoultella 1 1.69 2 3.39 0 0 3 5.08 5 泛菌属 Pantoea 0 0 4 6.78 0 0 4 6.78 6 欧文氏菌属 Erwinia 0 0 1 1.69 0 0 1 1.69 7 杆菌属 Bacterium 0 0 2 3.39 1 1.69 3 5.08 8 假单胞菌属 Pseudomonas 3 5.08 0 0 0 0 3 5.08 9 鞘氨醇单胞菌属 Sphingomonas 1 1.69 1 1.69 0 0 2 3.39 10 克吕沃尔氏菌属 Kluyvera 0 0 1 1.69 0 0 1 1.69 11 类芽孢杆菌属 Paenibacillus 0 0 1 1.69 0 0 1 1.69 12 Paraburkholderia 0 0 1 1.69 0 0 1 1.69 13 Uncultured bacterium 0 0 2 3.39 0 0 2 3.39 合计 Total 21 35.59 34 57.63 4 6.78 59 1 表 4 枯死马尾松不同部位内生细菌的组成及分离频率
Table 4. The bacterial composition and isolation frequency of different tissues in dead Masson pine
编号
No.属
Genus根 Root 干 Stem 叶 Needle 菌株总数
Total number
of strains相对频率
Relative
frequency/%菌株数量
Number of
strains分离频率
Frequency/
%菌株数量
Number of
strains分离频率
Frequency/
%菌株数量
Number of strains分离频率
Frequency/
%1 伯克氏菌属 Burkholderia 19 33.93 3 5.36 0 0 22 39.29 2 沙雷氏菌属 Serratia 2 3.57 4 7.14 0 0 6 10.71 3 芽孢杆菌属 Bacillus 3 5.36 0 0 2 3.57 5 8.93 4 肠杆菌属 Enterobacter 0 0 8 14.29 0 0 8 14.29 5 果胶杆菌属 Pectobacterium 0 0 3 5.36 0 0 3 5.36 6 拉乌尔菌属 Raoultella 0 0 1 1.79 0 0 1 1.79 7 泛菌属 Pantoea 0 0 3 5.36 0 0 3 5.36 8 欧文氏菌属 Erwinia 0 0 1 1.79 0 0 1 1.79 9 克雷伯氏菌属 Klebsiella 0 0 1 1.79 0 0 1 1.79 10 杆菌属 Bacterium 0 0 2 3.57 2 3.57 4 7.14 11 Paraburkholderia 0 0 1 1.79 0 0 1 1.79 12 Uncultured bacterium 1 1.79 0 0 0 0 1 1.79 合计 Total 25 44.64 27 48.21 4 7.14 56 1 -
由表1可知,健康马尾松不同组织间的真菌分布存在较大差异,干部分离到的真菌有13个属,种类最多,Shannon-Wiener指数(H)为1.81,针叶分离的真菌有8个属,H为1.68,根部分离的真菌只有2个属,H为0.50。Simpson指数(D)以干部最高,为0.78,根部最低,为0.32;均匀度指数(J)以干部最高,为0.45,根部最低,为0.31。与健康马尾松相比,枯死马尾松不同组织分离到的真菌种类较少,干部和针叶都分离到5个属的真菌,H分别为0.99和1.09。Simpson指数(D)以针叶最高,为0.56,根部最低,为0.32;干部和针叶的均匀度指数(J)相同,为0.29,根部最低,为0.22(表2)。
从表5可看出,健康和枯死马尾松根叶的相似性均为0,健康马尾松干叶的相似性最高,枯死马尾松根干的相似性最高,但指数值均小于0.5,表明健康和枯死马尾松不同组织间的真菌菌群相似性都较低。
表 5 健康和枯死马尾松不同部位真菌菌群相似性系数
Table 5. The similarity indexes of fungal community of different tissue from healthy and dead Masson pine
健康马尾松
Healthy Masson pine枯死马尾松
Dead Masson pine根 Root 干S tem 针叶 Needle 根 Root 干 Stem 针叶 Needle 根 Root 0.07 0.00 0.17 0.00 干 Stem 0.13 0.17 0.29 0.11 针叶 Needle 0.00 0.29 0.00 0.2 注:表中对角线以上为Jaccard指数,以下为Sorenson指数。
The values above the diagonal in the table were Jaccard index, and the values below were Sorenson index.由表1和表2可知,健康马尾松的真菌多样性高于枯死马尾松,Shannon-Wiener指数(H)、Simpson指数(D)和均匀度指数(J)依次为2.37、0.87和0.51,均高于枯死马尾松的1.72、0.76和0.39。从相似性指数来看,健康和枯死马尾松的真菌菌群之间相似性较低,Sorenson指数(CS)和Jaccard指数(Cj)分别为0.48和0.32,均小于0.5。
-
由表3可知,健康马尾松不同组织间的细菌分布存在较大差异,干部分离到11个属(32株)的细菌,种类最多,其次是根部的5个属(21株),最少的是针叶(4株),只分离到3个属的细菌。在枯死马尾松中,也是干部分离到的细菌种类最多,有10个属(27株),根部和针叶分别分离到3个属(24株)和2个属(4株)的细菌(表4)。健康和枯死马尾松中均为干部的细菌多样性最高,针叶的细菌多样性最低。
健康和枯死马尾松中共有的细菌种类有8个属,其中干部共有的类群有8个属,根部有2个,针叶有1个,Sorenson指数(CS)和Jaccard指数(Cj)分别为0.70和0.53,表明健康和枯死马尾松的细菌群落具有一定的相似性。
-
本研究共分离到6种木霉,在6 cm培养皿内的PDA培养基上培养4 d后菌落形态见图1。以Protocrea farinosa为外族群,6种木霉的系统发育进化树结果见图2。6个种类的木霉在发育树中归于3个进化支,FXY7与T. harzianum亲缘关系最近,FXY8与T. spirale亲缘关系最近,FXY37归于longibrachiatum分支,FXY19、FXY32和FXY56这3种木霉归于同一分支。

图 1 6种木霉的菌落形态
Figure 1. Colony morphology of six Trichoderma species
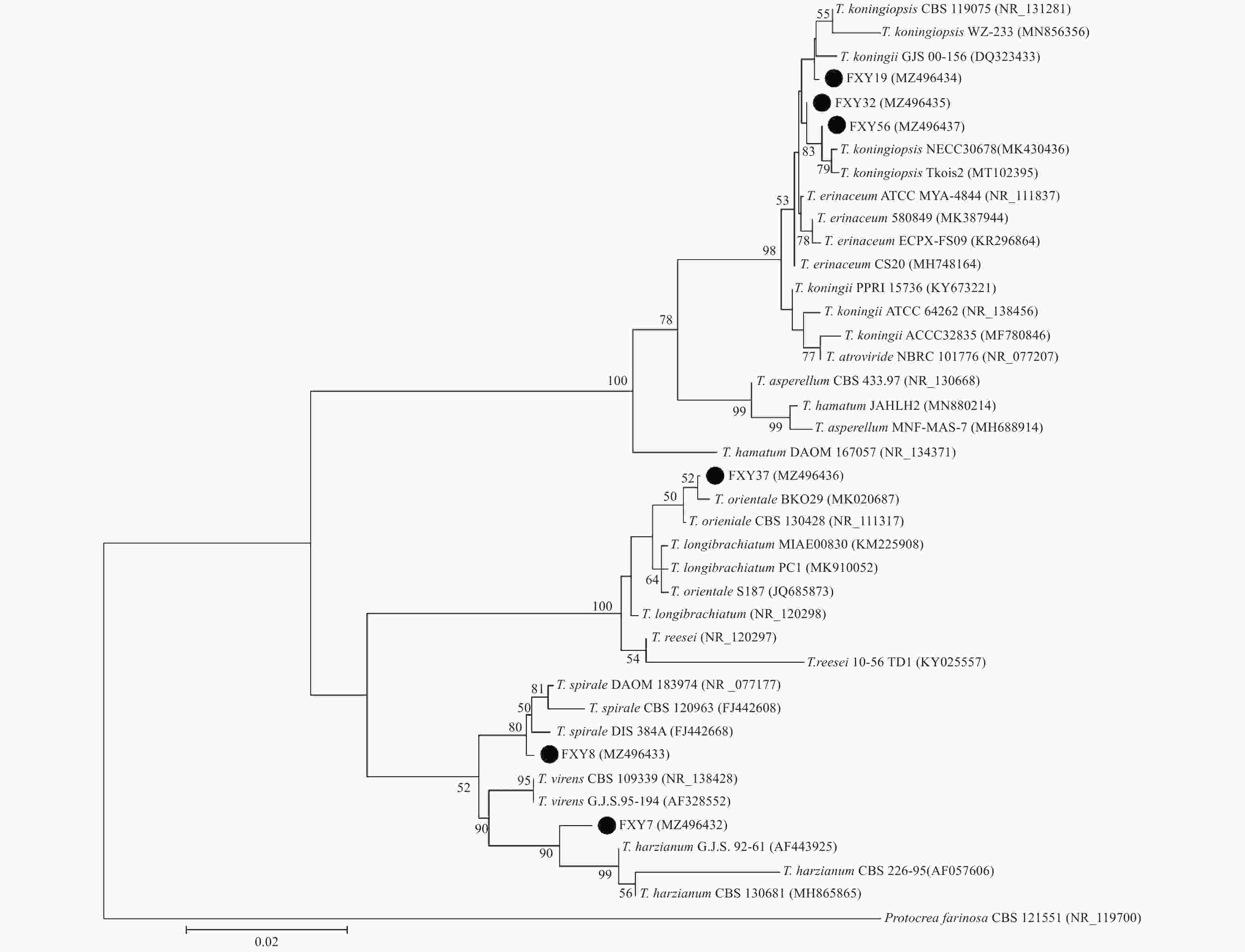
图 2 基于ITS序列构建的木霉系统发育进化树
Figure 2. Phylogenetic tree of Trichoderma strains inferred from ITS gene sequences
-
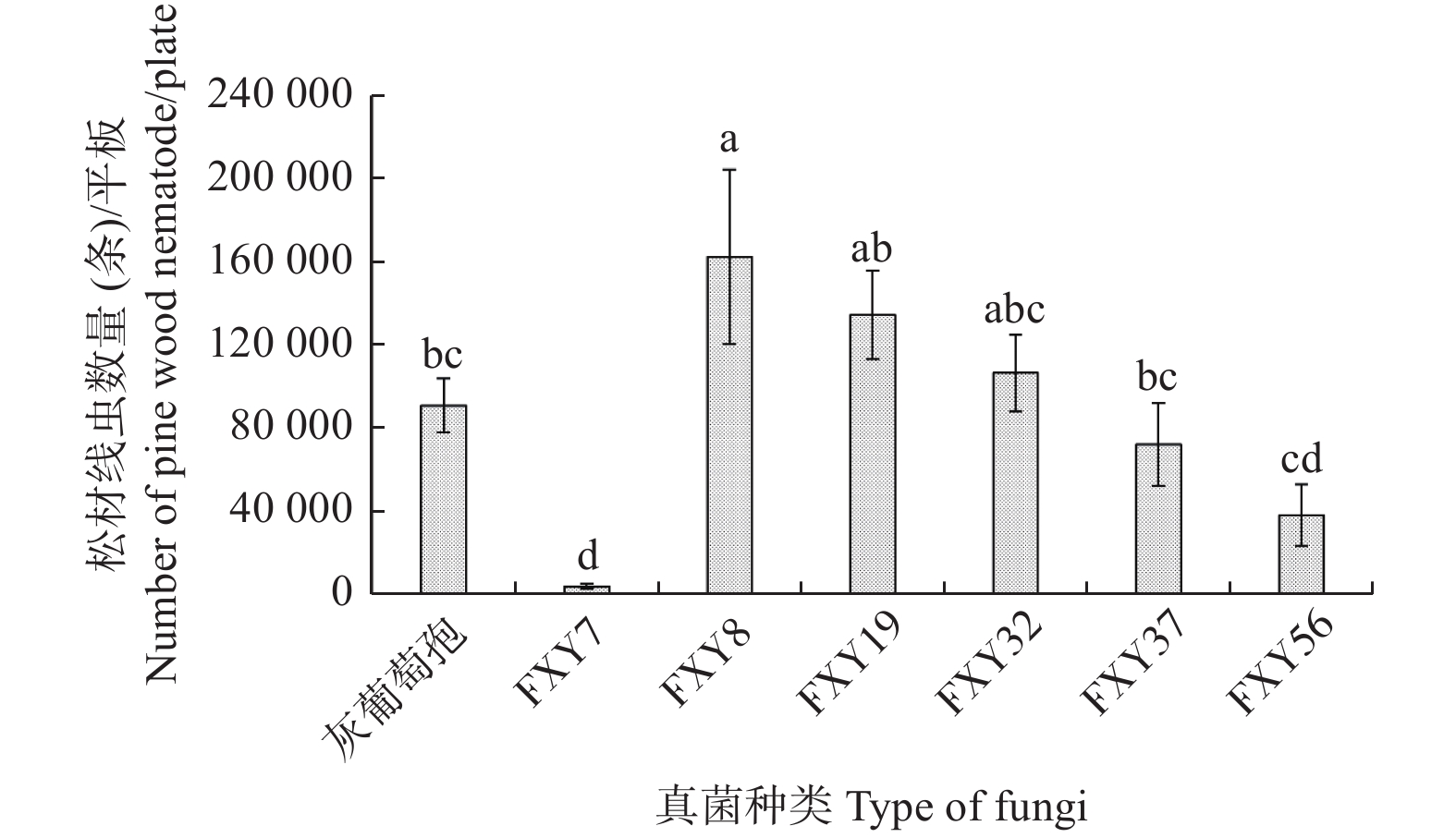
由图3可知,不同木霉培养下的松材线虫种群数量具有显著差异,取食木霉FXY8的松材线虫数量最多,达到162300条,显著高于灰葡萄孢培养的松材线虫数量,取食木霉FXY7的松材线虫数量最少,只有4275条,最不利于松材线虫繁殖(F = 6.21,df = 6, 28,p<0.05)。其它4种木霉的培养效果与灰葡萄孢没有显著差异,但用FXY19培养得到的线虫数量明显多于FXY56。
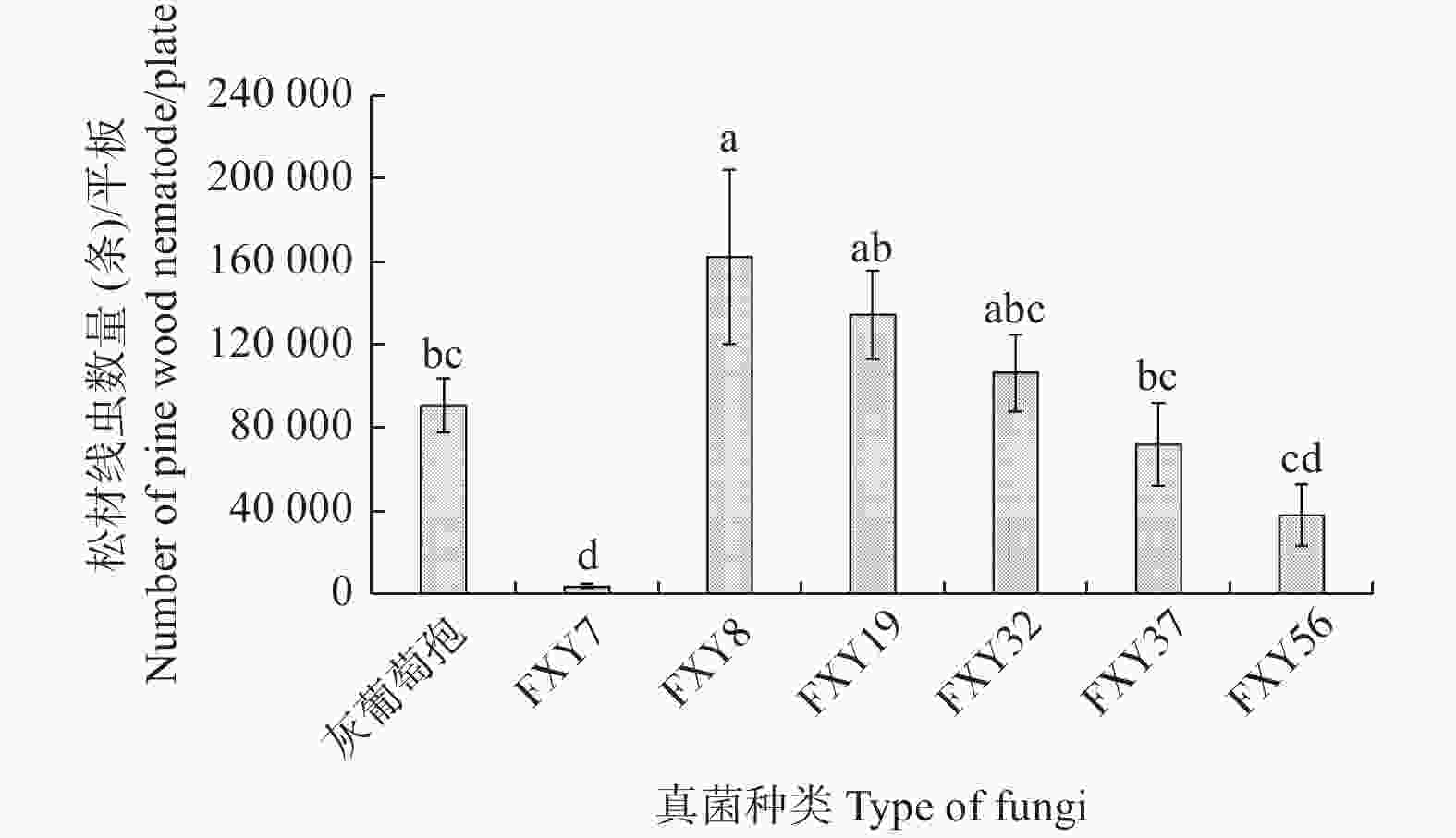
图 3 取食不同真菌后活体松材线虫的种群数量
Figure 3. Populationquantity of live pine wood nematodes fed on different fungi
-
本研究从浙江健康和枯死马尾松中共分离出隶属于22个属的192株真菌和16个属的105株细菌,健康和枯死马尾松真菌菌群之间的相似性较低,不同部位的优势菌群不同,但优势类群均为木霉属真菌(表1和表2),而健康和枯死马尾松细菌菌群之间的相似性较高,优势菌群均为伯克氏菌属、沙雷氏菌和肠杆菌属,而且不同部位的优势类群也较为一致(表3和表4)。从物种多样性来看,健康马尾松的真菌和细菌多样性都高于枯死马尾松,尤其是真菌,这表明由于松材线虫的侵染,降低了马尾松内生真菌的多样性。Zhang等人[19]利用高通量测序技术对浙江健康和枯死马尾松的内生真菌和细菌群落结构进行分析,发现健康马尾松中优势真菌为青霉属、拟盘多毛孢属和木霉属真菌,在枯死马尾松中这三类真菌的丰度有所下降,而优势细菌在健康和枯死马尾松中的分布较为一致。他们还发现枯死马尾松的内生细菌和真菌的丰富度和多样性都低于健康马尾松,尤其是真菌,这与我们的分离结果也是一致的,表明我们的分离结果可以反映浙江健康和枯死马尾松内生真菌和细菌的菌群结构。
曾凡勇[20]对浙江健康、衰弱和枯死马尾松树干的内生真菌进行了分离,木霉均为优势菌,且衰弱木中分离到的真菌数量最多,枯死木中最少。肖育贵等[21]分析了四川不同健康状态马尾松树干木居真菌的种类,分离出的真菌种类主要有木霉属、镰刀菌属和多毛孢属等,健康木分离到的真菌数量最少。鲁国华等[22]对安徽省不同健康状态马尾松树干内的真菌种类进行了研究,发现健康马尾松树干的优势真菌为镰刀菌和木霉,濒死马尾松和死亡马尾松均为长喙壳和木霉,健康马尾松上分离到的真菌数量最少。本研究中对树干的真菌分离结果显示健康木的真菌种类和数量均高于枯死马尾松,这可能与采集枯死马尾松的时间和状态有关,树木的腐烂程度会影响树体内真菌的群落结构[23-24]。邓慧华[25]对福建省健康马尾松不同组织的内生真菌进行了分离,叶部分离到的内生真菌数量和种类最多,茎部次之,根部最少,毛霉属和木霉属是马尾松内生真菌中的优势属。结合本研究和前人的分离结果可知,木霉属是马尾松体内的优势真菌。
松树体内的真菌种群与松材线虫病的发生和发展密切相关。吕全等[26]研究了3个马尾松树栖真菌菌株对媒介昆虫携带松材线虫数量的影响,发现不适宜松材线虫繁殖的真菌能促进松材线虫群体内更多个体转化为扩散型虫态,不同真菌对蛹室中线虫种群大小有显著影响。Maehara和Futai[27]发现在只接种Trichoderma sp.的木段上天牛携带的线虫数量很少,另外在木段上接种Trichoderma sp. 3可以抑制蓝变菌的扩散,从而减少天牛携带的线虫数量[28-29],由此可见,木霉对松材线虫种群的扩散也有一定的抑制作用。本研究中共分离到6种木霉,通过取食实验可知,FXY7显著抑制了松材线虫的种群数量。木霉属真菌是一类高效的拮抗、促生菌,对松材线虫也具有一定的致死作用。钩状木霉Trichoderma hamatum孢子悬浮液对松材线虫毒性较强,致死率可以达到85.6%[30]。木霉菌株T-28的乙酸乙酯提取物在72 h校正死亡率达到100%[31]。木霉的挥发性化合物对松材线虫也具有一定的杀虫活性[32-33]。本研究分离到的木霉对松材线虫病是否具有其它生防作用还需进一步展开研究。
关于马尾松内生细菌群落结构的研究较少,袁文婷对健康马尾松茎部的内生细菌进行了分离,共分离到20株细菌,通过内生性验证后对其中7个菌株进行了种类鉴定,并未对群落结构进行描述。尹诗恒等[34]研究了松材线虫侵染下马尾松苗不同部位内生细菌菌群的结构变化,发现松干部位菌群结构变化极显著,其次是松针,根部基本不受影响。前人也从马尾松体内分离出一些具有生防作用的内生细菌,李亮亮等[35]从马尾松茎部分离到一株松材线虫拮抗细菌短小芽孢杆菌,并通过荧光标记法证明了该菌株在马尾松体内的内生性,为该菌今后的实际应用提供了理论依据。Song等人[36]从马尾松顶芽和针叶中分离到一株Pantoea eucalypti,根据其基因组特征和携带质粒的功能推测该菌可促进松树生长。沙雷氏菌属细菌也是马尾松中经常分离到的一类细菌,从蛹室分离到的Serratia marcescens可以促进松褐天牛的死亡[37]。本研究中分离到多株沙雷氏菌属和芽孢杆菌属细菌,这些菌株对松材线虫病的控制作用还有待进一步研究。
-
健康马尾松树体内的真菌多样性高于枯死马尾松,二者真菌菌群相似性较低,而且不同组织间真菌菌群相似性也较低;对于细菌来说,健康和枯死马尾松的细菌群落具有一定的相似性。这表明松材线虫入侵会影响马尾松树体内的真菌多样性,但对细菌的影响较小。另外本研究中分离到多株可能对松材线虫病具有生防潜力的内生菌,为探索松材线虫病的新型防控技术奠定了基础。
松材线虫侵染前后马尾松树体内微生物多样性分析
Microbial Diversity Analysis of Pinus massoniana Before and After Infected by Pine Wood Nematode
-
摘要:
目的 以浙江省松材线虫病疫区健康和枯死马尾松(Pinus massoniana Lamb.)为研究对象,探究松材线虫(Bursaphelenchus xylophilus)侵染前后马尾松树体内微生物群落结构的差异,同时为利用马尾松内生微生物防治松材线虫病积累微生物种质资源。 方法 采用组织分离法、形态学和分子生物学相结合的方法,对健康和枯死马尾松不同组织内的(根部、干部和针叶)真菌和细菌进行分离鉴定。对分离的木霉菌株构建系统发育树,并分析了松材线虫取食木霉对其种群数量的影响。 结果 从健康和枯死马尾松树体内分别分离得到真菌19个属共106个菌株和10个属共86个菌株,优势类群均为木霉属真菌。不同组织间真菌多样性水平存在差异,干部和针叶的多样性指数高于根部。健康马尾松真菌的Shannon-Wiener指数(H)、Simpson指数(D)和均匀度指数(J)依次为2.37、0.87和0.51,高于枯死马尾松的1.72、0.76和0.39,二者的相似性指数小于0.5。从健康和枯死马尾松树体内分别分离得到细菌13个属共59个菌株和12个属共56个菌株,优势类群均为伯克氏菌属、肠杆菌属和沙雷氏菌属细菌。健康和枯死马尾松中均为干部的细菌多样性最高,针叶的细菌多样性最低,二者的相似性指数大于0.5。木霉作为马尾松树体内的优势类群,本次共分离到6个种,通过取食实验发现木霉FXY7显著抑制了松材线虫的种群数量。 结论 健康马尾松树体内的真菌多样性高于枯死马尾松,二者的真菌菌群相似性较低,而且不同组织间真菌菌群相似性也较低,但健康和枯死马尾松的细菌群落具有一定的相似性。这表明松材线虫入侵会影响马尾松体内的真菌多样性,但对细菌的影响较小。本研究中还分离到多株可能对松材线虫病具有生防潜力的内生菌,为探索松材线虫病的新型防控技术奠定了基础。 Abstract:Objective Taking healthy and dead Pinus massoniana Lamb. in Bursaphelenchus xylophilus infected area in Zhejiang province as research object to examine the differences in the microbial communities structure of P. massoniana trees before and after infected by B. xylophilus, and to accumulate the microbial resources for controlling pine wilt disease by using endophytic microorganisms of P. massoniana. Method The methods of tissue isolation, morphology and molecular biology were used to isolate and identify the fungi and bacteria in different tissues (root, stem and needle) of healthy and dead P. massoniana. Phylogenetic tree was constructed for the isolated Trichoderma strains, and the effect of Trichoderma spp. on B. xylophilus population was analyzed by feeding experiments. Result 106 fungal strains belonging to 19 genera were isolated from healthy P. massoniana, and 86 strains belonging to 10 genera were isolated from the dead pine, the dominant genera were both Trichoderma. There were differences in fungal diversity among different tissues, the diversity indexes of stem and needle were higher than those of root. The Shannon-Wiener index, Simpson index and evenness index of fungal community from healthy pine were 2.37, 0.87 and 0.51, respectively, they were all higher than those of dead pine, which were 1.72, 0.76 and 0.39. The similarity indexes of fungal community in healthy and dead P. massoniana were less than 0.5. 59 bacterial strains belonging to 13 genera were isolated from healthy P. massoniana, and 56 strains belonging to 12 genera were isolated from the dead pine, the dominant groups were Burkholderia, Enterobacter and Serratia. The bacterial diversity of stem was the highest in both the healthy and dead pine, and that of needle was the lowest. The similarity indexes of bacterial community in healthy and dead P. massoniana were more than 0.5. Trichoderma was the dominant group in P. massoniana, six species were isolated in this study. The feeding experiment showed that the sample FXY7 of Trichoderma significantly inhibited the population of B. xylophilus. Conclusion The fungal diversity in healthy P. massoniana is higher than that in dead P. massoniana, the similarity of fungal community between healthy and dead pine is low, and the similarity of fungal community among different tissues is also low. However, the bacterial communities of healthy and dead pine are similar to some extents. This indicates that the invasion of B. xylophilus will affect the fungal community diversity in P. massoniana, but there is a little effect on the bacterial community. In addition, a number of endophytic bacteria with bio-control potential against pine wilt disease were isolated in this study, which is helpful for exploring prevention and control technology of pine wilt disease. -
Key words:
- Bursaphelenchus xylophilus
- / Pinus massoniana
- / fungi
- / bacteria
- / diversity
-
图 2 基于ITS序列构建的木霉系统发育进化树
Figure 2. Phylogenetic tree of Trichoderma strains inferred from ITS gene sequences
图 3 取食不同真菌后活体松材线虫的种群数量
Figure 3. Populationquantity of live pine wood nematodes fed on different fungi
表 1 健康马尾松不同部位内生真菌的组成及分离频率
Table 1. The fungal composition and isolation frequency of different tissues in healthy Masson pine
编号
No.属 Genus 根 Root 干 Stem 叶 Needle 菌株总数
Total number of
strains相对频率
Relative
frequency/%菌株数量
Number of
strains分离频率
Frequency/
%菌株数量
Number of
strains分离频率
Frequency/
%菌株数量
Number of
strains分离频率
Frequency/
%1 木霉属
Trichoderma0 0 23 21.70 1 0.94 24 22.64 2 链格孢属
Alternaria0 0 2 1.89 5 4.72 7 6.60 3 小枝孢属
Brachysporiella0 0 2 1.89 0 0 2 1.89 4 青霉属
Penicillium1 0.94 7 6.60 0 0 8 7.55 5 散斑壳属
Lophodermium0 0 0 0 7 6.60 7 6.60 6 拟盘多毛孢属
Pestalotiopsis0 0 6 5.66 3 2.83 9 8.49 7 短柄霉属
Aureobasidium0 0 1 0.94 0 0 1 0.94 8 炭疽菌属
Colletotrichum0 0 0 0 8 7.55 8 7.55 9 拟盾壳霉属
Paraconiothyrium0 0 0 0 19 17.92 19 17.92 10 毛色二孢属
Lasiodiolodia4 3.78 0 0 0 0 4 3.77 11 被孢霉属
Mortierella0 0 3 2.83 0 0 3 2.83 12 伞状霉属
Umbelopsis0 0 5 4.72 0 0 5 4.72 13 炭团菌属
Hypoxylon0 0 1 0.94 0 0 1 0.94 14 亚罗酵母属
Yarrowia0 0 1 0.94 0 0 1 0.94 15 皮司霉属
Pithomyces0 0 2 1.89 0 0 2 1.89 16 黑孢属
Nigrospora0 0 0 0 2 1.89 2 1.89 17
Kirschsteiniothelia0 0 1 0.94 0 0 1 0.94 18 Eurotiomycetes 0 0 1 0.94 0 0 1 0.94 19
Atractidochium0 0 0 0 1 0.94 1 0.94 合计 Total 5 4.72 55 51.88 46 43.40 106 1 香农-威纳指数/H Shannon-Wiener index/H 0.50 1.81 1.68 2.37 辛普森指数/D Simpson index/D 0.32 0.78 0.76 0.87 均匀度指数/J Evenness index/J 0.31 0.45 0.44 0.51  下载: 导出CSV
下载: 导出CSV
表 2 枯死马尾松不同部位内生真菌的组成及分离频率
Table 2. The fungal composition and isolation frequency of different tissues in dead Masson pine
编号
No.属
Genus根 Root 干 Stem 叶 Needle 菌株总数
Total number
of strains相对频率
Relative
frequency/%菌株数量
Number of
strains分离频率
Frequency/
%菌株数量
Number of
strains分离频率
Frequency/
%菌株数量
Number of
strains分离频率
Frequency/
%1 木霉属
Trichoderma8 9.30 22 25.58 0 0 30 34.88 2 链格孢属
Alternaria0 0 1 1.16 1 1.16 2 2.33 3 半帚孢属
Leptographium0 0 4 4.65 0 0 4 4.65 4 小枝孢属
Brachysporiella0 0 1 1.16 0 0 1 1.16 5 青霉属
Penicillium2 2.33 0 0 0 0 2 2.33 6 长喙壳属
Ophiostoma0 0 4 4.65 0 0 4 4.65 7 散斑壳属
Lophodermium0 0 0 0 27 31.40 27 31.40 8 拟盘多毛孢属
Pestalotiopsis0 0 0 0 10 11.63 10 11.63 9 微色二孢属
Microdiplodia0 0 0 0 3 3.49 3 3.49 10 短柄霉属
Aureobasidium0 0 0 0 3 3.49 3 3.49 合计 Total 10 11.63 32 37.21 44 51.16 86 1 香农-威纳指数/H Shannon-Wiener index/H 0.50 0.99 1.09 1.73 辛普森指数/D Simpson index/D 0.32 0.49 0.56 0.76 均匀度指数/J Evenness index/J 0.22 0.29 0.29 0.39
下载: 导出CSV
表 3 健康马尾松不同部位内生细菌的组成及分离频率
Table 3. The bacterial composition and isolation frequency of different tissues in healthy Masson pine
编号
No.属 Genus 根 Root 干 Stem 叶
Needle菌株总数
Total number
of strains相对频率
Relative
frequency/%菌株数量
Number of
strains分离频率
Frequency/
%菌株数量
Number of
strains分离频率
Frequency/
%菌株数量
Number of
strains分离频率
Frequency/
%1 伯克氏菌属 Burkholderia 15 25.42 3 5.08 0 0 18 30.51 2 沙雷氏菌属 Serratia 1 1.69 7 11.86 2 3.39 10 16.95 3 肠杆菌属 Enterobacter 0 0 9 15.25 1 1.69 10 16.95 4 拉乌尔菌属 Raoultella 1 1.69 2 3.39 0 0 3 5.08 5 泛菌属 Pantoea 0 0 4 6.78 0 0 4 6.78 6 欧文氏菌属 Erwinia 0 0 1 1.69 0 0 1 1.69 7 杆菌属 Bacterium 0 0 2 3.39 1 1.69 3 5.08 8 假单胞菌属 Pseudomonas 3 5.08 0 0 0 0 3 5.08 9 鞘氨醇单胞菌属 Sphingomonas 1 1.69 1 1.69 0 0 2 3.39 10 克吕沃尔氏菌属 Kluyvera 0 0 1 1.69 0 0 1 1.69 11 类芽孢杆菌属 Paenibacillus 0 0 1 1.69 0 0 1 1.69 12 Paraburkholderia 0 0 1 1.69 0 0 1 1.69 13 Uncultured bacterium 0 0 2 3.39 0 0 2 3.39 合计 Total 21 35.59 34 57.63 4 6.78 59 1
下载: 导出CSV
表 4 枯死马尾松不同部位内生细菌的组成及分离频率
Table 4. The bacterial composition and isolation frequency of different tissues in dead Masson pine
编号
No.属
Genus根 Root 干 Stem 叶 Needle 菌株总数
Total number
of strains相对频率
Relative
frequency/%菌株数量
Number of
strains分离频率
Frequency/
%菌株数量
Number of
strains分离频率
Frequency/
%菌株数量
Number of strains分离频率
Frequency/
%1 伯克氏菌属 Burkholderia 19 33.93 3 5.36 0 0 22 39.29 2 沙雷氏菌属 Serratia 2 3.57 4 7.14 0 0 6 10.71 3 芽孢杆菌属 Bacillus 3 5.36 0 0 2 3.57 5 8.93 4 肠杆菌属 Enterobacter 0 0 8 14.29 0 0 8 14.29 5 果胶杆菌属 Pectobacterium 0 0 3 5.36 0 0 3 5.36 6 拉乌尔菌属 Raoultella 0 0 1 1.79 0 0 1 1.79 7 泛菌属 Pantoea 0 0 3 5.36 0 0 3 5.36 8 欧文氏菌属 Erwinia 0 0 1 1.79 0 0 1 1.79 9 克雷伯氏菌属 Klebsiella 0 0 1 1.79 0 0 1 1.79 10 杆菌属 Bacterium 0 0 2 3.57 2 3.57 4 7.14 11 Paraburkholderia 0 0 1 1.79 0 0 1 1.79 12 Uncultured bacterium 1 1.79 0 0 0 0 1 1.79 合计 Total 25 44.64 27 48.21 4 7.14 56 1
下载: 导出CSV
表 5 健康和枯死马尾松不同部位真菌菌群相似性系数
Table 5. The similarity indexes of fungal community of different tissue from healthy and dead Masson pine
健康马尾松
Healthy Masson pine枯死马尾松
Dead Masson pine根 Root 干S tem 针叶 Needle 根 Root 干 Stem 针叶 Needle 根 Root 0.07 0.00 0.17 0.00 干 Stem 0.13 0.17 0.29 0.11 针叶 Needle 0.00 0.29 0.00 0.2 注:表中对角线以上为Jaccard指数,以下为Sorenson指数。
The values above the diagonal in the table were Jaccard index, and the values below were Sorenson index.
下载: 导出CSV
-
[1] Rosier A, Bishnoi U, Lakshmanan V, et al. A perspective on inter-kingdom signaling in plant-beneficial microbe interactions[J]. Plant Molecular Biology, 2016, 90(6): 537-548. doi: 10.1007/s11103-016-0433-3 [2] Sasse J, Martinoia E, Northen T. Feed your friends: Do plant exudates shape the root microbiome?[J]. Trends in Plant Science, 2017, 23(1): 25-41. [3] Soliman S, Greenwood J, Bombarely A, et al. An endophyte constructs fungicide-containing extracellular barriers for its host plant[J]. Current Biology, 2015, 25(19): 2570-2576. doi: 10.1016/j.cub.2015.08.027 [4] Terhonen E, Sipari N, Asiegbu F O. Inhibition of phytopathogens by fungal root endophytes of Norway spruce[J]. Biological Control, 2016, 99: 53-63. doi: 10.1016/j.biocontrol.2016.04.006 [5] Hiruma K, Gerlach N, Sacristán S, et al. Root endophyte Colletotrichum tofieldiae confers plant fitness benefits that are phosphate status dependent[J]. Cell, 2016, 165(2): 464-474. doi: 10.1016/j.cell.2016.02.028 [6] Khan A L, Hussain J, Al-Harrasi A, et al. Endophytic fungi: resource for gibberellins and crop abiotic stress resistance[J]. Critical Reviews in Biotechnology, 2013, 35(1): 62-74. [7] Wu F L, Li Y, Tian W, et al. A novel dark septate fungal endophyte positively affected blueberry growth and changed the expression of plant genes involved in phytohormone and flavonoid biosynthesis[J]. Tree Physiology, 2020, 40(8): 1080-1094. doi: 10.1093/treephys/tpaa047 [8] Carro-Huerga G, Compant S, Gorfer M, et al. Colonization of Vitis vinifera L. by the endophyte Trichoderma sp. strain T154: biocontrol activity against Phaeoacremonium minimum[J]. Frontiers in Plant Science, 2020, 11: 1170. doi: 10.3389/fpls.2020.01170 [9] 理永霞, 张星耀. 我国中温带面临的松材线虫入侵扩张高风险[J]. 温带林业研究, 2018, 1(1):3-6. doi: 10.3969/j.issn.2096-4900.2018.01.002 [10] Wingfield M J. Transmission of pinewood nematode to cut timber and girdled trees[J]. Plant Disease, 1983, 67(1): 35-37. doi: 10.1094/PD-67-35 [11] Alves M, Pereira A, Vicente C, et al. The role of bacteria in pine wilt disease: insights from microbiome analysis[J]. FEMS Microbiology Ecology, 2018, 94(7): 1-13. [12] 曾凡勇, 骆有庆, 吕 全, 等. 松材线虫入侵的黑松内栖真菌区系初步研究[J]. 林业科学研究, 2006, 19(4):537-540. doi: 10.3321/j.issn:1001-1498.2006.04.025 [13] 鲁国华. 马尾松、黑松树干内真菌种类及其变化[D]. 江苏: 南京林业大学, 2008. [14] 徐风美. 松萎蔫病发生区和未发生区油松根部真菌研究[D]. 杨凌: 西北农林科技大学, 2014. [15] Ma Y, Qu Z L, Liu B, et al. Bacterial community structure of Pinus thunbergii naturally infected by the nematode Bursaphelenchus xylophilus[J]. Microorganisms, 2020, 8: 307. doi: 10.3390/microorganisms8020307 [16] Ponpandian L N, Rim S O, Shanmugam G, et al. Phylogenetic characterization of bacterial endophytes from four Pinus species and their nematicidal activity against the pine wood nematode[J]. Scientific Reports, 2019, 9: 12457. doi: 10.1038/s41598-019-48745-6 [17] Kim N, Jeon H W, Mannaa M, et al. Induction of resistance against pine wilt disease caused by Bursaphelenchus xylophilus using selected pine endophytic bacteria[J]. Plant Pathology, 2019, 68: 434-444. doi: 10.1111/ppa.12960 [18] 刘济明, 柳嘉佳, 颜 强, 等. 小蓬竹植物内生真菌多样性[J]. 东北林业大学学报, 2016, 44(12):71-75. doi: 10.3969/j.issn.1000-5382.2016.12.015 [19] Zhang W, Wang X, Li Y X, et al. Pinewood nematode alters the endophytic and rhizospheric microbial communities of Pinus massoniana[J]. Microbial Ecology, 2021, 81: 807-817. doi: 10.1007/s00248-020-01619-1 [20] 曾凡勇. 松树内栖真菌对松材线虫种群动态的影响[D]. 北京: 北京林业大学, 2006. [21] 肖育贵, 周建华, 肖银波. 四川马尾松木居真菌数量与松材线虫病发生的可能性分析[J]. 四川林业科技, 2009, 30(2):30-32. doi: 10.3969/j.issn.1003-5508.2009.02.007 [22] 鲁国华, 叶建仁. 安徽省马尾松树干内真菌种类研究[J]. 南京林业大学学报:自然科学版, 2011, 35(1):132-134. [23] Høiland K, Bendiksen E. Biodiversity of wood-inhabiting fungi in a boreal coniferous forest in Ser-Trendelag County, Central Norway[J]. Nordic Journal of Botany, 1997, 16(6): 643-659. [24] Ódor P, Heilmann-Clausen J, Christensen M, et al. Diversity and composition of dead wood inhabiting fungal and bryophyte communities in semi-natural beech forests in Europe[J]. Biological Conservation, 2006, 131(1): 58-71. doi: 10.1016/j.biocon.2006.02.004 [25] 邓慧华. 马尾松内生真菌的分离鉴定及多样性[J]. 东北林业大学学报, 2015, 43(7):140-142. doi: 10.3969/j.issn.1000-5382.2015.07.031 [26] 吕 全, 刘慧敏, 曾凡勇, 等. 树栖真菌对扩散型松材线虫生活史的影响[J]. 植物病理学报, 2015, 45(2):121-129. [27] Maehara N, Futai K. Effect of fungal interactions on the numbers of the pinewood nematode, Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae), carried by the Japanese pine sawyer, Monochamus alternatus (Coleoptera: Cerambycidae)[J]. Fundamental and Applied Nematology, 1997, 20(6): 611-617. [28] Maehara N, Hata K, Futai K. Effect of blue-stain fungi on the number of Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae) carried by Monochamus alternatus (Coleoptera: Cerambycidae)[J]. Nematology, 2005, 7(2): 161-167. doi: 10.1163/1568541054879557 [29] Maehara N, Tsuda K. , Yamasaki M, et al. Effect of fungus inoculation on the number of Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae) carried by Monochamus alternatus (Coleoptera: Cerambycidae)[J]. Nematology, 2006, 8(1): 59-67. doi: 10.1163/156854106776179944 [30] 田忠玲. 大豆孢囊线虫生防真菌的筛选、鉴定及应用基础研究[D]. 浙江: 浙江大学, 2016. [31] 魏博涵, 张 锴, 申加兴, 等. 15个生防菌株乙酸乙酯提取物对松材线虫的杀虫活性[J]. 林业科技, 2017, 42(1):24-26. doi: 10.3969/j.issn.1001-9499.2017.01.008 [32] Yang Z S, Yu Z F, Lei L P, et al. Nematicidal effect of volatiles produced by Trichoderma sp.[J]. Journal of Asia-Pacific Entomology, 2012, 15(4): 647-650. doi: 10.1016/j.aspen.2012.08.002 [33] 袁文婷. 松树内生细菌种群初探及生防益菌的室内筛选[D]. 南京: 南京林业大学, 2010. [34] 尹诗恒, 张绍勇, 刘骕骦, 等. 松材线虫侵染对马尾松苗不同部位内生细菌菌群结构的影响[J]. 浙江农林大学学报, 2021, 38(4):846-853. doi: 10.11833/j.issn.2095-0756.20200562 [35] 李亮亮, 谈家金, 陈凤毛. GFP标记短小芽孢杆菌LYMC-3在马尾松体内的定殖[J]. 华中农业大学学报, 2016, 35(6):68-73. [36] Song Z W, Lu Y M, Liu X W, et al. Evaluation of Pantoea eucalypti FBS135 for pine (Pinus massoniana) growth promotion and its genome analysis[J]. Journal of Applied Microbiology, 2020, 129(4): 958-970. doi: 10.1111/jam.14673 [37] Zhang C, Wickham J, Zhao L L, et al. A new bacteria-free strategy induced by MaGal2 facilitates pinewood nematode escape immune response from its vector beetle[J]. Insect Science, 2020. doi: 10.1111/1744-7917.12823. -

 点击查看大图
点击查看大图

计量
- 文章访问数: 3921
- HTML全文浏览量: 2645
- PDF下载量: 109
- 被引次数: 0





