



-
由于邻近植物个体对有限资源的共同需求,群落中植物种间存在相互竞争,从而使竞争双方或某一方植物的生长和发育受到阻碍[1],而植物间的竞争作用会对森林群落物种的天然更新、分布格局、多样性、结构等方面产生影响[2-3],进而影响群落的生产力和稳定性。生态位能有效反映物种在其所处的生态系统中的地位关系,可以将种间关系定量化描述和研究,其理论成为解释群落中物种共存与竞争机制的基本理论[4-6],而物种多样性和群落结构可以由物种生态位宽度和种间生态位重叠决定,能定量反映物种对资源的利用能力、在群落或生态系统中的功能位置及其所在群落的稳定性[7-9]。所以,基于生态位理论研究群落内物种间的相互关系,对深入揭示群落物种分布、共存及群落稳定性机制至关重要。
油松(Pinus tabuliformis Carr.)为我国特有树种,是华北植物区系的代表种之一,其根系发达、枝叶繁茂、具有较强的抗旱性和天然更新能力[10-11],对区域水土保持起到非常重要的作用[12]。本研究通过对晋南油松人工林林下植被的调查发现,油松天然更新存在障碍。牛健[13]通过综合前人研究发现,球果和种子质量、枯落物、坡向、坡位、郁闭度、干扰等是导致油松天然更新障碍的主要原因;但近年来,中条山林业主管部门对其辖区内不同类型油松人工林采取了人工促进天然更新(图1),如清除枯枝落叶、间伐等措施,发现当年春季油松萌发数量较多,但第2年存活较少。所以,种实、枯落物、非生物环境因素并不是影响该地区油松更新障碍的主要原因,基于此,本研究提出以下假设:物种之间的竞争可能是影响该地区油松天然更新的主要原因。为了证明该假设,本研究基于中条山油松人工林群落样地调查数据,通过分析油松人工林下主要植物的种间关联性、生态位重叠及群落稳定性,探讨种间关联和生态位理论在油松人工林天然更新障碍及群落稳定性形成机制中的作用,为油松人工林经营管理提供理论依据。

图 1 人工促进油松天然更新
Figure 1. Artificial promotion of natural regeneration of P. tabuliformis
-
本研究位于山西省中条山十河林场(35°31′~35°51′ N,111°50′~112°00′ E),属太行山向南延伸的余脉,处于黄土高原向华北平原过渡的东南边缘区,土壤类型随海拔由低到高分别为山地褐土、淋溶褐土、棕壤、亚高山草甸土。该区属于暖温带半湿润大陆季风气候,年均气温10.3~14.3℃,≥10℃的年积温为3 400~4 700℃,年均降水量为524.3~654.3 mm,无霜期199~229 d[14-16]。植被属暖温带落叶阔叶林带,主要树种有油松、华山松(P. armandii Franch.)、辽东栎(Quercus wutaishanica Mary)、橿子栎(Q. baronii Skan)、侧柏(Platycladus orientalis Linn.)、鹅耳枥(Carpinus turczaninowii Hance)、栓皮栎(Q. variabilis Bl.)和白桦(Betula platyphylla Suk.)等,林下植被主要由辽东栎、华山松、油松、连翘(Forsythia suspensa Thunb.)、土庄绣线菊(Spiraea pubescens Turcz.)、披针薹草(Carex lancifolia C. B. Clarke)、早开堇菜(Viola prionantha Bunge)、茜草(Rubia cordifolia L.)和金线草(R. membranace Thunb.)等物种组成。
-
在对中条山油松人工林全面踏查的基础上,在油松人工林分布的核心地区进行样地调查,样地概况见表1。样地面积为675 m2或1 200 m2,利用网格法将每个样地分割成3个20 m×20 m或15 m×15 m的样方,对样方内胸径(DBH)≥5 cm的木本植物进行每木检尺,记录每个个体的物种名、胸径、树高等,根据二类调查数据查询林分年龄。在每个20 m×20 m(15 m×15 m)样方内设置16(9)个5 m×5 m和1 m×1 m的灌木和草本样方,记录物种名、多度、株高、盖度等。共计录207种植物,其中,乔木29种、灌木47种、草本112种、藤本19种;同时记录每个样地的经纬度、海拔、郁闭度、坡度、坡向等环境因子。
表 1 油松人工林群落样地概况
Table 1. The information of community plots of P. tabuliformis Carr. plantation
序号
No.经度(E)
Longitude纬度(N)
Latitude树种组成
Species composition林龄
Age/a海拔
Elevation/
m样地面积
Area/
m2坡向
Slope aspect坡度
Slope/
(°)郁闭度
Coverage/
%平均胸径
Dq/
cm平均树高
Height/
m密度
Density/
(株∙ hm−2)1 111.92° 35.50° 6 油+4 华 40 1 916 1 200 西南 SW 22 80 24.08 14.89 808 2 111.92° 35.50° 6 油+4 辽 46 1 924 675 东南 SE 34 75 17.56 9.31 1 511 3 111.88° 35.50° 10 油 28 1 880 1 200 西 W 19 60 14.77 12.43 1 600 4 111.94° 35.51° 8 油+2 辽 38 1 750 1 200 西南 SW 36 55 18.66 11.20 925 5 111.93° 35.51° 8 油+2 华 48 1 722 1 200 东 E 21 85 25.64 15.46 508 6 111.90° 35.49° 9 油+1 华 48 1 933 1 200 东北 NE 13 80 27.84 16.31 583 7 111.90° 35.49° 10 油 41 1 917 1 200 西南 SW 20 60 27.79 14.74 500 8 111.94° 35.48° 8 油+2 辽 36 1 509 1 200 南 S 26 80 17.39 11.60 1 383 9 111.92° 35.51° 8 油+1 辽+1 华 39 1 997 1 200 西南 SW 25 75 14.79 9.07 2 475 10 111.89° 35.49° 9 油+1 落 32 1 872 1 200 东南 SE 11 70 17.39 12.95 1 250 11 111.90° 35.49° 10 油 48 1 928 1 200 西 W 8 60 29.32 15.94 467 12 111.90° 35.50° 7 油+2 华+1 辽 46 1 960 1 200 西南 SW 23 75 21.76 10.90 558 -
基于样地调查数据,选取油松林下植物物种为研究对象,建立“样方—物种多度”矩阵,共计36个样方数据用于种间关联分析。采用方差比率(VR)检验群落物种总体联结性,若VR>1则表示群落物种总体正关联,VR<1则表示负关联。采用W统计量来检验方差比率的显著程度,若W落入95%的置信区间内,则群落物种总体关联不显著,反之则表示显著关联[7]。采用经过Yates[17]矫正的χ2检验物种之间是否存在显著关联;基于联结系数(AC)得到物种之间联结的紧密程度,若AC值越趋向于1.00,表示种间正联结越紧密,AC值越趋向于−1.00,表示种间负联结越紧密,AC=0则表示种间相对独立;采用Pearson相关系数判断种对之间的相关程度[8];采用Levins指数度量物种的生态位宽度(Bi),同时计算种对之间的Levins生态位重叠指数(Oik)[9],并绘制物种生态位重叠指数半矩阵图。由于物种数较多,群落中常见种种间关系更容易监测到,且常见种决定群落物种分布[18-19],所以,选择群落中频度最高的30个常见物种进行分析。
-
采用Godron稳定性测定方法评价群落的稳定性。以36个样方为单位,将群落中不同物种的频度由大到小排序,将物种的频度换算成相对频度,按相对频度由大到小逐步累积相加,同时,一一对应逐步累积总物种数倒数,采用散点平滑曲线模拟两次方程和两个坐标轴100处连接的对角直线来判断累积总物种数倒数与累积相对频度比值的交点坐标,20/80是群落的稳定点,该交点坐标越接近20/80,群落就越稳定[20]。
-
数据整理采用Excel 2016(Microsoft,USA)完成,分析及作图采用R 4.0.3软件中“vegan”、“spaa”程序包完成。
-
中条山油松人工林下最常见的30个物种的总体联结性分析结果表明,30个物种的方差比率为1.98,大于1,表明种间存在正联结,其W统计量为71.22,高于χ2分布临界值(17.71),表明物种间总体上显著正联结。
表2表明:30个物种共形成406个种对,其中,辽东栎因频度太大在每个样方中都有分布而无法分析其与共处群落中其他种之间的关联性,即偏离种。χ2检验结果显示:无联结种对共计372对,占全部种对的91.63%;有联结种对共计34对,占全部种对的8.37%,其中,极显著和显著联结的种对分别为15对和19对,分别占有联结种对的44.12%和55.88%,说明常见种间联结较松散。
表 2 常见物种间关联分析结果
Table 2. Results of interspecific association analysis of common species
种间关联 Interspecific association 类型 Types 数值范围 Value range 种对数 Species pair number 百分比/% χ2 检验
Chi−square test无联结 No association p≥0.05 372 91.63 有联结 Association p<0.01 15 3.69 0.01≤p<0.05 19 4.68 总数 Total 406 100.00 联结系数
Association coefficient正联结 Positive association AC=1 26 6.40 0.60≤AC<1.00 23 5.67 0.20≤AC<0.60 96 23.65 0<AC<0.20 66 16.26 无联结 No association AC=0 1 0.25 负联结 Negative association −0.20<AC<0 26 6.40 −0.60<AC≤−0.20 73 17.98 −1.00<AC≤−0.60 23 5.67 AC=−1 72 17.73 总数 Total 406 100.00 Pearson相关
Pearson correlation正相关 Positive correlation p<0.01 50 11.49 0.01≤p<0.05 25 5.75 p≥0.05 139 31.95 负相关 Negative correlation p<0.01 1 0.23 0.01≤p<0.05 10 2.30 p≥0.05 210 48.28 总数 Total 435 100.00 联结系数分析结果(表2)显示:406个种对中(存在1个偏离种),正联结211个种对,负联结194个种对,无联结1个种对,分别占全部种对的51.97%、47.78%、0.25%。正联结显著的种对(AC≥0.20,145对)小于负联结显著的种对(AC≤−0.20,168对),其中,极显著正联结(AC≥0.60)种对共计49对,显著正联结(0.20≤AC<0.60)种对共计96对,极显著负联结(AC≤−0.60)种对共计95对,显著负联结(−0.60<AC≤−0.20)种对73对。此外,AC=1.00和AC=−1.00的种对分别为26对和72对,其中,油松天然更新竞争最激烈的物种主要为土庄绣线菊、五角枫、粗齿铁线莲、青榨槭、金线草、披碱草,均为AC=−1.00。
Pearson相关分析结果(表2)显示:435个种对中显著相关(p<0.05)的种对为86对,无显著相关(p>0.05)的种对为349对,分别占全部种对的19.77%和80.23%,其中,极显著(p<0.01)和显著(0.01≤p<0.05)正相关的种对分别为50对和25对,极显著和显著负相关的种对分别为1对和10对。常见种整体正相关与负相关比值为0.97,其中,显著(p<0.05)正相关与显著负相关比值为6.82。
-
基于Levins指数计算常见种生态位宽度,结果(表3)表明:中条山油松人工林下常见种披针薹草的生态位最宽(23.47),辽东栎次之(20.60),而红椋子的生态位最窄(3.87),三裂绣线菊次之(4.86),且披针薹草的生态位宽度是红椋子的6.06倍,灌木、草本、藤本、乔木更新的平均生态位宽度分别为10.13、10.25、11.97、12.79,油松天然更新的生态位宽度为14.69,相对较高。
表 3 基于Levins指数计算的常见种生态位宽度
Table 3. Calculating the niche width of common species based on Levins index
物种
Species生态位宽度
Niche width物种
Species生态位宽度
Niche width物种
Species生态位宽度
Niche width披针薹草 Carex lancifolia 23.47 青榨槭 Acer davidii 12.89 高山露珠草 Circaea alpina 8.09 辽东栎 Quercus wutaishansea 20.60 茜草 Rubia cordifolia 11.84 费菜 Sedum aizoon 7.99 华山松 Pinus armandii 18.79 五角枫 Acer pictum subsp. mono 11.69 南蛇藤 Celastrus orbiculatus 7.98 金线草 Rubia membranacea 17.15 土庄绣线菊 Spiraea pubescens 11.55 赤瓟 Thladiantha dubia 7.85 粗齿铁线莲 Clematis argentilucida 15.03 香薷 Elsholtzia ciliata 10.83 青麸杨 Rhus potaninii 7.07 油松 Pinus tabuliformis 14.69 早开堇菜 Viola prionantha 10.72 筋骨草 Ajuga ciliata 5.52 木梨 Pyrus xerophila 14.04 接骨木 Sambucus williamsii 10.41 鹅耳枥 Carpinus turczaninowii 5.31 太平花 Philadelphus pekinensis 14.02 毛丁香 Syringa tomentella 10.06 披碱草 Elymus dahuricus 5.18 榆叶梅 Amygdalus triloba 13.87 灰栒子 Cotoneaster acutifolius 9.82 三裂绣线菊 Spiraea trilobata 4.86 鞘柄菝葜 Smilax stans 13.08 唐古特忍冬 Lonicera tangutica 9.73 红椋子 Swida hemsleyi 3.87 注:表中乔木均为油松人工林下天然更新。
Notes: The trees in the table are natural regeneration under the P. tabuliformis Carr. plantation. -
基于Levins生态位重叠指数分析了30个常见种种对间的生态位重叠情况,并绘制半矩阵图(图2)。由图2和表4可知:生态位重叠指数Oik<0.59的种对有338对(77.70%),而Oik≥0.59的种对有97对(22.30%),30个常见种总体生态位重叠较低,但有11个种对生态位重叠指数大于0.98,如筋骨草与辽东栎、披针薹草、青榨槭、木梨、五角枫等物种的生态位重叠均较大,说明部分种之间的生态相似性较大。油松天然更新与费菜、披针薹草、华山松、赤瓟等物种的生态位重叠较大,说明油松更新与以上几种植物资源利用相似或占据了相近的生态空间。

图 2 Levins生态位重叠半矩阵图
Figure 2. Semi-matrix map of Levins niche overlap
表 4 Levins生态位重叠统计
Table 4. Statistics of Levins niche overlap
项目 Item Oik≥0.98 0.78≤Oik<0.98 0.59≤Oik<0.78 0.40≤Oik<0.59 0.21≤Oik<0.40 Oik≤0.21 种对数 Species pair number 11 36 50 87 115 136 百分比/% 2.53 8.28 11.49 20.00 26.44 31.26 -
将油松人工林内所有物种(207)的累积相对频度与所有物种数的累积倒数一一对应,绘制散点图(图3)。根据种总数倒数累计-相对频度累计的散点图,完成平滑曲线的模拟,其Godron稳定性曲线为y=0.009 6x2−0.038 3x+5.006 9(R2=0.993 7),求得交点坐标(61.28,38.72),距离群落稳定点(20,80)相对较远。
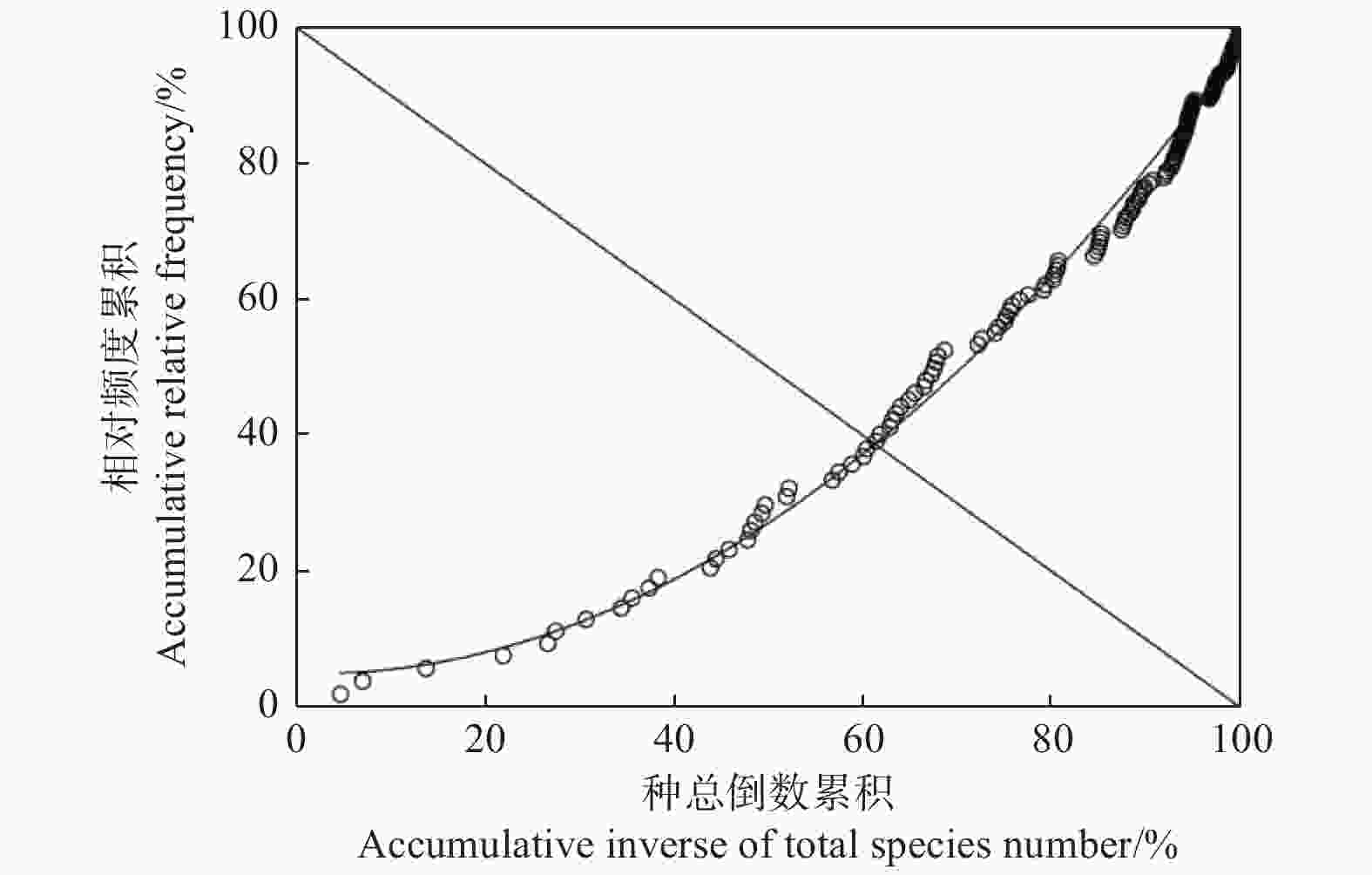
图 3 油松人工林群落Godron稳定性
Figure 3. Godron community stability in the community of P. tabuliformis plantation
-
常见种间总体关联性分析结果表明,30个常见种间存在显著的正联结,与Gu等[21]对五台山油松天然次生林研究结果相似,而简敏菲等[8]认为,若群落物种总体呈正联结,则群落结构及其种类组成将逐渐趋于完善和稳定,种间关系也将逐步趋于正相关。本研究χ2检验结果显示:大部分常见种间的联结关系为无联结,即种间联结非常松散,物种间的相对独立性较强。有研究表明,形成种间关联的过程依赖于环境和植物的生命阶段[22-23]。所以,中条山油松人工林群落常见种联结松散可能与其当前的演替阶段及现存物种本身的生物学特性有关。
物种间的联结性主要受生存环境的异(负联结)同(正联结)、偏利共生或互利共生造成的正联结、竞争相同资源时相互排斥造成的负联结及物种间物理或化感作用导致的正联结或负联结影响[24-25],且理论上认为群落的正负关联比越高,群落结构越趋于稳定,多物种可以稳定共存,群落达到与环境条件相适应的稳定阶段[7]。本研究基于联结系数分析结果发现,油松人工林下植被显著负联结种对数高于正联结,且AC=−1.00的种对有72对,表明油松人工林下部分物种间竞争较激烈,但种间Pearson相关分析结果显示,油松人工林下显著正相关物种高于负相关,主要原因可能是种对Pearson相关系数是反映2个物种间线性关系[26],其处理的是定量数据,反映2个物种同时出现在同一样方的可能性程度。Pearson相关系数分析结果反映油松人工林下30个常见种对间大多数呈正相关,说明常见种对生境具有相同的生态适应性和相互重叠的生态位,这与VR检验结果相一致,而这种较强的正相关性,说明群落的成熟度相对较低,存在一些以竞争相同资源的物种,群落的总体资源利用率和稳定性较低,这与本文的Gordon群落稳定性分析结果一致(图3)。
联结系数将物种之间联结的紧密程度定量化反映[8],可以用来确定与目标植物联系竞争最激烈的物种,而相邻植物间对光照和养分的竞争将直接影响群落中物种的生长[27]。本研究发现,中条山油松人工林下油松天然更新密度为150株·hm−2,总体偏少,对维持油松人工林的稳定性不利;而种间关联分析结果显示,与油松天然更新竞争最激烈的物种主要为土庄绣线菊(AC=−1.00)、五角枫(AC=−1.00)、粗齿铁线莲(AC=−1.00)、青榨槭(AC=−1.00)、金线草(AC=−1.00)、披碱草(AC=−1.00),均为生长速度相对较快的阔叶乔木、灌木、藤本或多年生丛生草本,遮光导致喜光植物油松生长受限,此外五角枫、青榨槭与油松同为深根性植物,在根部也会产生养分竞争。这可能是导致该区域油松天然更新障碍的主要原因之一,所以,今后在以油松为目标树种的森林经营管理中,应加重清除林下与油松天然更新竞争较强的物种。而与油松正关联的植物主要为贴地草本或生长较慢的松科乔木植物,其与油松天然更新间的竞争较小,如披针薹草(AC=1.00)、华山松(AC=1.00)、茜草(AC=0.67)和早开堇菜(AC=0.53)等。
生态位宽度主要反映物种对资源的利用程度[9],生态位越宽,物种对资源的利用程度越高[28]。本研究中,披针薹草与辽东栎的生态位宽度均较高,说明这2个物种对环境中资源的利用和适应能力较强。此外,本研究还发现,油松人工林下不同植物类型平均生态位宽度排序为乔木更新>藤本>草本>灌木,但不同植物类型间的生态位宽度差距较小,说明油松林下乔木物种存在一定的优势,但并不明显,通过森林经营管理可以进一步提升,即适度的去除非目标物种,可以保持林下常见乔木物种在群落中的较大生态位宽度和较强的生态适应性[1]。
物种由于群落中复杂的生态关系其生态位通常不是离散的,而是倾向于分享其他物种的基础生态位,结果导致2个或更多的物种对某些资源需求相似,进而使不同物种的生态位之间处于不同程度的重叠[26]。本研究发现,30个常见种总体生态位重叠较低,与张忠华等[29]研究结果相似,即群落的离散度较大。此外,生态位重叠可以用来衡量种间生态相似性或种间竞争,在资源不足的情况下,物种间的生态位重叠不仅可以反映物种间的生态形似程度,还反映了种间的竞争关系,而在资源充足情况下,生态位重叠并不反映竞争程度,仅表明这2个种间的生态相似性或占据了相近的生态空间[2]。本研究发现,油松天然更新与费菜、披针薹草、华山松、赤瓟等物种的生态位重叠较大,但联结系数分析结果显示,油松与以上物种的联结系数均大于0(即正联结),进一步说明中条山油松人工林下资源利用率低,与油松天然更新生态位重叠较大的物种均与其具有相似生态需求。本研究还发现,生态位越宽的物种,其与其他种间生态位重叠值相对较大,但生态位宽度较小的物种与其他种间生态位重叠值并不一定小,如三裂绣线菊与木梨,这与张国强等[30]研究结果相似,其可能是由于种间对资源的竞争作用,或者是物种间的促进作用使彼此的生态位得到扩张[31]。
基于本研究结果,发现物种间的竞争是影响油松天然更新的关键因素之一,并为该地区今后人工促进油松天然更新提供了一定的支撑,但人工林天然更新障碍除了受竞争的影响,还受环境、地被物、土壤、地形、干扰强度、种源、气候等方面的影响。此外,近年来,基于功能性状及功能多样性的研究方法已成为探索物种共存于生物多样性维持机制的一个新的突破口,并有力地推动了生物群落构建规则和恢复机理的研究[32-34],未来研究应该综合考虑,以便得出更深层次的结论,从而为人工林经营管理提供科技支撑。
-
中条山油松人工林下常见物种总体上显著正联结,而χ2检验结果显示无联结种对远高于有联结种对,说明常见种间联结较松散;联结系数分析结果显示,正联结种对大于负联结种对,而Pearson相关分析表明常见种对间大多数为正相关。此外,油松人工林群落稳定性交点坐标为(61.28,38.72),以上研究结果可以说明中条山油松人工林群落的成熟度相对较低,存在一些以竞争相同资源的物种,群落的总体资源利用率和稳定性较低。
油松天然更新与费菜、披针薹草、华山松、赤瓟等物种的生态位重叠较大,但联结系数分析结果显示,油松天然更新与以上物种正联结,说明油松天然更新与这几种植物资源利用相似或占据了相近的生态空间;土庄绣线菊、五角枫、粗齿铁线莲、青榨槭、金线草、披碱草与油松天然更新存在竞争,其可能是导致该区域油松天然更新障碍的主要原因之一,所以,今后在以油松为目标树种的森林经营管理中,应注重清除林下与油松天然更新竞争较强的物种。
种间关系对中条山油松人工林天然更新及群落稳定性的影响
Effects of Interspecific Relationship on Natural Regeneration and Community Stability of Pinus tabuliformis Plantation in Zhongtiao Mountain
-
摘要:
目的 为了揭示种间关系对中条山油松人工林天然更新及群落稳定性的影响,为油松人工林经营管理提供理论依据。 方法 基于中条山油松人工林群落样地调查数据,分析油松人工林下主要植物的种间关联性、生态位重叠及群落稳定性,并在此基础上探讨种间关联和生态位理论在油松人工林天然更新障碍及群落稳定性形成机制中的作用。 结果 中条山油松人工林下常见物种总体上显著正联结;χ2检验结果显示,无联结种对(372对)远高于有联结种对(34对),说明常见种间联结较松散;联结系数分析结果表明,正联结种对大于负联结种对,但显著正联结种对(AC≥0.20,145对)小于显著负联结种对(AC≤−0.20,168对),且与油松天然更新竞争最激烈的物种主要为土庄绣线菊、五角枫、粗齿铁线莲、青榨槭、金线草、披碱草。Pearson相关分析结果显示:正相关种对大于负相关种对,但显著相关种对远小于无显著相关种对。油松人工林下披针薹草的生态位最宽(23.47),红椋子最窄(3.87),而油松天然更新的生态位宽度为14.69,相对较高;常见种总体生态位重叠较低,但有11个种对生态位重叠指数大于0.98,说明部分种之间存在较大的生态相似性;油松天然更新与费菜、披针薹草、华山松、赤瓟等物种的生态位重叠较大,但联结系数分析结果显示,油松天然更新与以上物种正联结,说明油松天然更新与这几种植物资源利用相似或占据了相近的生态空间;油松人工林群落稳定性交点坐标为(61.28,38.72),显示群落稳定性较差。 结论 中条山油松人工林群落的成熟度相对较低,存在一些以竞争相同资源的物种,群落的总体资源利用率和稳定性较低。竞争可能是导致该区域油松天然更新障碍的主要原因之一,今后在以油松为目标树种的森林经营管理中,应注重清除林下与油松天然更新竞争较强的物种。 Abstract:Objective To examine the influence of interspecific relationship on the natural regeneration and community stability of Pinus tabuliformis plantation in Zhongtiao Mountain, and provide a theoretical basis for the management of P. tabuliformis plantation. Method Based on the survey data of the P. tabuliformis plantation community plots in Zhongtiao Mountain, the interspecific association, niche overlap and community stability of the dominant plants under the P. tabuliformis plantation were analyzed, and based on this, the role of interspecies association and niche theory in the formation of natural renewal barriers and community stability of P. tabuliformis plantations were discussed. Result The common species under the P. tabuliformis plantation in Zhongtiao Mountain are generally positively connected. The chi-square test results show that the unlinked species (372 pairs) are much higher than the linked species pairs (34 pairs), indicating that the common interspecies connections are loose. Association coefficient analysis results show that the positively connected species pairs are greater than the negatively connected species pairs, but the species pairs with significant positive association (AC≥0.20, 145 pairs) are smaller than those with significant negative association (AC≤−0.20, 168 pairs), and the most intensely competitive species with the natural regeneration of P. tabuliformis are Spiraea pubescens, Acer pictum subsp. mono, A. davidii, Clematis argentilucida, Rubia membranacea and Elymus dahurica. Pearson correlation analysis results show that the positively related species pairs are greater than the negatively related species pairs, but the significantly related species pairs are much smaller than the non-significantly related species pairs. Under the P. tabuliformis plantation, the niche of Carex lancifolia is the widest (23.47), and that of the Swida hemsleyi is the narrowest (3.87). The niche width of the natural regeneration of P. tabuliformis is 14.69, which is relatively high. The overall niche overlap of common species is low, but 11 species-pair niche overlap indexes are greater than 0.98, indicating that some species have greater ecological similarities. The niche of the natural regeneration of P. tabuliformis has the largest overlap with the niche of Sedum aizoon, Carex lancifolia, Pinus armandii and Thladiantha dubia, however, the results of the association coefficient analysis show that the natural regeneration of P. tabuliformis are positively associated with the species mentioned above, it shows that the natural regeneration of P. tabuliformis is similar to the resource utilization of these types of plants or it occupies a similar ecological space with these types of plants. The coordinate of the intersection point of the stability of the P. tabuliformis plantation community is (61.28, 38.72), which shows that the community stability is poor. Conclusion The maturity of the P. tabuliformis plantation community in Zhongtiao Mountain is relatively low, there are some species competing for the same resources, and the overall resource utilization and stability of the community is low. Competition may be one of the main reasons impeding the natural regeneration of P. tabuliformis in this area. Therefore, in the forest management activities targeting at P. tabuliformis species, more attention should be paid to removing the species that compete with the natural regeneration of P. tabuliformis under forest. -
图 1 人工促进油松天然更新
Figure 1. Artificial promotion of natural regeneration of P. tabuliformis
图 3 油松人工林群落Godron稳定性
Figure 3. Godron community stability in the community of P. tabuliformis plantation
表 1 油松人工林群落样地概况
Table 1. The information of community plots of P. tabuliformis Carr. plantation
序号
No.经度(E)
Longitude纬度(N)
Latitude树种组成
Species composition林龄
Age/a海拔
Elevation/
m样地面积
Area/
m2坡向
Slope aspect坡度
Slope/
(°)郁闭度
Coverage/
%平均胸径
Dq/
cm平均树高
Height/
m密度
Density/
(株∙ hm−2)1 111.92° 35.50° 6 油+4 华 40 1 916 1 200 西南 SW 22 80 24.08 14.89 808 2 111.92° 35.50° 6 油+4 辽 46 1 924 675 东南 SE 34 75 17.56 9.31 1 511 3 111.88° 35.50° 10 油 28 1 880 1 200 西 W 19 60 14.77 12.43 1 600 4 111.94° 35.51° 8 油+2 辽 38 1 750 1 200 西南 SW 36 55 18.66 11.20 925 5 111.93° 35.51° 8 油+2 华 48 1 722 1 200 东 E 21 85 25.64 15.46 508 6 111.90° 35.49° 9 油+1 华 48 1 933 1 200 东北 NE 13 80 27.84 16.31 583 7 111.90° 35.49° 10 油 41 1 917 1 200 西南 SW 20 60 27.79 14.74 500 8 111.94° 35.48° 8 油+2 辽 36 1 509 1 200 南 S 26 80 17.39 11.60 1 383 9 111.92° 35.51° 8 油+1 辽+1 华 39 1 997 1 200 西南 SW 25 75 14.79 9.07 2 475 10 111.89° 35.49° 9 油+1 落 32 1 872 1 200 东南 SE 11 70 17.39 12.95 1 250 11 111.90° 35.49° 10 油 48 1 928 1 200 西 W 8 60 29.32 15.94 467 12 111.90° 35.50° 7 油+2 华+1 辽 46 1 960 1 200 西南 SW 23 75 21.76 10.90 558  下载: 导出CSV
下载: 导出CSV
表 2 常见物种间关联分析结果
Table 2. Results of interspecific association analysis of common species
种间关联 Interspecific association 类型 Types 数值范围 Value range 种对数 Species pair number 百分比/% χ2 检验
Chi−square test无联结 No association p≥0.05 372 91.63 有联结 Association p<0.01 15 3.69 0.01≤p<0.05 19 4.68 总数 Total 406 100.00 联结系数
Association coefficient正联结 Positive association AC=1 26 6.40 0.60≤AC<1.00 23 5.67 0.20≤AC<0.60 96 23.65 0<AC<0.20 66 16.26 无联结 No association AC=0 1 0.25 负联结 Negative association −0.20<AC<0 26 6.40 −0.60<AC≤−0.20 73 17.98 −1.00<AC≤−0.60 23 5.67 AC=−1 72 17.73 总数 Total 406 100.00 Pearson相关
Pearson correlation正相关 Positive correlation p<0.01 50 11.49 0.01≤p<0.05 25 5.75 p≥0.05 139 31.95 负相关 Negative correlation p<0.01 1 0.23 0.01≤p<0.05 10 2.30 p≥0.05 210 48.28 总数 Total 435 100.00
下载: 导出CSV
表 3 基于Levins指数计算的常见种生态位宽度
Table 3. Calculating the niche width of common species based on Levins index
物种
Species生态位宽度
Niche width物种
Species生态位宽度
Niche width物种
Species生态位宽度
Niche width披针薹草 Carex lancifolia 23.47 青榨槭 Acer davidii 12.89 高山露珠草 Circaea alpina 8.09 辽东栎 Quercus wutaishansea 20.60 茜草 Rubia cordifolia 11.84 费菜 Sedum aizoon 7.99 华山松 Pinus armandii 18.79 五角枫 Acer pictum subsp. mono 11.69 南蛇藤 Celastrus orbiculatus 7.98 金线草 Rubia membranacea 17.15 土庄绣线菊 Spiraea pubescens 11.55 赤瓟 Thladiantha dubia 7.85 粗齿铁线莲 Clematis argentilucida 15.03 香薷 Elsholtzia ciliata 10.83 青麸杨 Rhus potaninii 7.07 油松 Pinus tabuliformis 14.69 早开堇菜 Viola prionantha 10.72 筋骨草 Ajuga ciliata 5.52 木梨 Pyrus xerophila 14.04 接骨木 Sambucus williamsii 10.41 鹅耳枥 Carpinus turczaninowii 5.31 太平花 Philadelphus pekinensis 14.02 毛丁香 Syringa tomentella 10.06 披碱草 Elymus dahuricus 5.18 榆叶梅 Amygdalus triloba 13.87 灰栒子 Cotoneaster acutifolius 9.82 三裂绣线菊 Spiraea trilobata 4.86 鞘柄菝葜 Smilax stans 13.08 唐古特忍冬 Lonicera tangutica 9.73 红椋子 Swida hemsleyi 3.87 注:表中乔木均为油松人工林下天然更新。
Notes: The trees in the table are natural regeneration under the P. tabuliformis Carr. plantation.
下载: 导出CSV
表 4 Levins生态位重叠统计
Table 4. Statistics of Levins niche overlap
项目 Item Oik≥0.98 0.78≤Oik<0.98 0.59≤Oik<0.78 0.40≤Oik<0.59 0.21≤Oik<0.40 Oik≤0.21 种对数 Species pair number 11 36 50 87 115 136 百分比/% 2.53 8.28 11.49 20.00 26.44 31.26
下载: 导出CSV
-
[1] 贾呈鑫卓, 李帅锋, 苏建荣, 等. 择罚对思茅松天然林乔木种间与种内关系的影响[J]. 植物生态学报, 2014, 38(12):1296-1306. [2] 钟 宇, 张 健, 刘泉波, 等. 巨桉人工林草本层主要种群的生态位分析[J]. 草叶学报, 2010, 19(4):16-21. [3] 毛子昆, 郝占庆, 原作强, 等. 物种聚集分布与种间关系的多度不对称性[J]. 中国科学:生命科学, 2020, 50(4):381-390. [4] 牛克昌, 刘怿宁, 沈泽昊, 等. 群落构建的中性理论和生态位理论[J]. 生物多样性, 2009, 17(6):579-593. [5] 柴永福, 岳 明. 植物群落构建机制研究进展[J]. 生态学报, 2016, 36(15):4557-4572. [6] 储诚进, 王酉石, 刘 宇, 等. 物种共存理论研究进展[J]. 生物多样性, 2017, 25(4):345-354. doi: 10.17520/biods.2017034 [7] 周先叶, 王伯荪, 李鸣逛, 等. 广东黑市顶自然保护区森林次生演替过程中群落的种间联结性分析[J]. 植物生态学报, 2000, 24(3):332-339. doi: 10.3321/j.issn:1005-264X.2000.03.015 [8] 简敏菲, 刘琪璟, 朱 笃, 等. 九连山常绿阔叶林乔木优势种群的种间关联性分析[J]. 植物生态学报, 2009, 33(4):672-680. doi: 10.3773/j.issn.1005-264x.2009.04.005 [9] 钱逸凡, 伊力塔, 胡军飞, 等. 普陀山主要植物种生态位特征[J]. 生态学杂志, 2012, 32(3):561-568. [10] 徐化成, 孙肇风, 郭广荣, 等. 油松天然林的地理分布和种源区的划分[J]. 林业科学, 1981, 17(3):258-270. [11] 刘铁岩, 毕 君, 王 超, 等. 冀北山地油松人工林天然更新研究[J]. 中南林业科技大学学报, 2017, 37(7):55-58,65. [12] 魏 曦, 梁文俊, 毕华兴, 等. 晋西黄土区油松林分结构与水土保持功能的多因子复合关系[J]. 林业科学研究, 2020, 33(3):39-47. [13] 牛 健. 影响北京市油松人工林天然更新的几项因子研究[D]. 北京: 北京林业大学, 2018. [14] 傅志军, 郑雪婷. 山西中条山植物区系的特征[J]. 西北植物学报, 1994, 14(2):148-152. doi: 10.3321/j.issn:1000-4025.1994.02.012 [15] 王 好, 康慕谊, 刘全儒, 等. 中条山植物区系与植被研究进展[J]. 北京师范大学学报:自然科学版, 2004, 40(5):676-683. [16] 上官铁梁, 张 峰. 中条山木本植物区系地理成分分析[J]. 植物研究, 2000, 20(2):143-155. doi: 10.3969/j.issn.1673-5102.2000.02.005 [17] Yates F. Contingency tables involving small numbers and the χ2 test[J]. Journal of the Royal Statistical Society, 1934, 1(2): 217-235. [18] Chacón-Labella J, Cruz M, Escudero C, et al. Evidence for a stochastic geometry of biodiversity: the effects of species abundance, richness and intraspecific clustering[J]. J Ecol, 2017, 105(2): 382-390. doi: 10.1111/1365-2745.12710 [19] Lennon J J, Koleff P, Greenwood J J, Gaston K J. Contribution of rarity and commonness to patterns of species richness[J]. Ecology Letters, 2004, 7(2): 81-87. [20] 郑元润. 森林群落稳定性研究方法初探[J]. 林业科学, 2008, 36(5):28-32. [21] Gu L, Gong Z W, Li W Z. Niches and interspecific associations of dominant populations in three changed stages of natural secondary forests on loess plateau, P. R. China[J]. Scientific Reports, 2017, 7(1): 6604-6615. doi: 10.1038/s41598-017-06689-9 [22] 刘润红, 陈 乐, 涂洪润, 等. 桂林岩溶石山青冈群落灌木层主要物种生态位与种间联结[J]. 生态学报, 2020, 40(6):2057-2071. [23] He D, Biswas S R. Negative relationship between interspecies spatial association and trait dissimilarity[J]. Oikos, 2018, 128(5): 1-9. [24] Greig-Smith P. Quantitative plant ecology. 3rd ed[M]. Oxford: Blackwell Scientific Publications, 1983. [25] Kershaw K A, Looney J H. Quantitative and dynamic plant ecology. 3rd ed[M]. London: Edward Arnold Limited, 1985. [26] 张金屯. 数量生态学[M]. 北京: 科学出版社, 2004. [27] Zhang W P, Liu G C, Sun J H, et al. Temporal dynamics of nutrient uptake by neighbouring plant species: evidence from intercropping[J]. Functional Ecology, 2017, 31(2): 469-479. doi: 10.1111/1365-2435.12732 [28] Gross N, Bagousse-Pinguet Y L, Liancourt P, et al. Functional trait diversity maximizes ecosystem multifunctionality[J]. Nature Ecology & Evolution, 2017, 1(5): 132. [29] 张忠华, 梁士楚, 胡 刚. 桂林岩溶石山阴香群落主要种群生态位研究[J]. 林业科学研究, 2009, 22(1):63-68. doi: 10.3321/j.issn:1001-1498.2009.01.011 [30] 张国强, 巨天珍, 刘文桢, 等. 天水小陇山红豆杉群落特征的生态学意义[J]. 林业科学研究, 2020, 33(5):184-190. [31] Pulla S, Suresh H S, Dattaraja H S, et al. Multidimensional tree niches in a tropical dry forest[J]. Ecology, 2017, 98(5): 1334-1348. doi: 10.1002/ecy.1788 [32] Díaz S, Lavorel S, de Bello F, et al. Incorporating plant functional diversity effects in ecosystem service assessments[J]. Proceedings of the National Academy of Sciences, 2007, 104(52): 20684-20689. doi: 10.1073/pnas.0704716104 [33] Lavorel S. Plant functional effects on ecosystem services[J]. Journal of ecolgoy, 2013, 101(1): 4-8. doi: 10.1111/1365-2745.12031 [34] Funk J L, Cleland E E, Suding K N, et al. Restoration through reassembly: plant traits and invasion resistance[J]. Trands in ecology and evolution, 2008, 23(12): 695-703. doi: 10.1016/j.tree.2008.07.013 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 4354
- HTML全文浏览量: 2482
- PDF下载量: 63
- 被引次数: 0
