




-
植物激素茉莉酸(JA)是脂肪酸衍生的羟脂素,在植物生长发育和逆境胁迫响应等过程中发挥重要作用[1]。JA通过其信号转导途径调控与多种激素互作或调控下游响应基因参与多种生物学过程。JA信号途径是一个由多基因参与的复杂调控过程[2]。 Coronatine insensitive 1 (COI1)是JA信号的受体,与JA结合后促进转录抑制因子jasmonate ZIM-domain (JAZ)家族蛋白泛素化并被26S蛋白酶体降解,从而解除JAZ对转录因子髓细胞组织增生蛋白(Myelocytomatosis proteins,MYCs)MYC2的抑制作用,最终激活下游响应基因的表达[3]。植物中MYC是JA信号途径中关键转录因子,通过形成COI/JAZs/MYC复合物发挥调控作用,参与多种激素信号转导过程[4]。
MYC转录因子含有保守的bHLH结构域,属于bHLH类转录因子家族。bHLH结构域由碱性域与HLH域组成, N端碱性域具有DNA结合功能,C端HLH域参与bHLH蛋白二聚化形成同源或异源二聚复合体[5]。bHLH类转录因子是转录因子中的超级家族,在拟南芥(Arabidopsis thaliana (L.) Heynh.)中共有158个家族成员,分为26个亚家族,其中,MYC转录因子分布于亚家族III [6],包含8个成员。已有研究证明,MYC转录因子在植物生长发育、次生代谢物的合成、逆境响应、植物激素信号转导及不同激素信号通路之间的相互作用中发挥关键作用[4,7-8]。MYC2是MYC类转录因子中研究最透彻的一个,在JA信号通路中发挥关键调控作用。在拟南芥根系发育过程中,MYC2可以直接结合到维持根尖细胞活性的关键转录因子PLT1和PLT2的启动子上抑制其表达,进而抑制拟南芥主根分生区细胞分裂[9]。在蓝光诱导下,拟南芥MYC2/MYC4可以结合NST1启动子激活其表达,进而调控次生壁合成,促进花序轴纤维细胞次生壁加厚[10]。改变红光与远红光比例,光敏色素互作因子PIF4能够与MYC2/MYC4直接相互作用影响MYC蛋白核定位,抑制MYC2/MYC4对NST1的激活进而影响细胞次生壁加厚[11]。在花发育过程中,myc2/3/4三突变体及myc2/3、myc2/4双突变体表现出早开花表型[12],同时伴有花萼发育延迟、花药不能正常开裂或开裂延迟以及花粉粒存活率低等表型[13]。此外,MYC2参与多种植物激素间的调控,包括生长素、细胞分裂素、乙烯等。MYC2可以与乙烯不敏感转录因子EIN3相互作用,抑制EIN3的转录活性,从而抑制HLS1的表达,最终影响拟南芥黄化苗顶端弯钩的形成[14]。在果实成熟方面,MYC2转录因子能通过结合苹果乙烯合成途径中的ACC合成酶基因和ACC氧化酶基因启动子调控乙烯合成进而影响果实成熟[15]。最近研究表明,MYC2可以通过抑制细胞分裂素氧化酶/脱氢酶(CKX)基因来提高细胞分裂素含量,从而抑制不定根的起始[16]。综上所述,MYC类转录因子对植物生长发育及生物和非生物胁迫响应具有重要调控作用。
尽管MYC基因家族已在拟南芥、水稻(Oryza sativa L.)等模式植物中研究得比较深入,但在林木中其功能研究还鲜有报道。杨树(Populus L.)是重要的人工林树种,而且具有基因组小、适应性强、遗传转化体系成熟等特性,已成为林木分子育种研究的模式树种。因此,本文系统分析了杨树MYC家族成员基因结构、蛋白保守结构域、组织表达特性及不同激素和胁迫响应模式,研究结果为进一步解析杨树MYC基因的生物学功能奠定了基础。
-
在拟南芥基因组网站TAIR(TAIR - Home Page (arabidopsis.org))数据库中获取MYC基因家族8个成员的氨基酸序列,并以此为靶序列,在Phytozome 网站(
https://phytozome-next.jgi.doe.gov/ )中通过BLASTP方法比对检索毛果杨基因组数据库V4.1,获得MYC直系同源氨基酸序列。利用ExPASy(http://www.expasy.org/ )在线工具分析杨树MYC蛋白基本理化性质。使用WoLEPSORT(WoLF PSORT: Protein Subcellular Localization Prediction Tool (genscript.com))在线工具分析杨树MYC蛋白的亚细胞定位情况。 -
参考MEGA7.0[17],利用MEGA11中ClustalW程序,将毛果杨、拟南芥MYC蛋白序列进行多重序列比对;采用Neighbor-Joining(NJ)法,模型为p-distance,成对删除,校验参数为1000,构建系统发育进化树。iTOL(
https://itol.embl.de/ )在线网站进一步完善进化树。 -
杨树MYC基因结构用GSDS2.0 (
http://gsds.cbi.pku.edu.cn/ )软件分析。杨树MYC蛋白结构域使用MEME(MEME - Submission form (meme-suite.org))在线分析,最大motif检索数为10,其余为默认参数。 -
实验所用材料84K杨(Populus alba × P. glandulosa)由林木遗传育种全国重点实验室保存。RNA提取使用(RNA Easy Fast Plant Tissue Kit)试剂盒。分别提取温室培养2个月 84K杨的根、茎、叶以及茎的不同部位的总RNA,反转录合成cDNA第一条链,并以cDNA为模板进行实时荧光定量PCR分析(qRT-PCR)。反应体系为TB Green® Premix EX Taq ™ Ⅱ 5 μL,正向引物(10 μmol·L−1)0.5 μL,反向引物(10 μmol·L−1)0.5 μL,cDNA 1 μL,超纯水3 μL;qRT-PCR程序为:95 ℃ 30 s;95 ℃ 5 s;60 ℃ 30 s;50 ℃ 30 s,45个循环。引物序列见表1。
引物
Primer序列(5′−3′)
Primer sequence(5′−3′)引物
Primer序列(5′−3′)
Primer sequence(5′−3′)PtrMYC2a-F TATGCCAATGTGTCTGTGATGAATG PtrMYC2a-R TCTTGCGTGTAAAACCGACTCC PtrMYC2b-F AATGCTGGGATTCCGTCAACTAC PtrMYC2b-R TGATGATTGTTATTGTTCGCAGTCT PtrbHLH14a-F GTGGTCCCGAATGTGTCAAAGATG PtrbHLH14a-R ATGTTCGCATTACCAGAGATTTTTG PtrbHLH14b-F TACTCAAGCAAGGTTAGCAACAAGC PtrbHLH14b-R ATCAACCAATCTCTCCAGGTCCAT PtrbHLH14c-F GCTCGTGAAACAGAAGAAGCAAAT PtrbHLH14c-R TAGAGGAAAGTCCGAGTCTGAATGC PtrbHLH14d-F TGACACTATGGACAATCAAAGCACC PtrbHLH14d-R ATTCACATTATCGGATTGGACCCTA PtrbHLH3a-F CAACAAGCACGAATCAAGTTTATGG PtrbHLH3a-R GGACAAGTCATCATTTGGCTGC PtrbHLH3b-F GGAGTCTCACCCCGTTTCTACC PtrbHLH3b-R CAGTCTTGAGCAGTGATTTGATGTT PtrbHLH17a-F CAAGGGACACCAGGTGAGGTTTAT PtrbHLH17a-R CCTGCTTTTGACCTTGGTAGTGATT PtrbHLH17b-F AGGCTTCATTGTTGGGAGATGCTAT PtrbHLH17b-R AATGCTCCCCGACTTTTCCCTT PtrActin-F AAACTGTAATGGTCCTCCCTCCG PtrActin-R GCATCATCACAATCACTCTCCGA Table 1. The primers used for quantitative real-time PCR
-
激素处理:选择生长4周的84K组培苗,分别进行生长素(10 μmol·L−1 NAA)、细胞分裂素(10 μmol·L−1 6-BA)和茉莉酸(100 μmol·L−1 MeJA)处理。处理时间为:生长素(0.5、1、2、3、6 h)、细胞分裂素(0.5、1、2、3 h)、茉莉酸(1、3、6、9、12 h)。每个处理包含3次生物学重复,每次重复包含3株独立植株,取样位置为顶端向下第4片叶,以未处理的84K杨作为对照。
非生物胁迫处理:84K杨温室培养4周后,分别进行4 ℃、20% PEG6000及200 mmol·L−1 NaCl处理,处理3、6、12、24 h后取顶端第4片叶,每处理重复3次,每重复3株植株混合取样,以未处理84K杨作为对照。
使用RNA Easy Fast Plant Tissue Kit试剂盒提取处理后杨树叶片RNA,并通过反转录试剂盒合成单链cDNA。使用Primer Premier 5软件,在杨树MYC基因序列的非保守区设计qRT-PCR引物(表1),引物序列的Tm值为60~62 ℃,并由生工生物公司合成。qRT-PCR使用TB Green® Premix EX Taq ™ Ⅱ(TaKaRa Dalian,China)试剂盒,并根据Roche480实时荧光定量PCR仪说明书进行操作。反应体系为:TB Green® Premix EX Taq ™ Ⅱ 5 μL,上下游引物各0.5 μL,cDNA 1 μL,超纯水3 μL。qRT-PCR所用内参基因为杨树Actin基因(基因序列号:Potri.001G309500)。每个模板设置4个重复,所有实验均进行3次生物学重复,采用2−ΔΔct方法进行定量数据分析。
-
利用Phytozome网站BLAST工具查找拟南芥MYCs在毛果杨基因组中同源基因,共筛选获得10个杨树MYC基因成员(表2),编码的蛋白质序列长度为466~660 aa,蛋白质分子量介于52.34~72.09 kDa,等电点为5.38~7.72。亚细胞定位预测结果显示:10个MYC蛋白亚细胞定位不同,其中,6个定位在细胞核,2个定位在细胞核和细胞质, 2个定位在细胞核与叶绿体。基于鉴定出的毛果杨PtrMYCs以及拟南芥MYC基因家族编码的氨基酸序列,使用NJ法构建系统发育进化树,结果(图1)显示:与拟南芥MYC基因家族类似,毛果杨MYC基因家族分为3个分枝,但不同分枝成员数量与拟南芥存在较大差异。分枝Ⅰ包含拟南芥MYC2/3/4/5共4个基因,但杨树中只有2个同源基因,分别命名为PtrMYC2a和PtrMYC2b;分枝Ⅱ包含拟南芥AtbHLH14及杨树4个同源基因,分别命名为PtrbHLH14a~14d;分枝Ⅲ包括拟南芥AtbHLH3和AtbHLH17,每个基因在杨树中均有2个同源基因,命名为PtrbHLH3a、PtrbHLH3b和PtrbHLH17a、PtrbHLH17b。
基因名称
Gene基因ID
Gene number基因在染色体位置
Location蛋白质长度
Protein length/aa等电点
PI分子量
MW/kDa亚细胞定位
Subcellular localizationPtrMYC2a Potri.001G142200 Chr01( + )11649287-11651987 648 5.38 71.11 细胞核 PtrMYC2b Potri.003G092200 Chr03( − )11839709-11842763 660 5.49 72.09 细胞核 PtrBHLH14a Potri.001G083500 Chr01( − )6613720-6615721 472 7.17 52.73 细胞核/细胞质 PtrBHLH14b Potri.003G147300 Chr03( + )16158005-16160025 466 7.72 52.34 细胞质/细胞核 PtrBHLH14c Potri.002G176900 Chr02( + )13775139-13776876 492 5.40 54.70 细胞核 PtrBHLH14d Potri.014G103700 Chr14( + )6950999-6952650 494 5.91 54.87 细胞核/叶绿体 PtrBHLH3a Potri.002G042000 Chr02( − )2756159-2758458 504 6.59 54.92 细胞核/叶绿体 PtrBHLH3b Potri.005G221100 Chr05( + )22329285-22332096 505 6.13 55.44 细胞核 PtrBHLH17a Potri.002G172101 Chr02( − )13241064-13244032 619 6.14 68.25 细胞核 PtrBHLH17b Potri.014G099700 Chr14( − )6539405-6542974 620 6.16 68.02 细胞核 Table 2. Basic information of MYC family members in Populus

Figure 1. Phylogenetic tree of MYC family members between Populus and Arabidopsis
-
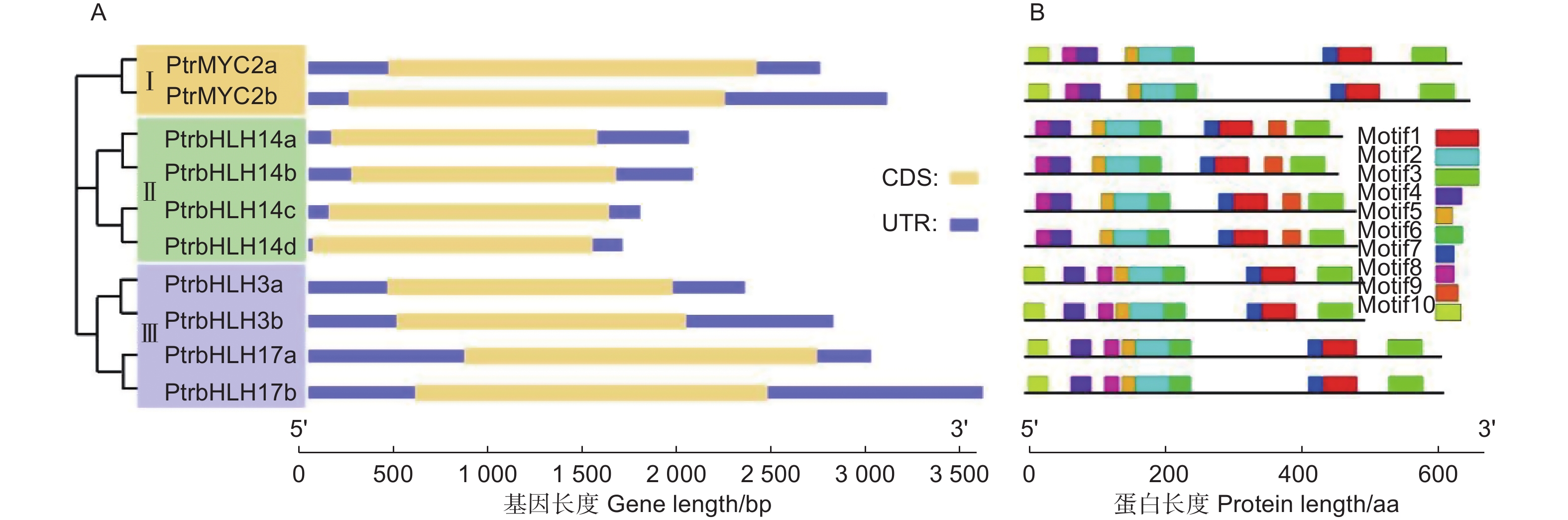
为了了解PtrMYC基因的结构特征,对其基因结构及保守基序进行分析,发现杨树所有MYC基因均不含内含子(图2A)。使用MEME软件对杨树MYC蛋白进行保守结构域分析,结果表明:所有MYC蛋白的结构域在进化上是高度保守的,Motif1、2、3、4、5、6、7、8存在于所有家族成员中,其中,Motif1、3、7和2、4、5、6、8为该家族基因保守的特征结构域即bHLH和bHLH-MYC_N。另外,有的家族分枝具有或缺失某些蛋白基序,如Motif9仅存在于分枝Ⅱ的家族成员中,Motif10仅存在于分枝Ⅰ和Ⅲ的成员中,大多数同源配对成员有高度相似的结构域组成,提示可能在基因功能上存在冗余(图2B)。

Figure 2. Gene structure (A) and distribution of conversed motifs (B) of PtrMYC family members in Populus
-
基因组织特异性表达模式研究对预测基因功能具有重要的作用。为了探讨PtrMYC基因家族在杨树生长发育中的功能,本研究利用qRT-PCR方法分析了MYC基因各成员在不同器官和组织的表达特异性,结果(图3)显示:不同MYC家族成员具有不同的组织表达特异性,但基因对之间多存在类似的表达模式。除PtrbHLH3a外,其余9个PtrMYC基因在根中均有较高表达量,说明大部分MYC家族成员可能在根系发育过程中发挥作用;在从顶端到基部的不同茎段中,PtrbHLH14a和PtrbHLH14b这一组基因对表达量呈现先上升后下降的趋势,说明它们可能在茎由初生生长向次生生长发育过程中具有重要调节作用;PtrbHLH14c和PtrbHLH14d这组基因对表达量则呈现下降的趋势,说明可能在顶端分生组织发育中具有重要的作用。杨树MYC家族成员在不同器官和组织存在显著表达差异,说明不同成员可能在杨树生长发育不同生物学过程中发挥调控作用。
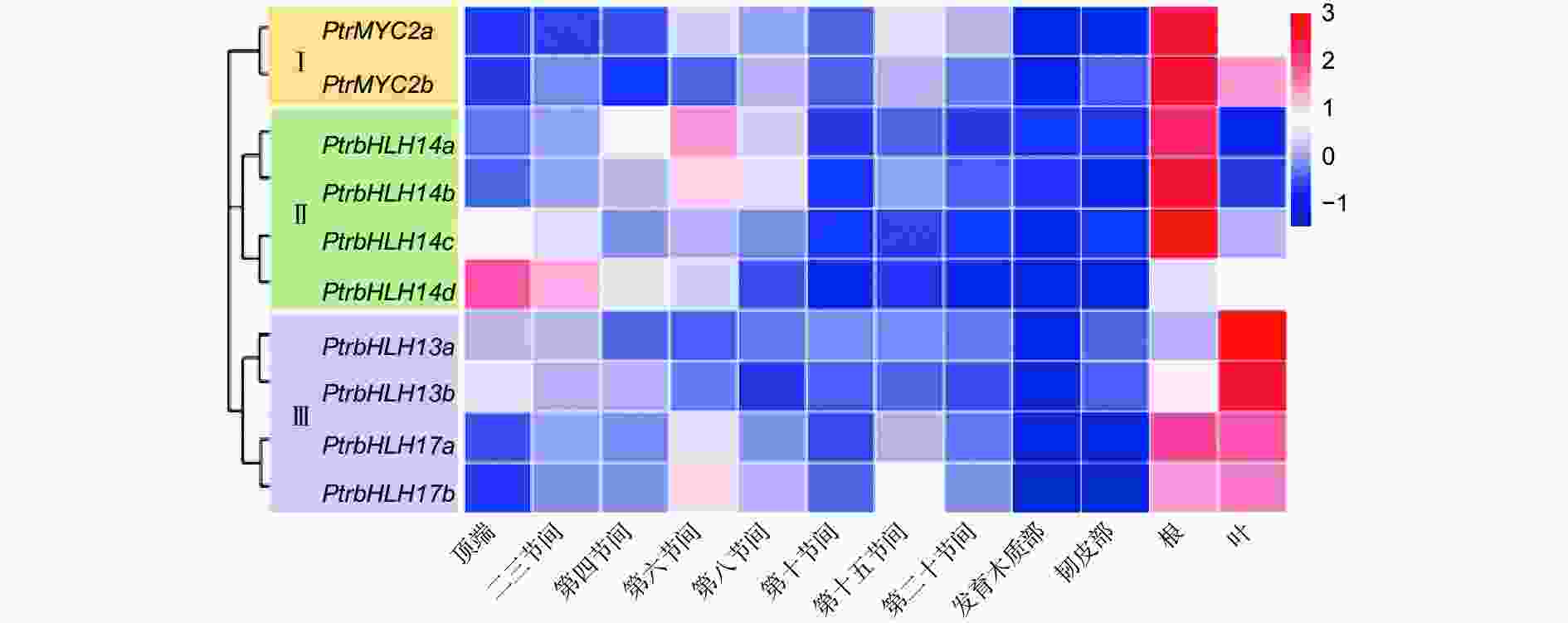
Figure 3. Expression analyses of MYCs in Populus
-
为进一步了解杨树MYC家族基因对不同植物激素的响应情况,本研究利用qRT-PCR方法检测了其对生长素( NAA)、细胞分裂素(6-BA)、茉莉酸( MeJA)响应的表达模式(图4)。在NAA、6-BA、MeJA处理后,杨树MYC基因家族各成员表达模式各不相同。PtrbHLH14a、PtrbHLH14b基因表达量受生长素诱导0.5 h后开始下调,表明生长素可以快速抑制特异MYC基因的表达。细胞分裂素处理0.5 h后, PtrbHLH14a、PtrbHLH14b、PtrbHLH14d基因表达量显著上调,说明它们可以快速响应细胞分裂素处理。大部分杨树MYC基因都能响应茉莉酸信号,茉莉酸处理后, PtrMYC2a、PtrMYC2b、PtrbHLH14c、PtrbHLH14d、PtrbHLH17a、PtrbHLH17b基因表达量显著上调。值得注意的是,PtrbHLH14a和PtrbHLH14b基因对能够响应生长素和细胞分裂素处理,但表达模式呈相反趋势,说明它们可能同时参与生长素与细胞分裂素信号通路。综上所述,MYC家族成员可能广泛参与不同激素调控植物生长发育的多个过程。
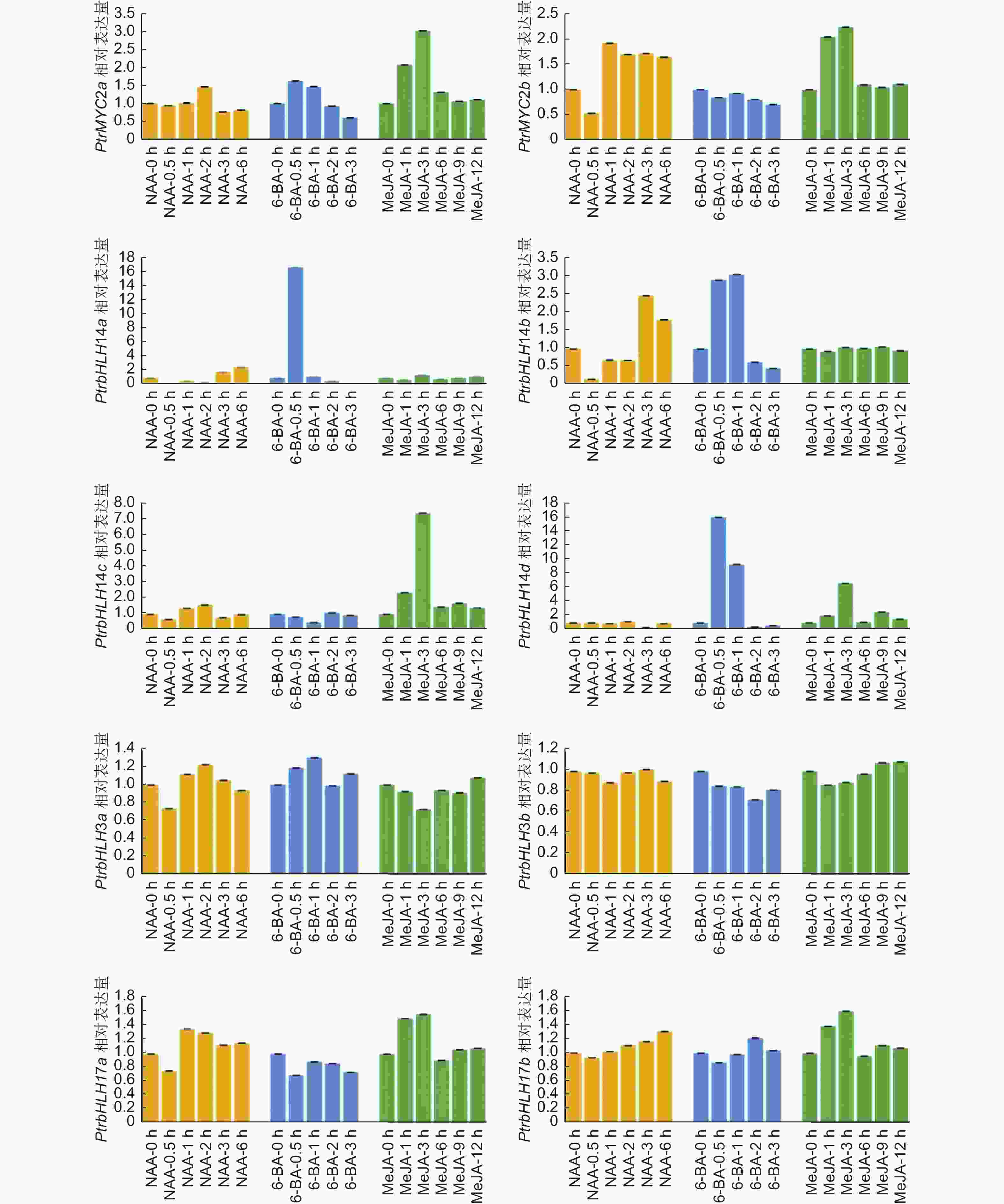
Figure 4. Expression patterns of PtrMYCs under NAA、6-BA、MeJA treatment
-
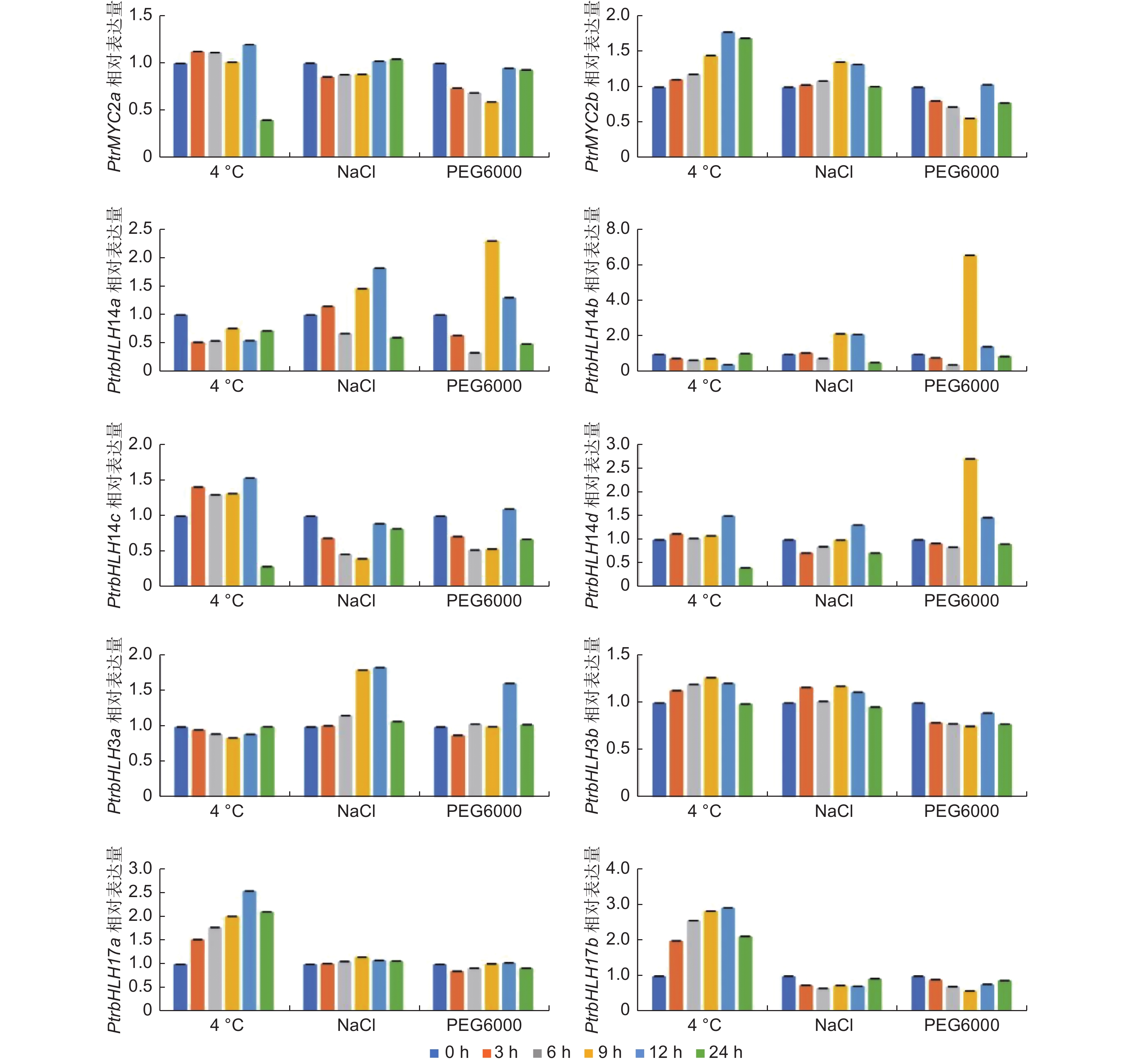
为揭示杨树MYC基因对非生物胁迫的响应机制,本研究利用qRT-PCR方法检测了该家族基因在4 ℃、PEG6000和NaCl胁迫下的表达模式(图5)。在200 mmol·L−1 NaCl、20% PEG6000及4 ℃低温处理后,杨树MYC基因家族各成员表达都受到不同程度的诱导,且大部分基因表达量在处理后12 h达到高峰。低温处理可以诱导6个杨树MYC基因上调表达,其中PtrbHLH17a、PtrbHLH17b两个基因受低温诱导上调表达最多,且在12 h达到峰值;PtrbHLH14a、PtrbHLH14b下调表达,说明不同MYC基因响应低温表达方式不同。在NaCl处理之后,PtrMYC2b、PtrbHLH14b、PtrbHLH3a、PtrbHLH3b、PtrbHLH14a、PtrbHLH14d基因的表达量先上调再下调,而PtrMYC2a、PtrbHLH14c、PtrbHLH17b基因则呈现相反表达模式。干旱处理后,PtrbHLH14a、PtrbHLH14b、PtrbHLH14d、PtrbHLH3a基因表达量显著上调,其中,PtrbHLH3a表达量在处理后12 h时达到高峰,其它3个基因在9 h时达到高峰,说明不同MYC基因响应干旱时间不同。另外,同一分支基因响应相同非生物胁迫时具有相似表达模式,说明基因功能存在冗余。综上所述,MYC家族成员在不同非生物胁迫中表达情况存在差异,推测在植物响应逆境胁迫中发挥不同作用。

Figure 5. Expression patterns of PtrMYCs under 4 ℃, NaCl and PEG6000 treatment
-
林木在长期进化过程中形成了自己独特的生物学特征和环境适应性,而基因组学和基因功能研究发现,不同物种同源基因可能在功能上存在较大差异,因此,在模式树种杨树中开展基因功能研究对揭示林木特有的生物学过程具有重要意义[18]。MYC基因家族是bHLH基因家族一个亚家族,对植物的生长发育及环境适应性具有重要作用。目前,在拟南芥中对MYC2基因进行了较为深入的研究,发现该基因不仅能参与调控根、叶、种子等器官的发育活动,并且还以调控开关的角色存在于多种激素通路互作过程中[5]。然而,模式树木毛果杨中,MYC家族基因基本特征及其逆境胁迫响应特性尚无报导。本研究根据拟南芥MYC基因家族氨基酸序列,在毛果杨基因组中通过序列比对获得了杨树MYC基因家族成员,并对其基因结构、组织表达特异性及对激素和逆境响应表达模式等进行了系统分析。在毛果杨基因组中共鉴定出10个MYC成员,系统进化树分析发现该基因家族分为3个分枝,其中,分枝Ⅰ包含2个成员,分枝Ⅱ有4个成员,分枝Ⅲ有4个成员,这与拟南芥中各分枝成员数量相差较大,说明MYC基因家族不同分枝在进化过程中可能存在基因扩张或丢失现象。蛋白结构域分析发现,PtrMYC蛋白含有多个基序,10个Motif中有8个Motif存在于所有MYC成员中(图2B),表明PtrMYC成员之间在蛋白结构上高度保守。
基因组织表达特异性对进一步探究其功能具有重要作用。本研究中,杨树PtrMYC在不同器官和组织存在明显表达差异,表明它们可能在不同生长发育过程中发挥作用。拟南芥中研究发现,MYC2抑制主根分生区细胞分裂[9],同时促进侧根形成[19]。另外,MYC2可以通过调控细胞分裂素含量抑制不定根起始[16],说明MYC2转录因子对根系发育的调控是多方面的。组织表达分析表明,PtrMYC家族9个成员在根部表达水平较高,暗示它们可能参与杨树根系不同发育过程。在叶片发育过程中,MYC2能通过抑制生长素合成进而抑制叶脉发育[20]。Qi等[13]研究表明,MYC2/MYC3/MYC4在激活叶片衰老相关基因SAG29诱导叶片衰老中存在功能冗余,而bHLH03、bHLH13、bHLH14和bHLH17与SAG29的启动子结合抑制其表达,拮抗MYC2/MYC3/MYC4诱导的叶片衰老[21]。另外,MYC2-Dof2.1形成前馈机制促进叶片衰老[22]。本研究中发现,PtrbHLH3a、PtrbHLH3b、PtrbHLH17a、PtrbHLH17b等在叶片中高丰度表达,可能参与杨树叶片发育和衰老过程的调控。
植物激素之间的相互作用组成一个非常复杂的信号调控网络,在植物生长发育和逆境响应过程中起着非常重要的调控作用[23] 。MYC基因在不同激素之间互作过程中具有重要作用。在不定根和叶片发育过程中,MYC家族基因能够抑制细胞分裂素代谢和生长素合成影响不定根起始和叶脉发育 [16, 20]。拟南芥中,MYC2可以与其靶基因ERD1的启动子结合,激活其表达,进而激活JA诱导的脱水应激反应[24]。此外,茉莉酸和生长素协同作用调控拟南芥热形态建成[25]。在苹果愈伤组织中过表达MdMYC2增加了MdCIbHLH1、MdCBF1、MdCBF2和MdCBF3的表达水平,导致苹果抗冻耐受性显著提高[26]。在番茄中MYC2激活乙烯响应因子SlERF.B8的转录,进而增强JA信号,增加了番茄耐寒性[27]。本研究发现,不同PtrMYC成员响应激素和非生物胁迫方式不同,一些MYC基因既受激素诱导,又响应某种非生物胁迫,如PtrMYC14a、PtbHLH14b表达可以被生长素抑制,也可以被细胞分裂素诱导,同时还可以响应盐和干旱胁迫;而PtrMYC2b表达能够被茉莉酸诱导,同时还可以响应低温和盐胁迫,提示MYC基因家族可能参与不同植物激素相互作用,进而广泛参与到植物生长发育和非生物胁迫响应中,且不同成员可能参与不同生物学过程,但其具体调控机制尚需进一步研究。
-
本文系统分析了杨树MYC基因家族的基因和蛋白结构、组织表达特异性以及响应激素和逆境的表达模式。杨树基因组中共有10个MYC成员,均含有bHLH转录因子典型结构域;组织表达分析发现,杨树MYC基因家族成员表达模式各异,可能参与了不同的生物学过程的调控。在不同的植物激素处理下,杨树MYC家族成员表达模式多样,表明不同的MYC基因可能参与到不同的植物激素信号通路。在不同的非生物胁迫下,杨树MYC基因家族各成员的表达情况也不同,表明杨树MYC基因在不同的非生物胁迫下发挥不同的功能。本研究为揭示MYC基因在杨树生长发育和胁迫响应过程中的基因功能奠定了基础。
Analysis of Expression Patterns of MYC Family Members in Populus
- Received Date: 2022-10-26
- Accepted Date: 2022-12-27
- Available Online: 2023-06-20
Abstract:

 DownLoad:
DownLoad: