




-
全球气候变化引发降水模式改变和气温升高,导致极端事件变得更加频繁、强烈和持续时间更长[1]。干旱区是全球生态系统最脆弱的区域之一,也是响应气候变化的敏感区域,因此植物的生理特征、形态结构、生物量等功能性状极易受到环境改变的影响,因此,评估干旱区植物功能性状的变化有助于了解植物对干旱环境变化的响应以及资源的利用策略[2-3]。由于长时间暴露在不断变化的自然环境之中,固着生长的植物可通过表型、生理和遗传等方面的有效进化以适应环境因子的改变[4-6]。大量研究表明,植物功能性状是全球生态系统对气候变化的可度量特征,植物可以通过其自身功能性状的改变来适应外界环境波动的影响[7],这对研究生态系统功能响应环境变化具有重要意义[8-9]。叶片是植物光合作用的主要器官,是生态系统中主要物质的能量转换器[10-11],促进大气-植物系统的能量交换[12]。叶片性状通过影响植物生长、繁殖和存活而间接影响植物适应性的形态、生理和物候特征[13],可作为连接植物与环境的纽带 [14],通常被用于解析植物与植物之间、植物与环境之间的相互作用[15-16]。已有研究表明株高(H)、叶面积(LA)、比叶面积(SLA)、叶片干物质含量(LDMC)、叶片氮含量(N)等功能性状的改变能够直观地反映物种对气候变化的响应,有效减缓或者遏制环境因素造成的不利影响[17-20]。
中间锦鸡儿(Caragana intermedia Kuang et H. C. Fu)根系发达、叶片小且多具被毛,有良好的保水作用和较强的适应性和抗逆性,是我国北方干旱半干旱地区植被恢复、防风固沙和水土保持的先锋树种。同时,该物种也是优良的饲料植物,生态和经济价值突出[21-23]。目前关于该物种的研究主要集中在生长特性[24]、形态解剖[25]、遗传多样性[26]、非生物胁迫相关基因功能验证和根际间微生物的功能多样性[27]等方面。有研究发现,随着中间锦鸡儿种植年限的增加,生态系统功能会受到功能性状的影响,其中基径、单株枝干质量、单株叶干质量对地上生物量均具有重要响应[28]。但关于不同种植年限中间锦鸡儿叶片形态与结构功能性状的研究报道较少,基于此,本研究以4个不同种植年限中间锦鸡儿人工林为研究对象,通过解析中间锦鸡儿叶片形态与结构性状随着种植年限增加的变化特征及规律,探究各叶片功能性状之间的组合与权衡,为探明中间锦鸡儿响应环境变化的生存策略提供理论依据。
-
采样地点为盐池县皖纪沟村(37°57′23″ N,106°77′99″ E),气候为典型的温带大陆性气候,年平均气温8.1 ℃,无霜期165 d,年平均降水量250~350 mm,且由东南向西北递减。土壤类型以灰钙土和风沙土为主,自然植被类型以荒漠植被为主,主要优势物种有达乌里胡枝子(Lespedeza davurica (Laxm.)Schindl.)、短花针茅(Stipa breviflora Griseb.)、白草(Pennisetum centrasiaticum Tzvel.)、赖草(Leymus secalinus (Georgi) Tzvel.)、老瓜头(Cynanchum komarovii AI. Iljinski)、苦豆子(Sophora alopecuroides L.)、绵蓬(Corispermum hyssopifolium L.)等。荒漠化长期困扰着当地以牧业为主体的生产经营活动,为此,从上世纪70年代开始种植中间锦鸡儿,面积逐年扩大。
-
采样地点为盐池县皖纪沟村,该地不同种植年限样地较为齐全,林相保存较为完整,中间锦鸡儿样地带间距6~7 m,株行距均为1 m,按两行一带配置。通过走访和查阅种植方案的方式,分别选择了1975年、1987年、1996年和2005年(46 a、34 a、25 a、16 a)4个不同种植年限的中间锦鸡儿人工林样地为研究对象,初步勘查结果显示,各样地内土壤均为风沙土,平地无坡度,生境与原始生境的一致性较高,同时,相关部门于2011年开始对该区域进行了围封禁牧,确保了样地不受外界干扰。2021年8月,在每个样地中选择10株长势一致的植株,随机采集200片完全伸展且无明显损伤的成熟叶片,将叶片按植株做好标记,放入泡沫箱中迅速带回实验室进行植物形态指标测定,每个指标重复5次。另选取部分叶片放置于FAA固定液中进行叶片结构性状指标的测定,每个指标重复5次。
-
叶片形态性状主要包括叶长(LL,cm)、叶宽(LW,cm)、叶面积(LA,cm2)、叶片鲜质量(LFW,g)、叶片干质量(LDW,g)、比叶面积(SLA,cm2·g−1)、叶片干物质含量(LDMC,g·g−1)。采用扫描仪(Epson, Expression 12000XL)对叶片进行扫描,再通过Photoshop(Adobe Photoshop CC 2019)计算叶长(LL)、叶宽(LW)、叶面积(LA)。利用电子天平(精度 ± 0.000 1)测量叶片鲜质量(LFW)后,将叶片置于105 ℃烘箱杀青30 min,再转为80 ℃烘干至恒质量,测定叶片干质量(LDW),计算叶片干物质含量(LDMC)、比叶面积(SLA )。计算公式如下:
叶片结构性状主要包括叶片厚度(LT,mm)、栏海比(PT/ST)、主脉维管组织宽度(VB,mm)、叶片上层表皮毛密度(UEH,个·mm−2)、下层表皮毛密度(LEH,个·mm−2)、上层表皮气孔密度(US,个·mm−2)、下层表皮气孔密度(LS,个·mm−2)、上层表皮气孔面积(USA,μm2)、下层表皮气孔面积(LSA,μm2)。将采回叶片切成1 cm × 1 cm大小,加入适量H2O2∶95%乙醇(2∶1(v∶v))溶液,将试管放置于水浴锅中沸水浴至叶片完全变成白色,再将其取出放置于载玻片上,用软毛刷去除多余叶肉组织,保留叶表皮组织后进行制片,后置于Olympus DP74显微镜下进行观察,采用软件cellsens 3.2进行拍照,并计算叶片上层表皮毛密度(UEH)、下层表皮毛密度(LEH)、上层表皮气孔密度(US)、下层表皮气孔密度(LS)、上层表皮气孔面积(USA)、下层表皮气孔面积(LSA),计算公式如下:
石蜡切片观察时,叶片组织首先用乙醇梯度脱水,然后用二甲苯和石蜡进行组织透明,将透明组织嵌入石蜡(60 ℃)中进行切片、着色并在Olympus DP74显微镜下进行观察,同时,采用软件cellsens 3.2 对叶片厚度(LT)、栏海比(PT/ST)、主脉维管组织宽度(VB)进行拍照、测量与计算,计算公式如下:
-
降水数据来源于中国气象数据网(
http://data.cma.cn/ ),调查范围为盐池县2005—2021年4—7月降水量,数据如图1所示。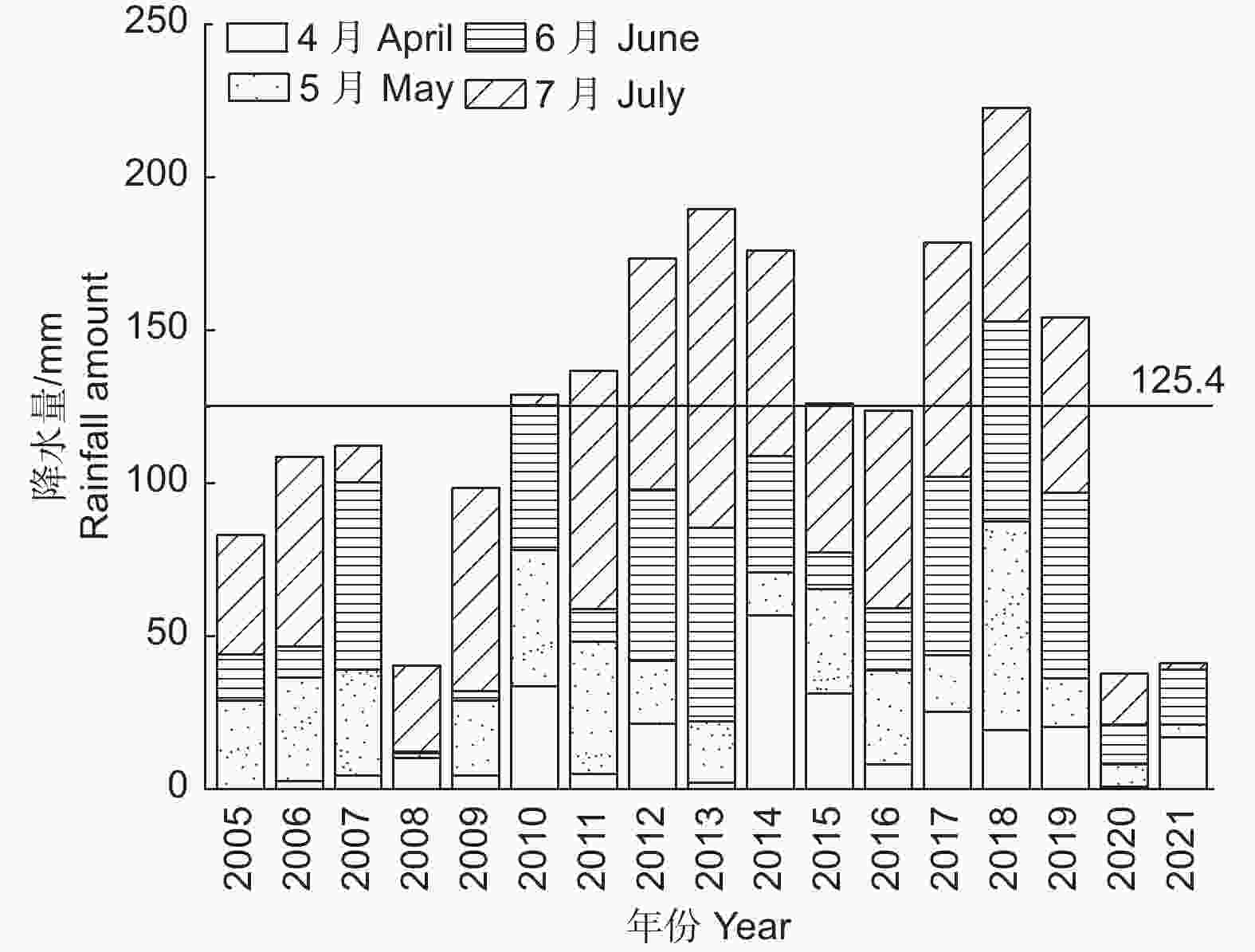
Figure 1. Trends of annual rainfall in Yanchi City from 2005 to 2021
-
利用Excel 2010对数据进行初步整理,用SPSS 26.0进行数据分析。采用变异系数计算各叶片功能性状的变异程度;用Pearson相关系数检验叶片功能性状之间的相关性;单因素方差分析和邓肯法对不同种植年限中间锦鸡儿叶片功能性状的平均值进行显著性检验;用Origin 2021对叶片功能性状进行主成分分析并可视化分析结果。
-
由表1可知,25 a中间锦鸡儿LL显著高于16、34、46 a(p<0.05),34 a中间锦鸡儿LL显著高于16、46 a,而16 a与46 a中间锦鸡儿LL并无显著差异(P<0.05)。25 a和34 a中间锦鸡儿LW显著高于16 a与46 a(P<0.05)。25 a中间锦鸡儿LFW、LDW显著高于16 a与46 a,与34 a中间锦鸡儿LFW、LDW并无显著差异(P<0.05)。25 a中间锦鸡儿LA显著高于16、34、46 a(P<0.05)。25 a中间锦鸡儿的SLA显著高于16、34 a,与46 a之间并无显著差异(P<0.05)。46 a中间锦鸡儿LDMC显著高于16、25、34 a,但16、25、34 a之间LDMC并无显著差异(P<0.05)。随着种植年限的增加,LL、LW、LFW、LDW、LA均呈现出先升高后降低的趋势,整体表现出25 a>34 a>16 a>46 a的生长趋势。
叶片形态性状
Leaf morphological traits种植年限 Planting years/a 16 25 34 46 叶长 LL/cm 0.89 ± 0.18 c(20.64%) 1.36 ± 0.19 a(13.64%) 1.08 ± 0.14 b(13.32%) 0.84 ± 0.15 c(17.64%) 叶宽 LW/cm 0.35 ± 0.08 b(23.40%) 0.52 ± 0.11 a(21.47%) 0.45 ± 0.08 a(18.59%) 0.30 ± 0.03 b(8.75%) 鲜质量 LFW/g 2.39 ± 0.95 b(39.63%) 3.78 ± 1.67 a(44.27%) 2.92 ± 0.94 ab(32.05%) 1.13 ± 0.33 c(29.44%) 干质量 LDW/g 1.13 ± 0.44 c(38.73%) 1.75 ± 0.82 a(46.98%) 1.48 ± 0.42 ab(28.59%) 0.71 ± 0.21 c(29.53%) 叶面积 LA/cm2 51.74 ± 19.70 c(38.09%) 108.10 ± 31.95 a(29.56%) 72.93 ± 18.11 b(24.83%) 39.78 ± 8.56 c(21.53%) 比叶面积 SLA/(cm2·g−1) 50.25 ± 19.90 b(39.61%) 66.50 ± 12.63 a(18.99%) 50.24 ± 7.03 b(14.00%) 59.37 ± 15.95 ab(26.87%) 叶片干物质含量 LDMC/(g·g−1) 0.48 ± 0.04 b(7.57%) 0.46 ± 0.05 b(10.79%) 0.51 ± 0.04 b(7.85%) 0.63 ± 0.10 a(15.76%) 注:数据为均值 ± 标准误(变异系数),每组数据重复5次;不同小写字母代表不同种植年间性状差异显著性(P<0.05)。
Note: The data are mean ± standard error(coefficient of variation), repeat each data set 5 times; different lowercase letters indicate significant differences among different planting years(P<0.05).LL,leaf length.LW,leaf width.LFW,leaf fresh weight.LDW,leaf dry weight.LA,leaf area.SLA,specific leaf area.LDMC,leaf dry matter content.Table 1. Morphological traits of Caragana intermedia leaves in different planting years
不同种植年限中间锦鸡儿叶片形态性状存在一定程度的变异特征。LFW、LDW和LA的变异系数明显高于其他叶片性状。16 a中间锦鸡儿叶片形态的总功能性状变异系数明显大于25、34、46 a,其中16 a中间锦鸡儿的LFW、LDW、LA和SLA为强变异(38.09%~39.63%),25 a中间锦鸡儿的LFW、LDW为强变异(44.27%~46.98%),LL、LW、LDMC均为弱变异或中等变异。
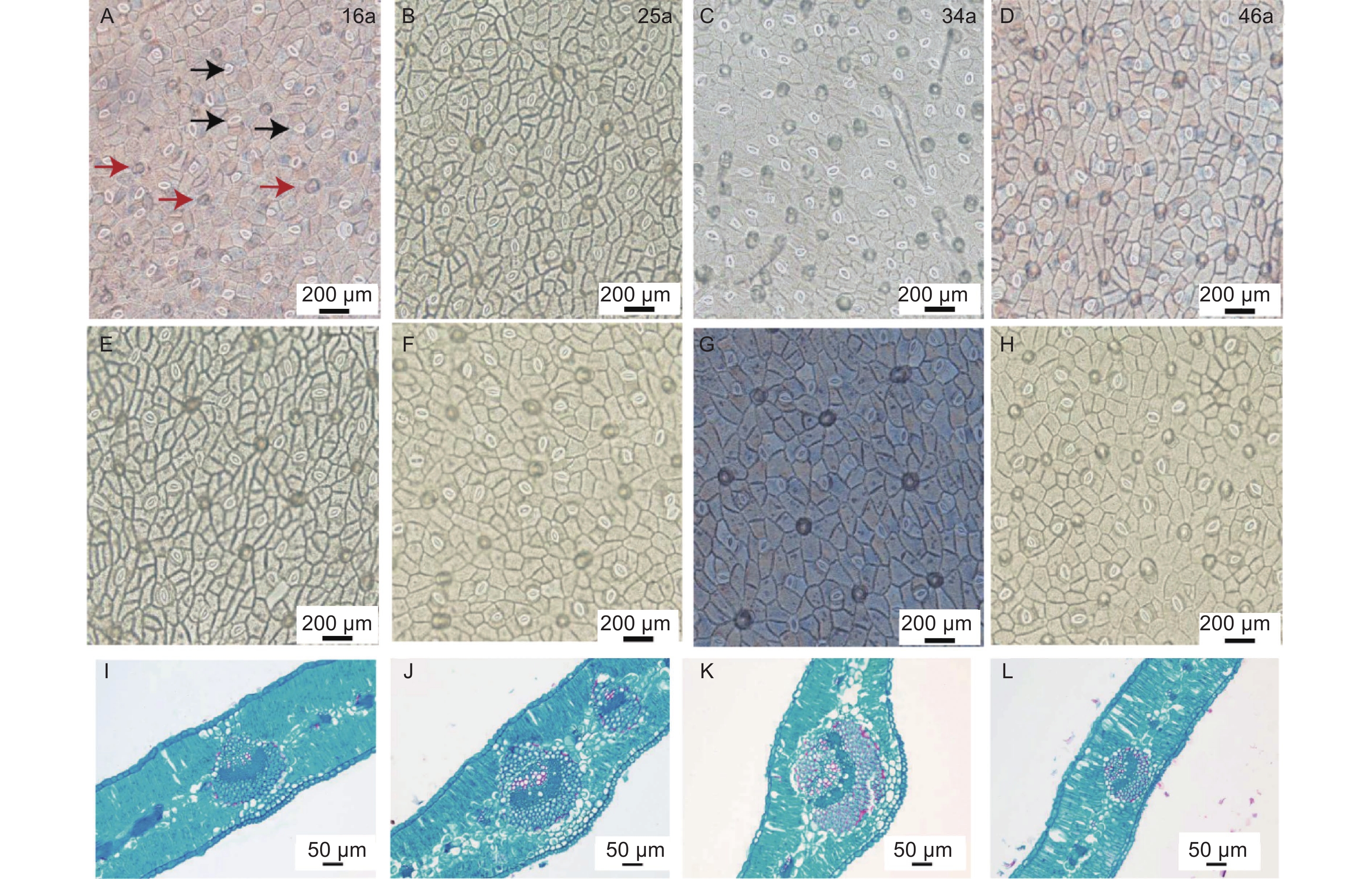
由表2和图2可知,LT、PT/ST、UEH、LEH、US、LS、LSA之间均无显著差异(P<0.05)。34 a中间锦鸡儿VB显著高于46 a(P<0.05),而与16、25 a之间无显著差异(P<0.05)。34 a中间锦鸡儿叶片的USA显著高于16 a与46 a,而与25 a之间并无显著差异(P<0.05)。
叶片结构性状
Leaf structural traits种植年限 Planting years/a 16 25 34 46 叶片厚度LT/mm 0.21 ± 0.03(14.58%) 0.22 ± 0.03(13.34%) 0.20 ± 0.02(11.73%) 0.22 ± 0.03(13.34%) 栏海比PT/ST 2.98 ± 0.80(29.97%) 2.02 ± 0.28(13.77%) 2.07 ± 0.61(26.31%) 2.84 ± 1.03(36.35%) 主脉维管组织宽VB/mm 0.16 ± 0.05ab(29.75%) 0.18 ± 0.03ab(15.66%) 0.21 ± 0.02a(7.53%) 0.16 ± 0.03b(19.01%) 上层表皮毛密度UEH/(个·mm−2) 270.80 ± 38.62(14.26%) 302.00 ± 73.83(24.45%) 280.00 ± 26.32(9.40%) 302.00 ± 39.54(13.09%) 下层表皮毛密度LEH/(个·mm−2) 232.60 ± 26.35(11.33%) 242.60 ± 78.89(32.52%) 175.60 ± 63.11(35.94%) 257.60 ± 58.26(22.62%) 上层表皮气孔密US/(个·mm−2) 161.00 ± 124.65(77.42%) 135.80 ± 49.19(36.22%) 164.60 ± 44.94(27.30%) 203.40 ± 109.29(53.73%) 下层表皮气孔密LS/(个·mm−2) 174.60 ± 137.95(79.01%) 116.00 ± 27.71(23.89%) 115.60 ± 60.26(52.13%) 176.00 ± 96.01(54.55%) 上层表皮气孔面积USA/μm2 172.93 ± 23.78b(13.75%) 183.31 ± 10.77ab(5.87%) 211.35 ± 23.01a(10.89%) 163.23 ± 30.34b(18.59%) 下层表皮气孔面积LSA/μm2 201.46 ± 29.05(14.42%) 212.39 ± 23.11(10.88%) 231.91 ± 51.23(22.09%) 207.77 ± 28.80(13.86%) 注:数据为均值 ± 标准误(变异系数),每组数据重复5次;不同小写字母代表不同种植年间性状差异显著性(P<0.05)。 Note: The data are mean ± standard error (coefficient of variation) , repeat each data set 5 times; different lowercase letters indicate significant differences among different planting years(P<0.05).LT,leaf thickness.PT/ST,palisade tissue/sponge tissue.VB,wide of vascular bundles.UEH,upper epidermal hair.LEH,lower epidermal hair.US,upper stomatal.LS,lower stomatal.USA,upper stomatal area.LSA,lower stomatal area. Table 2. Structural traits of Caragana intermedia leaves in different planting years
不同种植年限中间锦鸡儿叶片结构性状存在一定程度的变异特征。US和LS的变异系数显著高于其他叶片性状。16 a中间锦鸡儿各项叶片结构功能性状变异系数明显大于25、34、46 a,其中16 a和46 a中间锦鸡儿叶片的US、LS为强变异,分别为77.42%~79.01%和53.73%~54.55%,25 a中间锦鸡儿叶片的US为强变异(36.22%),34 a的LEH、LS为强变异(35.94%、52.13%),其余LT、PT/ST、VB、UEH、USA和LSA均为弱变异或中等变异。
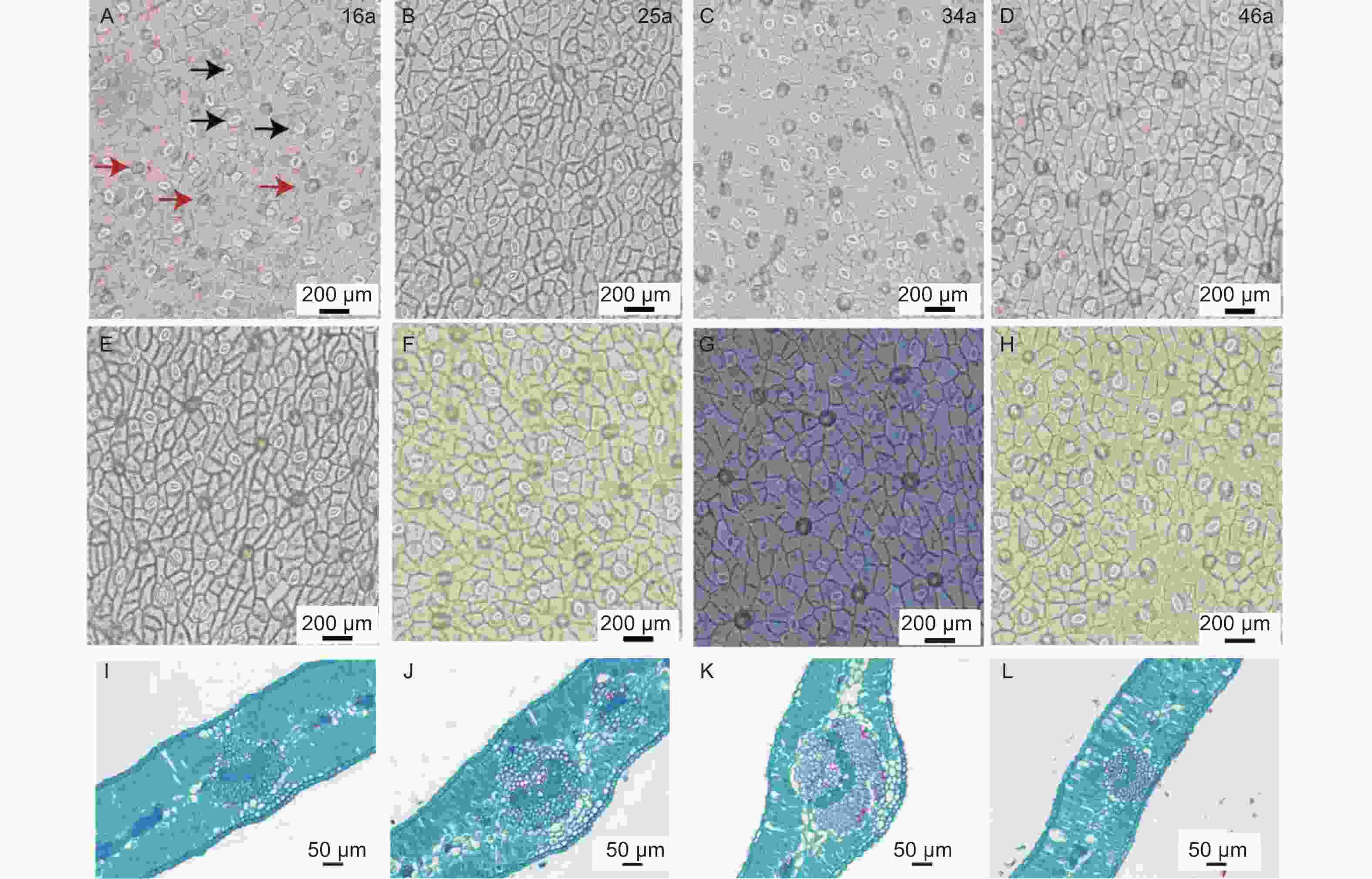
Figure 2. Stomatal anatomy and paraffin sections of Caragana intermedia in different planting years
-
叶片形态性状间存在显著的相关性,且各形态性状之间联系紧密。由图3可知,LL、LW、LFW、LDW、LA两两间呈极显著正相关(P<0.01); SLA与LL呈显著正相关(P<0.05), SLA与LDW之间呈显著负相关(P<0.05),与其他性状之间不存在显著相关关系;LDMC与LL、LFW、LA呈极显著负相关(P<0.01),与LW、LDW呈显著负相关(P<0.05),与SLA之间不存在显著相关关系。

Figure 3. Correlation between leaf morphological and structural traits of Caragana intermedia in different planting years
叶片结构性状间存在显著的相关性,但各结构性状之间联系不紧密。由图3 可知,PT/ST与UEH呈显著负相关(P<0.05);UEH与LEH呈极显著正相关(P<0.01),与US显著正相关(P<0.05);US与LS呈极显著正相关(P<0.01)。LEH与LSA呈极显著负相关(P<0.01)。
-
在植物叶片功能性状的主成分分析中(表3、4),不同种植年限中间锦鸡儿叶片形态性状指标与结构性状指标存在不同的表现,由表3可知,叶片形态形状的PCA分析得到3个主成分特征值大于1,累计方差贡献率达95.752%,说明这3个主成分能够反映7个指标的绝大部分信息。PC1贡献率为64.55%,贡献较大的为LL、LW、LFW和LDW,PC2贡献率为20.341%,贡献较大的为SLA,PC3贡献率为10.861%,贡献较大的为LDMC。
指标
Indices主成分1
Principal component 1主成分2
Principal component 2主成分3
Principal component 3叶长 LL 4.966 0.683 0.228 叶宽 LW 4.998 0.276 0.567 鲜质量 LFW 5.291 0.400 0.491 干质量 LDW 5.095 −0.812 −0.250 叶面积 LA 4.923 −1.050 0.193 比叶面积 SLA 0.268 2.431 0.308 叶片干物质含量 LDMC −2.977 −0.592 2.124 特征值 Eigenvalue 4.519 1.424 0.760 贡献率 Contribution rate/% 64.550 20.341 10.861 累计贡献率 Accumulative contribution rate/% 64.550 84.891 95.752 Table 3. Leaf morphological traits principal component factor loading, coefficient eigenvalues and variance contribution rates
由表4可知,叶片结构形状的PCA分析得到3个主成分特征值大于1,累计方差贡献率达72.488%,说明3个主成分能够反映9个指标的绝大部分信息。PC1贡献率为29.669%,贡献较大的为LEH,PC2贡献率为25.712%,贡献较大的为US和LS,PC3贡献率为17.107%,贡献较大的为UEH。
指标
Indices主成分1
Principal component 1主成分2
Principal component 2主成分3
Principal component 3叶片厚度 LT 0.657 −3.615 0.904 栏海比 PT/ST 1.099 −1.664 −3.273 主脉维管组织宽 VB −2.967 −0.266 2.162 上层表皮毛密度 UEH 2.351 1.376 2.885 下层表皮毛密度 LEH 3.799 −0.719 1.386 上层表皮气孔密度 US 1.952 3.837 −0.047 下层表皮气孔密度 LS 2.223 3.088 −1.700 上层表皮气孔面积 USA −3.244 2.501 0.702 下层表皮气孔面积 LSA −3.612 0.921 −0.975 特征值 Eigenvalue 2.670 2.314 1.540 贡献率 Contribution rate/% 29.669 25.712 17.107 累计贡献率 Accumulative contribution rate/% 29.669 55.381 72.488 Table 4. Leaf structure traits principal component factor loading, coefficient eigenvalues and variance contribution rates
-
植物功能性状是植物适应环境的外在表达,反映了植物在不同环境中的生存策略。探究干旱区荒漠植物功能性状及其与环境之间的关系,有助于理解植物适应极端环境的生态对策,可为保护荒漠地区生态系统提供理论依据[29]。本研究对4个不同种植年限中间锦鸡儿人工林调查发现,为适应极端干旱气候,各种植年限间,中间锦鸡儿叶片形态性状表现出显著的种内差异,变异系数在7.57%~46.98%之间,其中,LFW、LDW、LA、SLA对极端干旱气候更为敏感。已有研究表明,LDMC与叶片平均组织密度、植株潜在相对生长率以及叶片寿命显著相关[20,30]。本研究发现LDMC随着种植年限的增加,表现出先降低后增加的趋势,46 a中间锦鸡儿的LDMC显著高于其他3个种植年限,为(0.63 ± 0.10)g·g−1。同时,SLA和LDMC反映了植物性状中生物量的快速生产(高SLA,低LDMC)和养分的有效保存(低SLA,高LDMC)[31]。因此,本研究认为,25 a中间锦鸡儿表现出生物量的快速生产,而34 a和46 a表现出养分的有效保存。此外,已有研究认为,长期处于干旱环境的植物的LL出现缩短 [32],这是提高养分利用效率的一种生存策略[33]。而在本研究中,16 a和46 a中间锦鸡儿的LL显著低于25 a和34 a,这可能是16 a中间锦鸡儿叶片处于生长初期,自身叶片构建尚未完成,而46 a中间锦鸡儿LL随种植年限的增加而降低,以便更加有效地利用和储存养分。综合以上研究表明,在极端干旱条件下,46 a中间锦鸡儿能够通过调整资源存储与利用的生长策略,表现出对气候变化的高度适应能力[34]。
叶片解剖结构变化在植物应对干旱胁迫过程中起着关键作用,能够反映植物生长特性及其对环境的适应性[35]。本研究发现,极端干旱条件下不同种植年限间中间锦鸡儿叶片结构性状变异系数在5.87%~79.01%之间,其中,US、LS的变异程度较大,而LT、UEH、USA、LSA变异程度较低,表明US和LS对极端干旱气候较为敏感[36]。有研究表明,干旱条件下植物气孔密度受到叶片面积和气孔大小的共同影响[37],而气孔密度与面积的减少可直接导致植物生物量降低[38]。在本研究中,尽管46 a中间锦鸡儿的气孔密度最高,但是46 a中间锦鸡儿的气孔面积最小,说明在极端干旱发生时,46 a中间锦鸡儿通过降低气孔面积,增大气孔密度来阻止水分过度丧失,进而限制了植物获取资源的能力[39-40]。 16、46 a中间锦鸡儿PT/ST和VB变异程度均大于25、34 a,表明16、46 a中间锦鸡儿叶片结构对环境具有较强的潜在适应能力。高度发达的PT可避免强烈光照对叶肉细胞的灼伤[41],发达的VB有利于水分迅速运输, 加速光合产物的运输与形成, 以维持光合作用的继续进行[42], 是对干旱环境的一种结构性适应,有助于植物在干旱环境下正常生长。
植物通常通过多个性状协同变化的方式来响应环境变化,不同生长类型的植物会根据环境的变化特征制定不同的生存策略 [43-44]。本研究发现,LL、LW、LFW、LDW是中间锦鸡儿随着种植年限的增加,逐步响应干旱胁迫的主要形态性状指标,UEH、LEH、US与LS可作为中间锦鸡儿逐步响应干旱胁迫的主要结构性状指标,UEH和LS的整体变异显著高于其他叶片性状,16 a中间锦鸡儿叶片结构的总功能性状变异系数明显大于25、34、46 a。说明,中间锦鸡儿在长期生长发育过程中,根据当地环境变化,各个叶片性状之间相互组合,共同协调,已形成了一种适应于自身生长的生态策略。
综合以上结果,本研究认为,在极端干旱条件下,中间锦鸡儿人工林随着种植年限的增加,形成了从生物量快速生产到养分有效保存策略,这也是其各个叶片功能性状间相互调节与权衡的结果,以便其能够更好地适应干旱区资源、环境贫瘠的状况[45-46]。
-
(1)随种植年限的增加,46 a中间锦鸡儿LDMC逐渐增加,表明其用于叶片结构建设的碳投资较高,当极端干旱发生时,46 a中间锦鸡儿只能通过降低叶片气孔面积,增大气孔密度来阻止水分过度丧失,从而度过极度干旱时期;16 a可能是在未受干旱胁迫时优先关顾生长,叶片干物质含量储存较少,一旦严重干旱导致气孔关闭,缺少足够的资源用于叶片的发育和生长;相对而言,25 a和34 a较好的平衡了生长和储备,在极度干旱时仍然有较充足的物质以供叶片生长。
(2)在极端干旱发生时,气孔作为主要调节叶片性状的指标,通过调节叶片气孔密度以及大小之间的关系来应对环境变化,其中16 a和46 a的叶片受干旱胁迫最为严重,导致其变异系数最大。
Differences in Leaf Functional Traits of Caragana intermedia Planted in Different Years
- Received Date: 2023-03-03
- Accepted Date: 2023-04-03
- Available Online: 2023-08-20
Abstract:

 DownLoad:
DownLoad: