




-
植物开花是高等植物由营养生长到生殖生长的重要节点[1],竹类植物开花现象十分罕见,自然条件下竹类植物要经历几十年甚至上百年的营养生长阶段才会开花,且不同竹种的开花周期有所不同,如紫竹(Phyllostachys nigra (Lodd. ex Lindl.) Munro)、冷箭竹(Bashania fangiana (A. Camus) Keng f. et Wen)、毛竹(Ph.edulis (Carrière) J. Houz.)开花周期约为50 a,而角竹(Ph. fimbriligula T. W. Wen)开花间隔期至少在60 a以上[2],但由于地域性影响和环境变化会对竹子开花周期的考察记录产生局限性,因此前期记载的竹类植物开花周期数据可能存在误差。竹子开花现象常常伴随着竹子的死亡,且大多数竹类植物的结实率低,尤其是一些栽培种[3]。竹类植物开花的这些特点严重阻碍了科研工作者对竹类植物的生殖生物学和胚胎学等的探索和研究。近年来随着开花竹种逐渐增加,国内对竹子花器官的形态与解剖结构相关的研究报道也随之增加,主要集中在竹子花器官及雌雄配子发育的研究,如雷竹(Ph. Praecox C. D, Chu et C. S. Chao cv. Prevernalis)[4]、巨龙竹(Dendrocalamus sinicus Chia et J. L. Sun)[5]、鹅毛竹(Shibataea chinensis Nakai)[6]、孝顺竹(B. multiplex (Lour.) Raeusch. ex Schult. & Schult. f.)[7]、青丝黄竹(B. eutuldoides McClure var. basistriata McClure)[8]、绵竹(B. intermedia Hsueh et Yi)[9]、新小竹(Neomicrocalamus prainii (Gamble) P. C. Keng)[10]、‘霞早’绿竹 (B. oldhami ‘Xia Zao’ ZSX)[11] 、硬头黄竹(B. rigida Keng & P. C. Keng)[12]等竹种。
青皮竹(Bambusa textilis McClure)属于禾本科(Gramineae)竹亚科(Bambusoideae)箣竹属(Bambusa)竹种,产于广东、广西等地,江浙地区、福建、湖南、贵州、云南都有引种栽培,秆为优良的竹编用材和造纸用材,是广东省造纸厂的主要原料[13]。目前国内对青皮竹的变种及变型、竹林培育、病虫害防治及材性研究[14],尤其是力学性能方面[15]等方面进行了相关研究报道,国外关于青皮竹的研究大多与其竹叶提取物相关[16],但有关青皮竹花器官的研究未见,缺乏对自然生长条件下的青皮竹花器官具体的形态观察描述及胚胎发育、败育方面的系统性研究。本研究通过解剖及石蜡切片等手段,对青皮竹花器官进行形态与解剖观察,系统的描述青皮竹花器官的形态与解剖特征,分析青皮竹花器官雌、雄配子体发育过程,进而探讨青皮竹自然状态下结实率低的原因,并可为竹类植物分类学及生殖生物学研究积累资料。
-
实验材料于2013年3月采集自云南省昆明市西南林业大学珍稀竹种园(25°03′ N,102°45′ E),仅发现一丛青皮竹竹丛开花,其他竹丛均未开花。由于该竹丛引种历时久远,未查阅到相关引种记录。
西南林业大学竹园面积1 900 m2,红土,偏酸性。该地区海拔1 891 m,属于北纬低纬度亚热带高原山地季风气候,季节差异不明显但分旱、雨两季,5月—10月雨水较多为雨季,11月—来年4月降水稀少为旱季,年平均降水量1 450 mm,偶尔受北方冷空气影响出现降雪天气。全年温差较小,年均温14.7 ℃,最冷月气温7.7 ℃,年积温(≥10 ℃)4 480 ℃,日最低气温≤0 ℃日数18 d,极端低温−5.4 ℃,年平均有霜日数80 d。
-
选取并采集各个发育阶段的青皮竹小花,将花枝及小穗剪下快速浸泡于FAA固定液(50%乙醇∶甲醛∶冰醋酸=18∶1∶1)中,带回实验室并真空抽气保存。
对固定保存后的小穗及小花进行解剖和石蜡切片。使用体式解剖镜(Olympus HO11)对花器官进行解剖,按照顺序剥离小穗上的小花,并对各个小花的每个部位进行拍照、测量。将剥离出来的不同发育阶段的子房和花药进行逐级脱水、浸蜡包埋、染色及石蜡切片[17],切片厚度约为7 μm,番红-固绿双重染色,中性树胶封片,在光学显微镜下观察并使用二维测量软件 DS-3000拍照。
-
青皮竹的每个花枝各节约有3~5个小穗(图1A),青皮竹小穗形状饱满,呈淡黄色或黄色,小穗基部常包裹着2~3枚黄棕色苞片,近似卵形,无毛,苞片一般不脱落,附着在小穗基部(图2B)。小穗基部具有潜伏芽,可通过产生新的花芽生长成新的小穗,而老的小穗逐渐脱落,这具有有限花序的特征(图2A)。小穗轴节间清晰可见,但小穗柄不发达。青皮竹每个小穗具有8~10朵小花,基部小花较大,长约1.80~2.00 cm,靠近顶端小花逐渐减小,长约0.90~1.20 cm(图1B)。靠近小穗顶部的小花往往发育不正常,而靠近小穗基部的花先发育,同一小穗的小花由下至上的发育程度逐渐降低(图1M),具有明显的无限花序特征。通常小穗基部的小穗轴较短,一般约为0.30 cm,中部及上部的小穗轴较长,一般约为0.60 cm。小穗成熟后,相邻小花之间小穗轴易折断(图1C)。小穗轴各节顶宽底窄,宽度不相同,顶宽均长0.10 cm,底宽均长0.06 cm。小穗轴呈“喇叭状”上部中空,由上至下直径逐渐减小。表面光滑,具棱,棱间具4脉,棱上有刺毛,未成熟小花小穗轴为浅黄色或白色,成熟小花小穗轴颜色较深(图2D)。
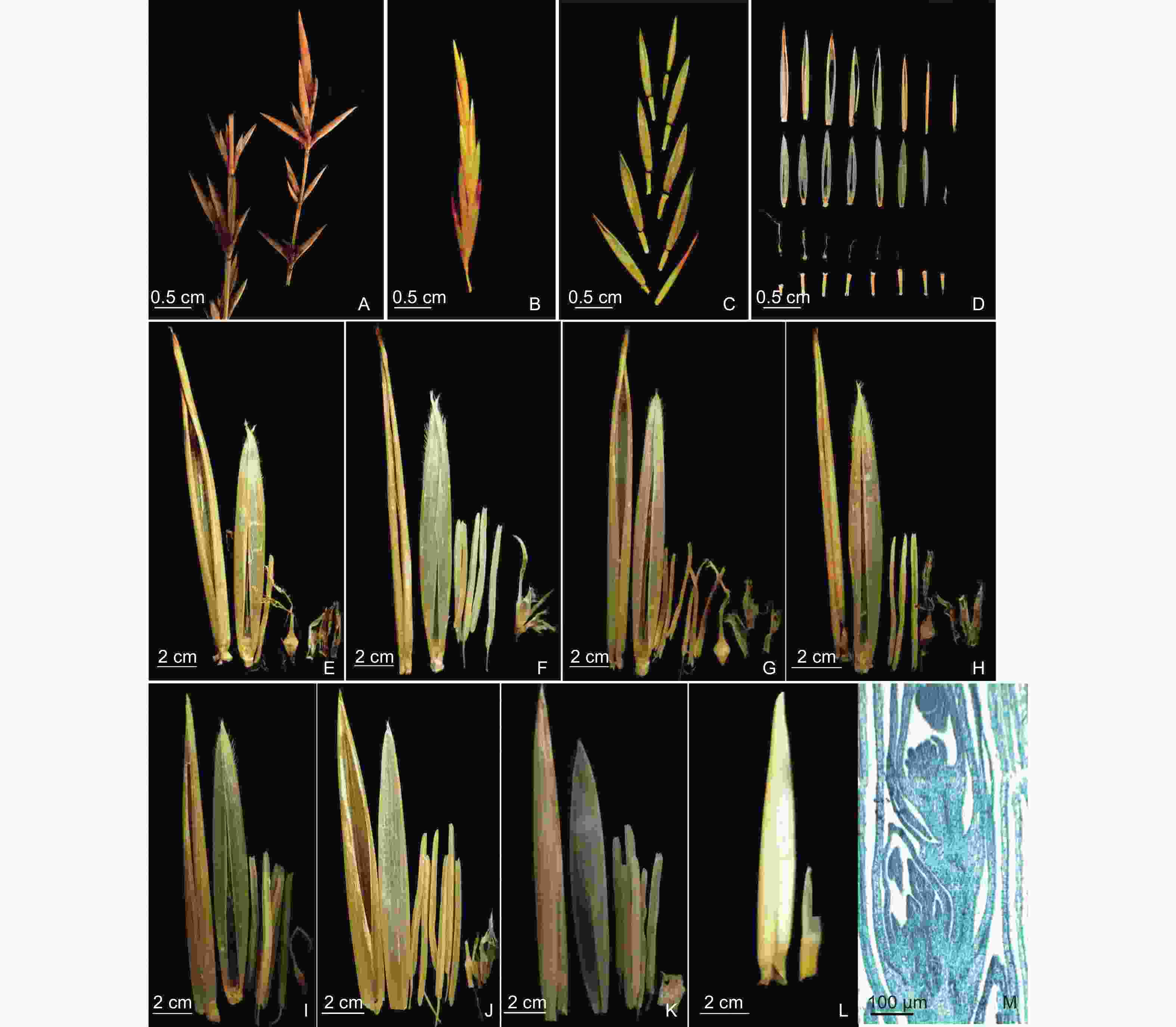
Figure 1. The structure of inflorescence and spikelet of B. textilis

Figure 2. Morphological anatomy of florets in B. textilis
-
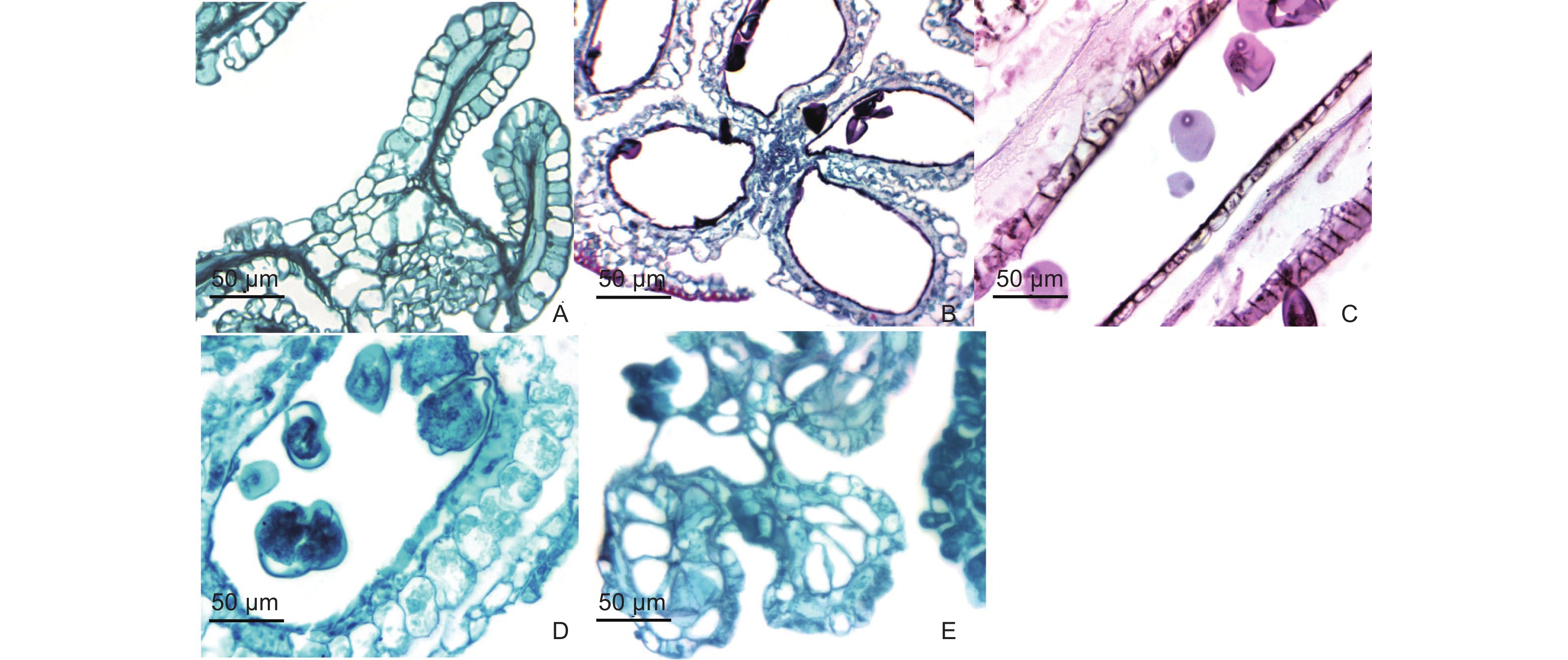
青皮竹每朵小花可分为外稃、内稃、雄蕊、浆片、雌蕊五部分(图1E~L)。青皮竹的小花含内、外稃各1枚,同一小穗下部小花的内、外稃最长,分别约为1.20 cm和1.80 cm(图1E)。越往上部,内、外稃越小,顶部小花的内、外稃最小,分别约为0.40 cm和1.00 cm(图1L)。未成熟小花的外稃长度远远大于内稃,但成熟小花内外稃长度相差较小,这与《中国竹类图志》记载的内、外稃长度相似的结论一致[18]。外稃呈椭圆形,内外表面均光滑无毛,无棱,尖端较钝,约有20脉(图2E)。内稃披针状,对生于外稃内侧,由外稃包裹,形状较扁平且质地较软,更易舒展。内稃具两脊,脊上覆有密而短的纤毛,脊间光滑,具8~10脉,脊外每边各具4脉(图2E)。内、外稃随着小花的成熟发生褐化(图2F)。浆片3枚,大小不一,长0.20~0.50 cm不等,呈透明膜质鱼鳞状,与金佛山方竹(Chimonobambusa utilis (Keng) Keng f.)[19]的肺形、斗竹(Oligostachyum spongiosum (C. D. Chu & C. S. Chao) G. H. Ye)[20]和笔竹(Pseudosasa viridula S. L. Chen & G. Y. Sheng)[21]的卵圆形不同,边缘具有较多较长的纤毛,围生于子房下部,与内稃、子房相邻(图2I)。与雷竹[22]等花相同,青皮竹小花成熟后,浆片通过吸水膨胀加厚撑开向外挤压内、外稃,露出柱头和花药,完成传粉后浆片全部褐化(图2J),所以青皮竹花也属于开放型花。雄蕊6枚,将雌蕊包围在中间(图3A)。成熟花药均长约1.20~1.50 cm。花药整体外形呈梭形,颜色为淡黄色和黄色。花药先端较尖,基部分两尖,花药四室纵裂散粉(图2G),与巨龙竹[5]、毛竹[23]、青丝黄竹[8]、新小竹[10]、鹅毛竹[6]的花药形态相似,雄蕊成熟后会发生褐化(图2H)。雌蕊1枚,未成熟时子房呈黄色,柱头为浅黄色或白色。成熟后子房下部呈乳白色,上部与花柱连接处呈黄色,柱头为黄色。成熟子房下部饱满光滑,呈花瓶状,直径约为0.20 cm。子房上部与花柱相连接处及花柱处均生长有密而短的纤毛,越靠近花柱纤毛越长。花柱长约0.10 cm,柱头为三分支羽毛状,长约0.50~0.70 cm(图2K)。子房成熟后会发生褐化(图2L)。张文燕和马乃训[24]根据稃片开合情况及柱头的长度将雌蕊分为长花柱型与短花柱型,青皮竹花的子房花柱较短,柱头远长于花柱,因此为短花柱长柱头型。青皮竹未成熟的子房上部同样具纤毛,且比成熟子房的纤毛更密更明显。青皮竹雌蕊分化出孢原细胞时,药室内的小孢子已经发育到小孢子母细胞时期,为雌雄异熟,应属异花授粉。
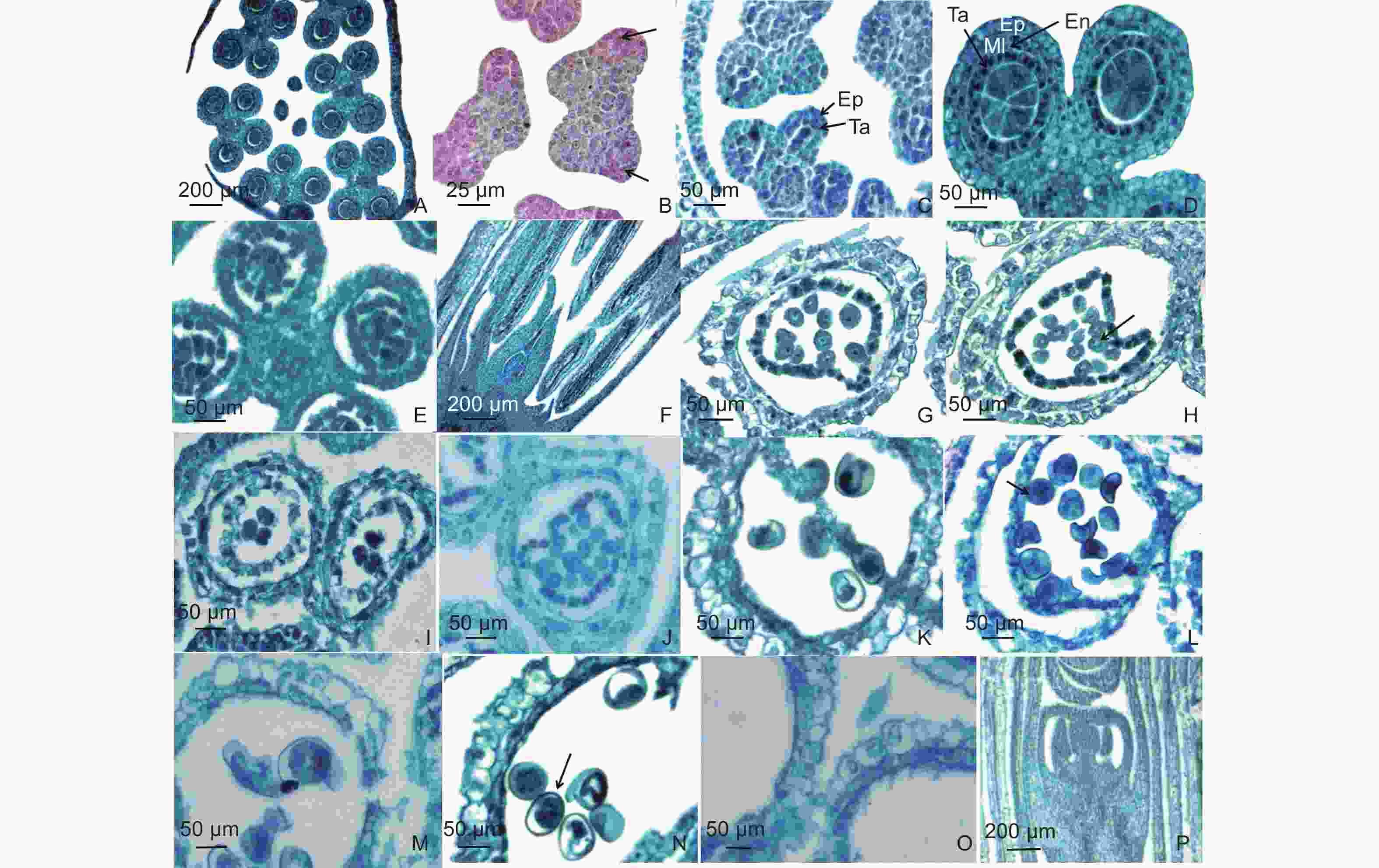
Figure 3. Anther anatomical structure of B. textilis
-
青皮竹花药属于基着药,4药室,两两对称,成熟后纵裂散粉。通过解剖青皮竹小花花药发现,花药每个药室的横切面形状大多呈圆形,少数为椭圆形(图3A)。花药壁的结构随着花药发育时期的不同而发生变化。花药分化初期,药隔组织将花药分为4个药室,每个药室角域处,表皮细胞下各含有1个孢原细胞,细胞较大,细胞质浓厚(图3B)。孢原细胞通过平周分裂形成初生壁细胞和初生造孢细胞,进入初生造孢细胞时期(图3C),此时期药室内的造孢细胞数量较少,花药壁开始出现分化。
初生造孢细胞继续分裂,进入次生造孢时期,此时期的造孢细胞数量增加,细胞质浓厚,细胞间紧密排列。同时,初生壁细胞经过平周分裂和垂周分裂形成两层次生壁细胞,外层次生壁细胞发育成为药室内壁细胞,内层次生壁细胞经过多次分裂,分化成两层细胞,分别为中层细胞和绒毡层细胞。绒毡层细胞属于腺质型,细胞质浓厚,核仁明显,花粉粒成熟后绒毡层会退化成为一薄层,紧贴药室内壁。中层细胞一般由1~2层细胞组成,后期受到另外两层壁细胞的挤压导致严重变形,最后退化消失。此时期具有明显的4层细胞结构,由内到外依次为绒毡层(Ta)、中层(Ml)、药室内壁(En)、表皮细胞(Ep)(图3D)。纵切花药发现,次生造孢时期各个药室的造孢细胞发育程度基本一致(图3F)。
次生造孢组织发生游离分散(图3E),发育成为小孢子母细胞,此时期细胞明显增大,细胞核较大,细胞质浓厚(图3G)。此时花药壁的中层细胞和部分表皮细胞仍可见细胞核。小孢子母细胞随后进行减数分裂,通过第一次减数分裂(图3H),小孢子母细胞分裂形成二分体,二分体进行第二次减数分裂形成四分体,产生的四分体呈平面左右两两对称型(图3I)。青皮竹每个花药的各个药室之间小孢子母细胞发育时期不同步,相同药室的小孢子发育时期也存在差异,因此青皮竹花药进入减数分裂时期的发育程度也并不完全不一致。减数分裂完成后,四分体的四个子细胞彼此分离形成4个单核小孢子,此时期的小孢子细胞细胞质浓厚,无液泡,核位于细胞中央,被称为单核中央期(图3J)。随着小孢子体积逐渐增大,细胞质液化,形成中央大液泡,液泡将细胞核推向细胞壁一侧,中层与绒毡层细胞逐渐退化,此时期为单核靠边期(图3K),小孢子壁处已经出现萌发孔(图3L)。随着花粉粒逐渐成熟,液泡被挤压消失,细胞质重新填满细胞,细胞核开始进行有丝分裂,形成双核(图3M)或3核花粉粒(图3N)。
在花药成熟过程中,花药壁中层细胞逐渐被挤压消失,花粉粒成熟以后,绒毡层完全退化为一层薄膜,与药室内壁紧密贴合。药室内壁的径向壁纤维增厚,纤维层逐渐生成,此时成熟的花药壁仅余纤维层和表皮细胞共2层细胞(图3O)。相邻药室的花药壁因未发生纤维化而纵向开裂,释放花粉。通过对青皮竹早期的花器官进行纵切可以发现,处于发育早期阶段的花药基部组织形态呈钝圆形,并不尖锐,与成熟的花药基部差别较大,因此花药基部是在花药发育成熟的过程中逐渐突起变尖的。通过纵切也可以发现,花药的维管束与小穗轴以及子房的维管束连接在一起(图3P)。
-
通过对青皮竹的成熟花药进行解剖和石蜡切片观察发现,青皮竹花药有较多的败育类型,这可能是导致青皮竹结实率低的主要原因。
观察到的青皮竹花药的败育类型主要有以下几种:(1)花药壁的表皮细胞发育良好,但其他几层细胞发育异常,药室完全皱缩,药室内物质成团(图4A);(2)花药室中空,无花粉粒形成(图4B);(3)小孢子发育成仅具有萌发孔但无完整细胞结构的空壳花粉粒(图4C);(4)成熟花粉粒开裂(图4D);(5)花药壁过度发育,向内侵占药室空间(图4E)。
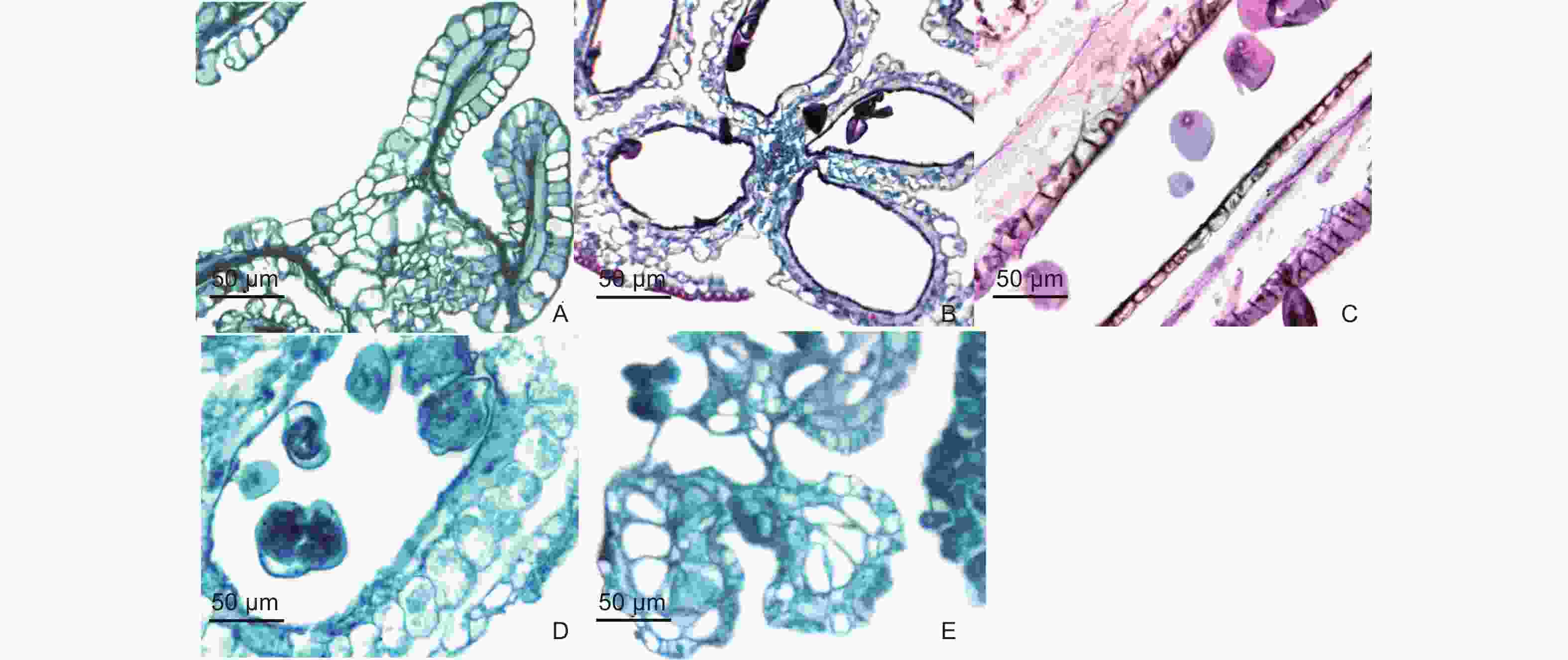
Figure 4. Abortive anthers of B.textilis
-
青皮竹子房1室,侧膜胎座,倒生胚珠,双层珠被(图5A)。孢原细胞时期子房胚珠的双珠被原基开始分化。孢原细胞位于珠孔端珠心表皮下方的第一层细胞,孢原细胞体积较大,细胞质较浓厚,细胞核较显著,核仁清晰(图5A)。孢原细胞进行分裂发育成大孢子母细胞(图5B),随后大孢子母细胞进行减数分裂,形成二分体和四分体,四分体靠近珠孔端的大孢子发育成功能大孢子,功能大孢子进行多次分裂,形成二核胚囊及四核胚囊(图5C),最终发育成具成熟胚囊的雌蕊。由于实验材料有限,观察到的子房不同发育时期较少,仅观察到孢原细胞、大孢子母细胞时期和四核胚囊时期,由于切片角度以及样品数量的问题,仅切到两核。成熟的种胚表明青皮竹的胚囊可以正常发育及受精(图5D),但在野外收集小穗时,并未采到成熟的种子。经解剖及石蜡切片纵切青皮竹成熟子房和不育小花发现,青皮竹的子房同样存在发育异常的现象,胚珠发生皱缩、退化,紧贴子房壁(图5E~F)。
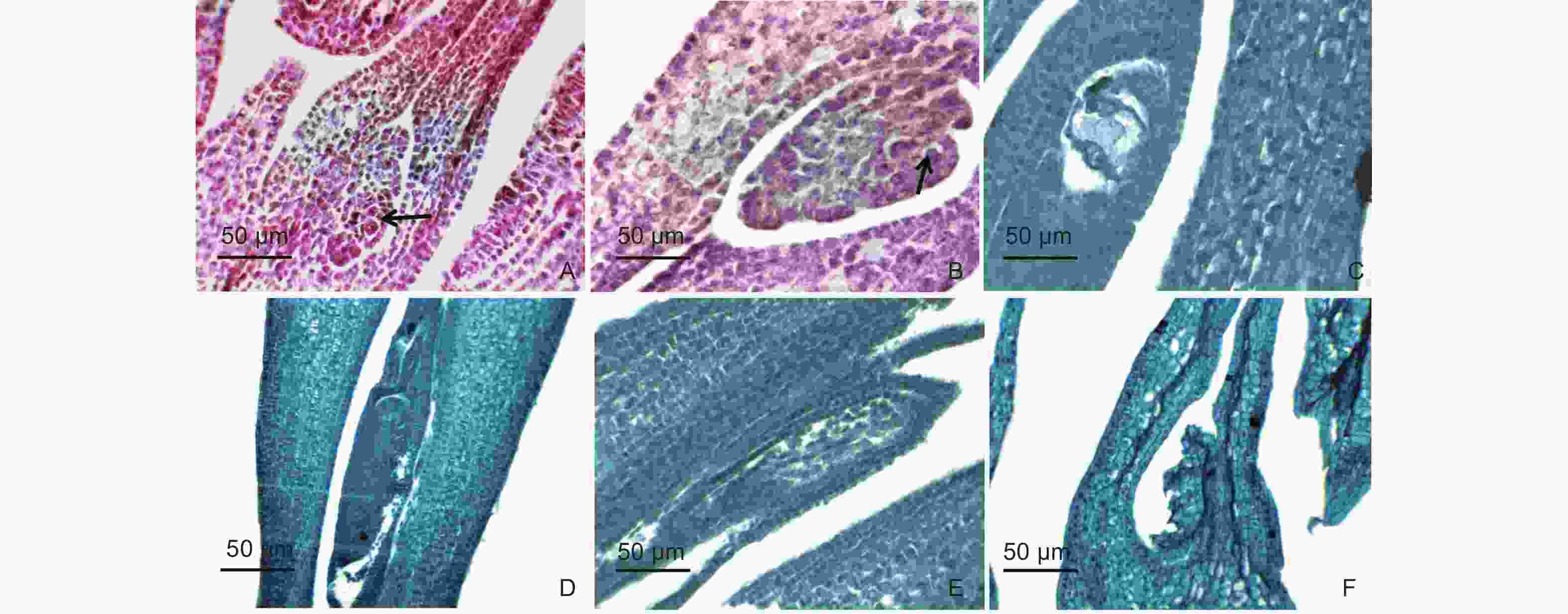
Figure 5. Ovary development and abortion of B.textilis
-
McClure[25]根据小穗基部是否具有潜伏芽将竹类植物的花序分为有限花序和无限花序两种类型,耿伯介[26]进一步定义花序为有限真花序和无限假花序,而林树燕等[27]通过对翠竹[28]、‘霞早’绿竹[11]等花序的研究,认为竹类植物的花序皆为混合花序。青皮竹小穗下部的小花先发育,具有无限花序的特征,同时小穗的基部具有潜伏芽,该潜伏芽能发育成新的小穗,并且产生新的潜伏芽,又具有有限花序的特征,因此青皮竹也应属于混合花序。
竹子的小花一般是由外稃、内稃、浆片、雄蕊和雌蕊五部分组成,它们在形态、数量和颜色上因竹种的不同而产生差异[2],青皮竹的小花同样具有这五部分。青皮竹小花包括内、外稃各1枚,外稃呈椭圆状,内稃对生于外稃内侧,较外稃短,呈披针状,这与笔竹[21]、金佛山方竹[19]内、外稃均为披针状的稃片形状略有差异。浆片3枚,呈透明膜质鱼鳞状,边缘具发达纤毛,开花时,浆片吸水膨胀,撑开内、外稃,促使雄蕊和柱头伸出小穗外,属于开放型花,与斗竹[20]、笔竹[21]和雷竹[22]等开放型花特征一致,但青皮竹小花属于雌雄异熟,异花授粉。雌蕊1枚,子房1室,侧膜胎座,倒生胚珠,双层珠被;花瓶状且光滑不具棱,上部被绒毛,短花柱长柱头,具有三分支羽毛状柱,因此与青丝黄竹的子房在形态上类似[8]。雄蕊6枚,花药4室,未成熟花药的花药壁4~5层,由内向外分别为绒毡层、中层、药室内壁和表皮;成熟后仅剩纤维层和表皮细胞两层。绒毡层为腺质型,随花药成熟发生退化,仅余一薄层紧贴花药壁。黄坚钦等[4]认为,在雷竹雄配子体发育过程中,其花药壁没有药室内壁的存在,表皮层不仅行使一般的保护功能,而且在花药成熟时有帮助花药开裂的作用,行使了药室内壁的功能。成熟花粉粒为2或3核花粉粒,这与绵竹[9]、青丝黄竹[8]报道类似。
通过切片可观察到,青皮竹的6个雄蕊原基在发育过程中存在不同步性,靠近雌蕊前端的1个花药原基首先发育,其次是雌蕊两边的4个原基,最后是雌蕊后端的1个花药原基发育,由此导致同一小花不同花药间发育程度的差异。此外,同一花药的不同药室之间也存在发育的差异性,其发育程度并不完全一致,同一花药的4个药室可能同时存在相邻的两个或三个发育时期。
竹类植物在自然条件下授粉难、开花少及结实率低等问题一直都是竹类植物胚胎学研究的重点[2],且竹类植物生殖周期长且难以预测,使研究工作更加困难。近几年国内关于竹类植物花粉败育和结实率低的报道逐渐增多,在竹类植物的有性生殖过程中,雄配子发育异常,雌配子发育正常的竹种有月月竹(Chimonobambusa sichuanensis (T. P. Yi) T. H. Wen)[29]、巨龙竹[5]、雷竹[19]、毛竹[30]、慈竹(B. emeiensis L. C. Chia & H. L. Fung)[31]等,而在孝顺竹[7]、水竹(Ph. Heteroclada Oliver)[32]中,其花药发育正常而子房发育异常。青皮竹的花药出现了多种败育情况,可能是青皮竹小花结实率低的重要原因。青皮竹雄蕊发育存在异常,从雄蕊原基开始到形成小孢子母细胞时期,细胞和花药壁均未见结构和发育异常,但从小孢子母细胞减数分裂开始到花粉粒成熟期间的各个阶段,细胞及花药壁开始出现异常,主要表现为花药的药室收缩变形至完全皱缩;花药中空,无花粉粒形成或小孢子发育成仅具有萌发孔但无完整细胞结构的空壳花粉粒;花药壁径向壁未加厚,无纤维层形成;花药壁过度发育,向内侵占药室空间。青皮竹雄蕊中花药药室皱缩和空壳花粉粒形成的现象,在棉花竹(Tongpeia fungosa (T. P. Yi) Stapleton)[33]、绵竹[9]、青秆竹(B. tuldoides Munro)[34]、龙竹(D. giganteus Wall. ex Munro)[35]等竹种中均有发现。青皮竹雌蕊发育过程中也存在败育现象,子房受精后胚珠发育异常也是青皮竹结实率低的一个原因。综上所述,在自然条件下竹类植物大多数败育花药在成熟过程中已经处于异常状态,甚至花粉成熟后无法正常释放花粉粒,这与de Souza[36]报道的花粉粒在花药成熟开裂释放后失去活性的结论不同,另外,雌蕊败育也是导致结实率低的重要原因,雌蕊和雄蕊任何一方出现异常都会导致种子无法正常形成。王曙光[5]、林树燕[37]认为除了内在原因外,外部条件如季节、气候和传粉条件等都会影响竹类植物的结实率。
-
青皮竹小花属于开放型花,具有内、外稃各1枚,浆片3枚,雌蕊1枚,雄蕊6枚。子房1室,侧膜胎座,倒生胚珠,双层珠被;花药4室,未成熟花药的花药壁4~5层。成熟花粉粒为2或3核花粉粒。青皮竹的雌蕊和雄蕊在发育过程中均出现了多种败育情况,并且青皮竹花为雌雄异熟,导致青皮竹无法自花授粉,大大降低了其开花授粉的成功率,且野外条件下竹子本身很少开花,可受粉的成熟子房较少,严重影响其结实率。本研究探讨了青皮竹的花器官形态特征,其雌、雄配子体的发育及结实率低的原因,为该属系统分类及生殖生物学提供基础资料。
Floral Morphology and Development of Female and Male Gametophytes of Bambusa textilis
- Received Date: 2023-05-24
- Accepted Date: 2023-11-13
- Available Online: 2024-02-20
Abstract:



 DownLoad:
DownLoad: