



-
赤霉素是控制植物生长并作用于植物整个生命周期的一种激素, 属于四环二萜类化合物。目前已知的赤霉素种类至少有130种, 但仅有部分赤霉素的活性较高, GA4与GA7是其中活性很高的两种。由于这两种赤霉素分子结构相似, 分离困难, 常以混合物即GA4+7的形式存在[1]。迄今为止, 有关GA4+7影响树木生长报道最多的是促进杉科(Taxodiaceae)、松科(Pinaceae)等针叶树种开花结实的研究[2-5]。如Almqvist等[6]人采用GA4+7茎干注射处理21年生的挪威云杉(Picea abies), 显著增加了雌雄花的数量; 赵鹏等[7]对15年生落叶松喷洒0.5 g·L-1的赤霉素GA4+7, 喷洒的枝条雌球花量是对照枝条的2.9倍。另外, 在果树中研究发现, GA4+7有效促进植株营养生长, 并且大幅提高开花率及座果率[8-9]。目前, GA4+7在植物生长调节和促进开花结实两方面所起的作用已被证实, 但是在国内利用GA4+7促进阔叶用材树种的生长相关报道还较少, 主要集中在施加GA3或其它易于提取的赤霉素, 研究其对植物的生长影响, 其主要原因是由于GA4+7难以获得, 且生产价格昂贵, 限制了其广泛应用[1], 所以GA4+7对于阔叶树种的处理方式、处理时间以及作用效果尚未明确。但随着GA4+7合成途径的确定, 生产菌株的获得, 使其使用成本大幅减少, 大大推动GA4+7在针阔叶树种中的应用。因此本试验以1个白桦(B. platyphylla × B. pendula)无性系为研究对象, 采用不同浓度赤霉素GA4+7处理, 在生长最旺盛的6月末取材测定基因的转录组, 目的是为了研究GA4+7对白桦生长的影响及分子机制, 也为GA4+7在白桦中的应用提供参考。
HTML
-
2013年于东北林业大学白桦强化种子园内选择1株干型挺直无病虫害的杂种白桦(B. platyphylla × B. pendula)优树的下部枝条, 带回实验室后将枝条切断, 流水冲洗2 d, 摘取枝条上腋芽置于70%乙醇中1 min, 无菌水漂洗1 min, 于超净台上剥去外层芽鳞, 0.1%(W/V)HgCl2中浸泡8 min, 10%的次氯酸钙Ca(ClO)2中10 min, 无菌水漂洗3~5次。消毒后的腋芽接种到WPM+1.0 mg·L-16-苄氨基嘌呤(6-Benzylaminopurine, 6-BA)中进行初代培养, 30 d后转接至WPM + 1.0 mg·L-16-BA +0.02mg·L-1 NAA中进行不定芽的继代培养, 2013年3月, 将组培苗进行生根培养, 5月中旬移栽至育苗盘中, 置于温室中常规管理, 6月初选取高度一致的白桦苗木, 移栽至20 cm×20 cm的花盆中, 置于东北林业大学白桦育种基地进行常规水肥管理。
-
参考Philipson的方法[10]设置0.2 g·L-1 GA4+7(T1处理)、0.4 g·L-1 GA4+7(T2处理)2种浓度, 以不喷施任何溶液为空白对照(CK), 每个处理9株, 共计27株白桦无性系。2014年4月20日至7月20日期间对参试苗木进行叶面喷施处理, 每隔15 d处理1次, 10月初待苗木封顶时调查1年生苗高、地径。2015年4月20日开始重复上述GA4+7处理, 并于2015年6月28日分别取T1、T2、CK的白桦顶端组织(包含顶芽及第一片幼叶), 迅速置于液氮中带回实验室。10月初待苗木封顶时调查2年生苗高、地径。
-
采用CTAB法提取样品总RNA后, 用DNaseⅠ(Promega, USA)消化去除DNA污染, 采用Qubit 2.0、Aglient 2100方法检测RNA样品的纯度、浓度和完整性, 从3个RNA样品中各取20 μg由百迈克生物科技有限公司进行cDNA文库的构建。分别使用Qubit2.0和Agilent 2100 Bioanalyzer对文库的浓度和插入片段大小进行检测, 使用Q-PCR方法对文库的有效浓度进行准确定量, 以保证文库质量。库检合格后, 用HiSeq2500进行测序并进行有参考基因组的转录组分析。
将测序所得Raw Data去除测序接头以及引物序列, 过滤低质量值数据, 获得高质量的Clean Data。将其与白桦基因组进行比对, 比对上的reads称为Mapped Data。通过检验插入片段在基因组上的分布, 评估mRNA片段化的随机性及mRNA的降解情况; 通过插入片段的长度分布, 评估插入片段长度的离散程度; 通过绘制饱和度图, 评估文库质量。
-
使用EBSeq进行差异表达分析, 获得2个样品之间的差异表达基因集, 采用RPKM(Reads Per Kb per Million reads)作为衡量转录本或基因表达水平的指标[11], 计算得到的基因表达量可直接用于比较不同样品间的基因表达差异。在差异表达基因检测过程中, 将Fold Change≥2且FDR(false discovery rate) < 0.01作为筛选标准定义差异表达基因。得到差异表达基因之后, 对差异表达基因做GO功能分析和KEGG Pathway分析。
RPKM计算公式为:
设RPKM(A)为geneA的表达量, 则C为唯一比对到gene A的reads数, N为唯一比对到所有gene的总reads数, L为gene A的碱基数, RPKM法能消除基因长度和测序量差异对计算基因表达的影响。
-
采用CTAB法分别提取T1、T2以及CK白桦顶端分生组织的总RNA, 经DNaseI(RNase free)消化后使用ReverTre Ace®qPCR RT Kit(Toyobo, Osaka, Japan)将提取的总RNA进行反转录。反转录产物稀释10倍, 用作定量PCR的模板, 以BpUBC和BpSAND作为内参基因, 进行实时定量PCR扩增(所有试样进行3次重复)。PCR的反应体系为: SYBR® Premix ExTaqTMⅡ(2×)10 μL, 引物各1 μL(10 μmol·L-1), 水6 μL, 模板2 μL, 反应参数为95℃预变性30 s, 95℃变性5 s, 58℃退火延伸34 s, 循环40次, 绘制溶解曲线, 温度由95℃ 15 s, 60 ℃ 1 min, 至95℃ 15 s止。以上反应在ABI PRISM® 7500荧光定量PCR仪上完成。选取未处理白桦(CK)作为对照, 用2-ΔΔCT方法[12]对定量PCR的结果进行分析, 引物序列见表 1。
基因
Gene上游序列Upstream sequence(5’-3’) 下游序列Downstream sequence(5’-3’) BpUBC GGTTTCTGGGTTACAGTAAAGTTTCAG CTTGAAGCCTGCATGTGGTC BpSAND CACAGAAGAGCCTTACGAGTCAT CCTCCAAGCAAAGATGTCATATCAAAC BP028367 GGTACTGCTGCGGGCCAAGC CAATGGCTCCTCCGCGTAGTTCTTC BP028372 GCTCATAAAGGTCCATTTACAGGCCAGGG GGATAAGGGGGCATCGAATACATATGG BP028382 CCTCGGGATCAAATTGGTTATTGGGCAG GGATTGTCCTACACTAGCACTTCCGCG BP029736 GAATCCACTGATTTCTGCCGCTTCCG GCTTCGGGCTGTCTCGCGATC BP028323 CCATTACTGATGGACAAATCTGTTCGG CTACCGCAGACTTGTTTCATAGCTTTC BP028346 CATAGATCACCCCTATTTGTGTGGTCCG CGGTTAATAACATGGTAATTGCCCCTGCC BP028374 CCGATTCTTATAGTACTGTTCGGGGAG GCTTCCGGTTTAGGTGGACAGCC BP028387 ATCCCGAAAACGCCCGGGTCG CCTCCTAGAGTAGGTATGGCATCGCC Table 1. qRT-PCR primer sequence
-
表型变异系数(PCV)采用公式PCV=δ/X×100%, δ为性状标准差, X为性状平均值[13]。
运用Microsoft Excel进行数据处理, 用SPSS v16.0软件进行方差分析及多重比较。
1.1. 材料
1.2. 方法
1.2.1. RNA的提取及cDNA文库的构建
1.2.2. 差异基因的筛选、GO、pathway分析
1.2.3. qRT-PCR验证
1.3. 遗传参数分析
-
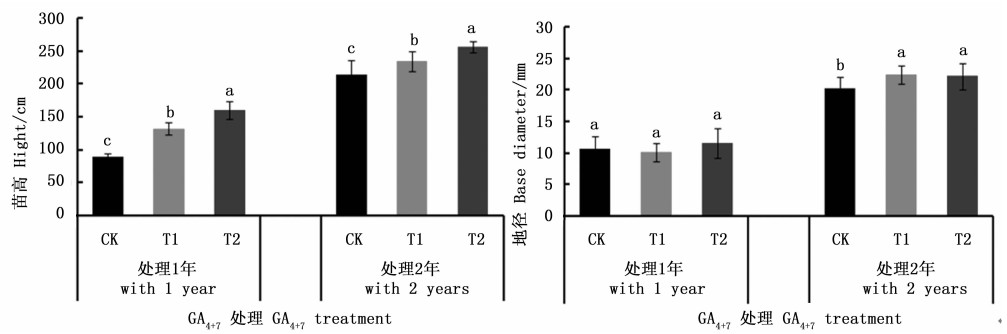
对GA4+7处理白桦无性系获得的2个年度生长性状进行方差分析以及主要遗传参数分析(表 2)。在第1个年度, 发现不同浓度的GA4+7对苗高的生长有极显著促进作用(P < 0.01), 而对地径的影响不明显(P > 0.05);在连续处理的第2个年度, GA4+7不但延续了对高生长的极显著促进作用(P < 0.01), 而且对地径生长的促进作用也达到了显著水平(0.01 < P < 0.05), 说明白桦无性系苗高对GA4+7反应较敏感, 而地径对GA4+7反应较迟钝。进一步对各性状遗传参数分析发现, 苗高与地径性状的表型变异系数均随处理时间延长而减小并且均小于25.00%, 说明无性系生长逐渐趋于稳定并且整齐度较好。
性状
Growth traits年龄
Age/a变异来源
Source of variationdf MS F P 均值
Mean标准差
Standard deviation变幅
Amplitude of variationPCV/% 苗高
Height/m1 处理间 2 11 663.59 112.63** <0.01 127.41 31.51 78.00~175.00 24.73 处理内 24 103.556 2 处理间 2 4 019.444 15.288** <0.01 235.44 23.49 180.00~269.00 9.98 处理内 24 262.907 地径
Basediameter/mm1 处理间 2 5.205 1.32 0.286 10.76 2.01 8.23~15.84 18.68 处理内 24 3.948 2 处理间 2 12.577 3.844* 0.036 21.65 2.00 17.24~25.49 9.24 处理内 24 3.272 Table 2. Analysis of variance and genetic parameters among gibberellin GA4+7 treatment birch clones on height and base diameter
对各处理的生长性状进行多重比较发现(见图 1), 白桦的苗高生长在各处理间的差异达到显著水平(0.01 < P < 0.05), 在2个年度内均表现为随着GA4+7浓度的提高其生长也加大, 其中GA4+7处理2年后, T2、T1处理的白桦无性系分别高于CK的19.65%、9.05%。在地径方面, 赤霉素GA4+7处理第1年后对苗木的地径生长影响差异不显著, 但在处理第2年后表现出明显差异, 2种浓度处理(T1, T2)苗木地径均显著高于对照(CK)苗木地径(图 1)。

Figure 1. Height and base diameter growth of gibberellin GA4+7 treatment birch clonal
-
对CK、T1、T2等白桦样品测序数据评估显示, Q30碱基百分比均不低于91.94%, 并且3个样品与基因组的比对率在79.00%以上, 说明转录组数据满足后续信息分析的需求及数据分析的可靠性。通过对插入片段在基因组上的分布及插入片段的长度分布进行检测、饱和图的绘制, 表明该转录组文库可以用于后续转录组测序。
样品
SamplesGC百分比/%
The GC percentage%≥Q30 与参考基因组的比对率/%
Mapped ratio/%CK 46.70 92.86 79.69 T1 46.72 92.40 79.87 T2 47.07 91.94 80.42 Table 3. Samples sequencing data statistics and mapped ratio
-
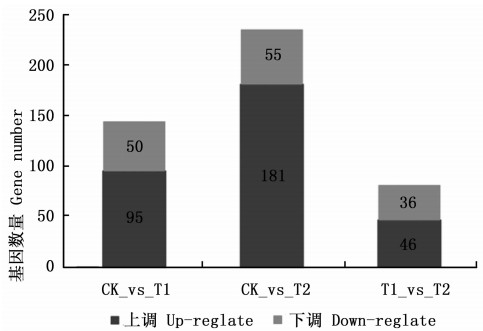
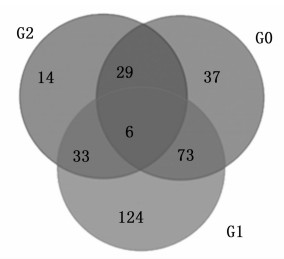
根据RPKM获得CK、T1、T2处理的白桦3个样本转录组文库的基因表达量, 比较每2样本间的差异表达基因(图 2), CK_vs_T1有145个差异基因(95个上调, 50个下调), CK_vs_T2有236个差异基因(181个上调, 55个下调), T1_vs_T2有82个差异基因(46个上调, 36个下调), 其中T1_vs_T2的差异基因最少。绘制维恩图分析3个处理差异表达基因的分布(图 3), 发现有6个基因在CK_vs_T1、CK_vs_T2、T1_vs_T2间差异表达, 有29个基因在CK_vs_T1和T1_vs_T2中差异表达, 有33个基因在CK_vs_T2和T1_vs_T2中差异表达, 有73个基因在CK_vs_T1和CK_vs_T2中差异表达。

Figure 2. The number of up-regulated and down-regulated differential expressed genes between the different treatments

Figure 3. Venn diagram of differentially expressed genes of birch under different treatments
-
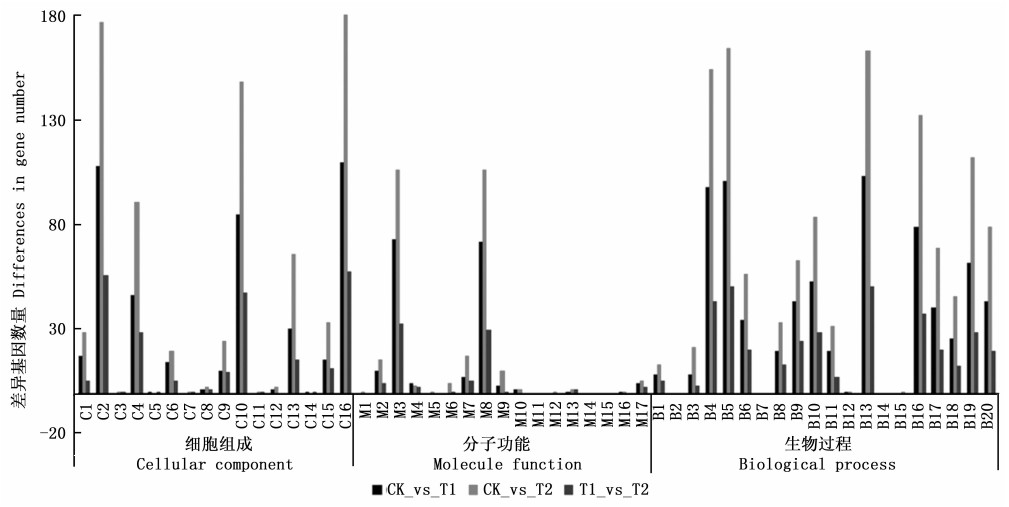
GO功能显著性富集分析能确定差异表达基因行使的主要生物学功能, 可将基因分为细胞组成、生物过程、分子功能3个大类, 因此对3个处理的差异表达基因进行了功能分类, 结果见图 4。差异表达基因主要集中在细胞组成的细胞(cell)、细胞器(organelle)、细胞部分(cell part); 分子功能的催化活性(catalytic activity)、结合(binding); 生物过程的代谢过程(metabolic process)、细胞过程(cellular process)、单一有机体过程(single-organism process)、刺激响应(response to stimulus)、生物调控(biological regulation)。由图 2可知, CK_vs_T1的差异基因数量明显少于CK_vs_T2的差异基因数量, 说明施加高浓度的GA4+7引起的差异基因数目增多。

Figure 4. GO classification of differentially expressed genes between the different birch samples
对每2个处理样品间的差异基因进行GO富集分析, 以CK_vs_T2为例(图 5), 差异基因在叶绿体类囊体膜(chloroplast thylakoid membrane)、红光的细胞响应(cellular response to red light)、远红光的细胞响应(cellular response to far red light)、蓝光的细胞响应(cellular response to blue light)等方面富集显著。

Figure 5. Analysis of CK_vs_T2 difference gene enrichment
-
Pathway显著性富集分析能确定差异表达基因参与的主要生物过程, 因此用KEGG数据库对差异基因进行Pathway富集分析, 研究富集的主要生物学过程。
-
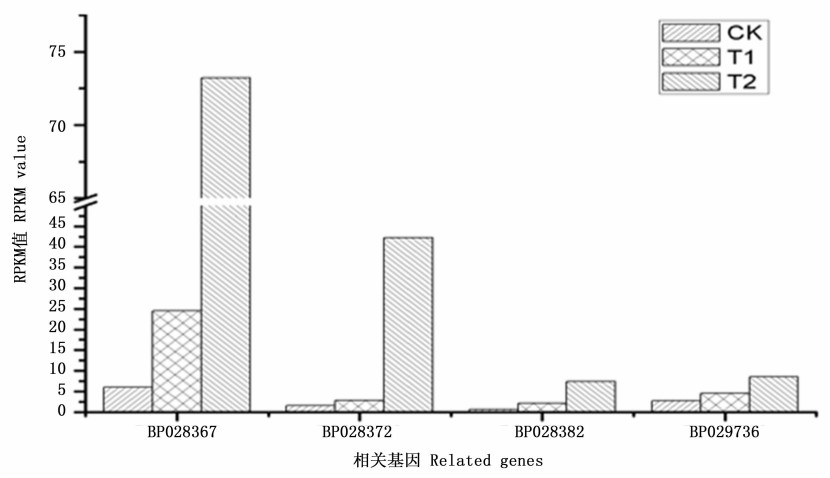
根据白桦无性系生长情况发现, GA4+7处理对白桦苗期高生长有显著促进作用, 且0.4 g·L-1 GA4+7 对白桦的促进作用较0.2 g·L-1的促进作用更显著。由于植物生物量与光合作用呈正相关关系[14], 因此对光合作用途径相关基因进行深入挖掘。Pathway富集分析结果显示: GA4+7处理白桦苗木后, CK_vs_T2的光合作用途径有4个差异基因上调表达, 基因的表达量见图 6, 这些差异基因编码的蛋白分别是光系统Ⅱ蛋白D1(photosystem II protein D1)、光系统ⅠP700叶绿素A脱辅基蛋白A2(photosystem I P700 chlorophyll A apoprotein A2)、细胞色素b6f亚组4(cytochrome b6-f complex subunit 4)、叶绿体中CF1ɑ亚组ATP合成酶(ATP synthase CF1 alpha subunit (chloroplast))。

Figure 6. Related gene expression levels among different samples in photosynthesis pathway
-
氧化磷酸化途径是有机物在分解过程中的氧化步骤所释放的能量, 驱动ATP合成的过程, 即ATP的生成过程, ATP是植物的各项生命活动直接能源物质[15]。Pathway富集结果表明: CK_vs_T2的氧化磷酸化途径有4个差异基因上调表达, 基因的表达量见表 4, 它们分别编码ATP合成酶F1亚组1(ATP synthase F1 subunit 1)、细胞色素c氧化酶亚组1(cytochrome oxidase subunit 1)、NADH-质体醌氧化还原酶亚组K(NAD(P)H-quinone oxidoreductase subunit K)、NADH锟氧化还原酶亚组2(NAD(P)H-quinone oxidoreductase subunit 2)。
基因样品
Gene samplesCK T1 T2 编码的蛋白
Encoded proteinsBP028323 0.841 1.860 5.334 ATP合成酶F1亚组1 BP028346 0.065 0.293 0.841 细胞色素c氧化酶亚组1 BP028374 4.907 11.468 15.583 NADH-质体醌氧化还原酶亚组K BP028387 10.183 53.625 128.167 NADH锟氧化还原酶亚组2 Table 4. Related gene expression levels between different treatment in oxidative phosphorylation pathway (RPKM value)
-
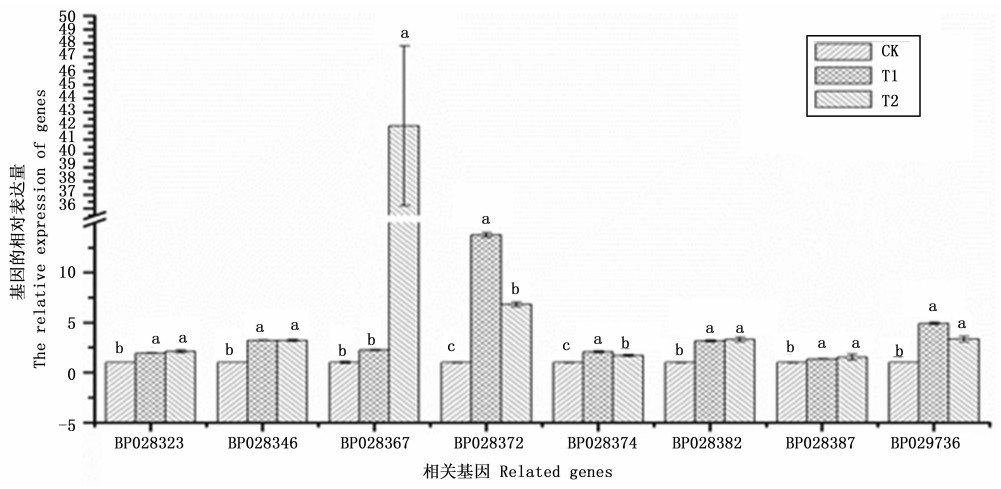
为验证转录组测序结果的可靠性, 分别以T1、T2和CK白桦顶尖组织的RNA为试材, 对光合作用途径表达差异显著的4条基因和氧化磷酸化途径表达差异显著的4条基因进行qRT-PCR分析, 结果显示: 与CK相比, T1、T2中上述8条基因均呈上调表达, 与Illumina/Solexa测序结果具有相同的表达趋势(图 7), 说明测序结果是可靠的。

Figure 7. Real-time quantitative PCR to verify differentially expressed between each candidate gene samples
2.1. 不同浓度赤霉素GA4+7对白桦无性系生长的影响
2.2. 不同浓度GA4+7处理下白桦转录组数据统计及文库质量评价
2.3. 不同浓度GA4+7处理下白桦无性系的差异表达基因
2.4. 差异基因的GO分类及富集分析

2.5. 赤霉素处理下白桦无性系生长相关基因的初步分析
2.5.1. 光合作用途径相关基因
2.5.2. 氧化磷酸化途径相关基因
2.5.3. qRT-PCR验证
-
调节树木生长, 使其短时间内获得较大生长增益从而缩短育种周期是林木育种的重要课题, 人们通常采取施肥、环割、断根、施加外源激素等措施促进树木生长[7-16-18]。试验证明, 对于果材兼用树种施加GA4+7在促进开花及生长、提高座果率方面效果显著, 因此, 备受育种工作者的重视[8-9]。
为了探讨GA4+7在促进白桦生长方面的作用, 试验利用遗传组成完全相同的1个白桦无性系为材料, 分别喷施0.2 g·L-1和0.4 g·L-1 GA4+7, 结果表明, 施加GA4+7对白桦苗期生长具有促进作用, 在苗高方面, 2个浓度的GA4+7均能显著促进苗高生长, 0.2 g·L-1和0.4 g·L-1 GA4+7处理2年后无性系苗高均值分别高于CK的9.05%、19.65%;在地径方面, 在GA4+7连续处理2年后地径均值显著高于CK, GA4+7浓度为0.4 g·L-1时对白桦苗期生长促进效果好。这一实验结果与Bottini等人[9]对苹果(Malus pumila)树的研究结果相吻合, 即赤霉素促进植株的营养生长。
GA4+7处理下白桦无性系生长量的增加, 必然与生长相关基因的上调表达密切关联, 对白桦苗木生长旺盛期的转录组测序也证明了0.4 g·L-1 GA4+7(T2)处理苗高增长显著, 同时与对照(CK)之间差异基因的数量也最多, 即上调表达的基因有181个, 下调表达的基因有55个(图 2), 这些差异基因在叶绿体类囊体膜、红光、远红光及蓝光的细胞响应等方面富集明显(图 5), 进一步Pathway富集分析发现: 叶绿体中CF1ɑ亚组ATP合成酶、细胞色素b6f亚组4、光系统Ⅱ蛋白D1、光系统ⅠP700叶绿素A脱辅基蛋白A2等4条光合代谢相关的基因呈上调表达, 这些基因参与光合作用的光反应过程。另外还有氧化磷酸化途径的ATP合成酶F1亚组1、细胞色素c氧化酶亚组1、NADH-质体醌氧化还原酶亚组K、NADH锟氧化还原酶亚组2等4条基因呈上调表达, 上述8条基因表达量的升高, 可能是通过加快电子传递过程、催化ATP的合成, 而促进植物的光合作用增强[19-25]。总之, GA4+7处理后白桦通过上调光合途径和氧化磷酸化途径的相关基因的表达, 加快其细胞内有机物的合成, 进而促进其苗高、地径的生长。由于参试的白桦无性系放置于塑料大棚外, 按常规管理这些苗木进入开花结实期还需2~3年的时间, GA4+7处理后能否加快白桦营养生长向生殖生长转变进程还有待于后续研究, 按照前期试验设计继续施加GA4+7, 观察试验组是否缩短童期、是否增加开花数量提高种子产量, 进而在基因表达水平上探讨GA4+7对白桦生长的影响。
-
本研究外源施加赤霉素GA4+7可促进白桦苗木的高生长和地径生长。转录组测序结果表明: 赤霉素GA4+7处理前后的差异基因主要富集在叶绿体类囊体膜、红光、远红光及蓝光的细胞响应等方面; 同时赤霉素GA4+7可上调氧化磷酸化途径和光合途径相关基因的表达, 研究结果可为深入了解赤霉素GA4+7对白桦生长的调控机制及生长相关途径重要基因的克隆提供参考。
 DownLoad:
DownLoad: