




-
海拔梯度包含了温度、湿度、光照和土壤等诸多环境因子的剧烈变化,使其成为研究植物对环境变化响应的理想场所[1]。海拔梯度环境条件的异质性会导致林线树木在形态和生理上产生一定的变化[2]。高海拔树木能够生存、生长发育和繁衍取决于对剧烈变化的环境条件的生理生态响应和适应能力。所以,生长在高海拔环境的树种需要充足的碳水化合物供应支持其生长和代谢。树木体内积累的非结构性碳水化合物(non-structural carbohydrates, NSC)能够为树木休眠后恢复生长提供碳源,而且能在植物光合产物不足时起到缓冲作用,因此在很大程度上影响着植株的生长及对环境的响应[3]。NSC积累是否充足,能够反映出树木是否受到碳限制。NSC在树木体内的积累有季节性和组织之间的差异,而组织之间的差异可以反映出碳源和碳库之间的平衡关系[4-5]。以往的大量研究,主要集中在生长季节树木体内海拔梯度对碳水化合物含量的影响[2, 6]。但是有人提出,在比较不同海拔梯度间的树木体内碳水化合物差异时应考虑到在同一发育时期或者是在同一物候期内比较(比如生长末期或者是休眠期)[7-8]。而对于非生长季内不同海拔梯度上林线树种碳水化合物含量变化的报道较少。非生长季内林线树种体内的NSC既要保证树种在低温环境下安全过冬,又提供来年新枝叶萌发所需要的营养。所以林线树种的冬季适应策略,决定着在林线树种的生存能力。
青海云杉(Picea crassifolia Kom.)是祁连山森林的建群种,是祁连山区重要的水源涵养林,随海拔垂直分布,生长状况存在明显的海拔差异。目前对祁连山林线树种碳水化合物含量及其变化的报道相对较少[9],尤其是青海云杉休眠期的研究还未见报道。本研究以祁连山林线树种青海云杉为对象,研究休眠前后不同海拔青海云杉成年树当年和1年生中枝条及叶片中NSC及其组分含量的变化,探讨青海云杉冬季的适应策略,为深入研究祁连山林线形成机制提供科学依据。
HTML
-
研究区域设在祁连山北坡中部祁连山水源涵养林西水自然保护站地区(38°34′ N,100°17′ E)。属高寒干旱半干旱地区,温度、降水具有明显的垂直梯度和水平差异。年均温度为-0.6~2.0℃,7月份平均气温10~14.0℃。年降水量在300~500 mm之间,其中60%以上集中在6—9月份,相对湿度50%~70%,年蒸发量1 200 mm,无霜期90~120天[8]。水热条件的垂直分布造成植被分布具有明显的垂直地带性,从低海拔到高海拔依次为荒漠草原植被带、干性灌丛草原植被带干性灌丛草原植被带、山地森林草原植被带、亚高山灌丛草甸植被带、高山寒漠草甸植被带。青海云杉是祁连山森林的建群种,水源涵养林的主体,分布海拔范围在2 500~3 300 m的阴坡、半阴坡。土壤为山地灰褐土。
-
分别于2013年9月(休眠前)、2014年5月(休眠后),采集青海云杉成年树(树龄1约50年)当年生及1年生枝条、叶片。沿海拔2 900~3 300 m海拔之间设(2 900、3 000、3 100、3 200、3 300 m)5个采样区(设定2 900 m为低海拔、3 000~3 100 m为中海拔、3 200~3 300 m为高海拔),其中海拔3 300 m为林线的上缘,3 200 m为林线的过渡带。在每个采样区上选择个体大小和生长状况一致的健康成年青海云杉各5棵。每次采样时间为12:00—14:00。在每株树冠的中上部采集不同方位1年生及当年生枝条和叶片,不同方位样品混合后为1个样本,纸袋封装后放在冰盒里带回。80℃烘箱杀青,70℃烘干,粉碎成粉末用于测定使用。
-
可溶性糖提取:参照于建国等[10]著《现代使用仪器分析方法》,略有改动。称1 g粉碎好的干燥植物样品于三角瓶中,加入50 mL蒸馏水,用锡纸封住瓶口,放入高压锅中蒸1小时,冷却后保证三角瓶中有20 mL溶液,过滤,滤液用Waters 244型高效液相色谱仪(美国)测定可溶性糖(果糖、葡萄糖)的含量。
淀粉提取:参照刘福岭等[11]著《食品物理化学分析方法》,略有改动。称取2 g粉碎好的干燥样品于三角瓶中,用乙醚洗涤4~5次,然后用10%乙醇溶液150 mL分次洗涤,弃去溶液,将残留物用蒸馏水200 mL洗入三角瓶中,加稀盐酸(2∶1)20 mL,置沸水浴中回流8小时,冷却后,用20%氢氧化钠中和,加入铝浆10 mL,转移至500 mL容量瓶中,用蒸馏水稀释至刻度,摇匀,过滤,滤液用Waters 244型高效液相色谱仪(美国)测定淀粉含量。
Waters 244型高效液相色谱仪(美国)色谱条件——色谱柱:sugar-pak 1(美国);流动相:蒸馏水;流速:0.6 mL·min-1;柱温:70℃;检测器:视差检测器。
NSC为可溶性糖和淀粉含量的总和,其中可溶性糖包括葡萄糖和果糖两部分,所有含量均用百分比干重表示。
-
所有数据均采用SPSS16.0软件进行分析。采用影响因子方差分析(ANOVA)分析海拔、采样时间及枝条年龄对非结构性碳水化合物含量的影响;采用one-way ANOVA方法分析青海云杉组织中NSC及其组分含量在不同海拔间的差异显著水平,并进行LSD多重比较分析。
1.1. 采样地点
1.2. 样品采集
1.3. 样品分析方法
1.4. 数据分析
-
本研究影响采样的因素包括采样时间和海拔梯度,通过影响因子方差法(ANOVA)分析了采样时间、海拔梯度对青海云杉枝条中非结构性碳水化合物(NSC)含量的影响(表 1)。结果显示,海拔梯度对当年生叶片中葡萄糖、可溶性糖含量以及糖/淀粉比值影响显著(P<0.05),对果糖、淀粉及NSC无显著影响(P>0.05)。而对1年生叶片中的各组分及糖/淀粉比值均有显著影响(P<0.05)。海拔梯度对当年生枝条中各组分的影响结果与1年生叶片相同。而在1年生枝条中,海拔只对果糖和NSC含量有显著影响(P<0.05)。
项目Items F df 葡萄糖
glucose果糖
fructose可溶性糖
Total Sugar淀粉
StarchNSC
NSC糖/淀粉
Sugar/starch当年叶
Current-year leaves时间(T) 1 0.637 166.55*** 6.042* 422.15*** 1 045.53*** 15.904** 海拔(E) 4 4.965* 2.237 3.668* 1.632 1.834 3.373* T*E 4 3.120* 3.354* 3.008* 6.562** 7.419** 3.511* 1年叶
One-year leaves时间(T) 1 12.529** 44.03*** 0.07 821.13*** 1 032.79*** 61.14*** 海拔(E) 4 12.253*** 3.777* 10.232*** 4.993** 9.162*** 7.065** T*E 4 4.797** 1.676 4.072* 17.313*** 13.146*** 7.856** 当年枝
Current-year stems时间(T) 1 8.618** 312.549*** 94.333*** 87.927*** 249.102*** 11.122** 海拔(E) 4 28.128*** 33.945*** 28.161*** 4.299* 8.54*** 5.347* T*E 4 5.168** 30.91*** 6.429** 3.471* 10.528*** 0.254 1年枝
One-year stems时间(T) 1 0.282 10.192** 0.512 57.196*** 261.917*** 10.002** 海拔(E) 4 1.052 3.327* 1.577 0.867 8.872*** 1.058 T*E 4 0.530 0.559 0.483 0.634 2.016 0.250 Table 1. ANOVA of factors influencing concentrations of NSC and its components of Picea crassifolia
采样时间对各组织中非结构性碳水化合物NSC组分含量影响表明,当年生叶中除葡萄糖含量外,其它各组分含量及糖/淀粉比值均受采样时间的显著影响(P<0.05)。1年生叶和当年生枝条中NSC各组分含量及糖/比值受采样时间显著影响(P<0.05)。1年生枝条中果糖、淀粉、NSC含量及糖/淀粉比值受采样时间显著影响(P<0.05),葡萄糖和可溶性糖含量未显著受到采样时间的影响(P>0.05)。
当年生叶、1年生叶及当年生枝条中所测各组分均受到海拔×采样时间交互作用的显著影响(P<0.05),而1年生枝条中所测各组分均未受到海拔×采样时间的显著影响(P>0.05)。
-
青海云杉不同组织可溶性糖含量随拔变化(图 1)。由图 1可以看出,休眠前当年生叶中可溶性糖含量随海拔升高呈上升趋势,在林线过渡带海拔3 200 m达到最高值,海拔间可溶性糖含量没有显著差异(P>0.05)。1年生叶中可溶性糖含量随海拔升高呈波动式变化,总体呈上升趋势,在海拔3 200 m达最高值,而在林线边缘3 300 m又有所下降,高海拔(3 200 m)、中海拔(3 100 m)与低海拔2 900 m相比可溶性糖含量显著提高(P<0.05),而3 300 m可溶性糖含量与其它各海拔间没有显著差异(P>0.05)。当年生枝和可溶性糖含量随海拔呈上升趋势,在330 m出现最高值,高海拔(3 200~3 300 m)当年生枝中可溶性糖含量显著高于低海拔(2 900 m)。1年生枝条中可溶性糖含随海拔变化趋势与当年生叶片相同。

Figure 1. The contents of soluble sugar in different tissue of Picea crassifolia before and after dormancy
休眠后,当年生叶中可溶性糖含量随海拔升高呈降低-升高-降低-升高的波动式变化,最高值在中海拔3 100 m出现,海拔间没有显著差异(P>0.05)。1年生叶可溶性糖含量随海拔变化趋势与当年生叶相同,但最高值出现在高海拔3 300 m,高海拔3 300 m可溶性糖含量显著高于低海拔2 900 m及中海拔3 000 m(P<0.05),而与其它海拔间差异不显著(P>0.05)。当年生枝条中可溶性糖含量随海拔升高呈升高-降低-升高的变化趋势,林线边缘3 300 m出现最高值,显著高于低海拔2 900 m和林线过渡带3 200 m(P<0.05)。1年生枝条中可溶性糖含量随海拔的变化趋势与当年生叶相同,最高值出现在3 300 m,显著高于低海拔2 900 m、中海拔3 000 m及林线过渡带3 200 m可溶性糖含量(P<0.05)。
葡萄糖含量在可溶性糖含量中占得比重高于果糖,但是休眠后果糖在可溶性糖含量中的比重增加。休眠前,各组织中可溶性糖中果糖占得比重18%~33%,而休眠后占得比重31%~44%。
-
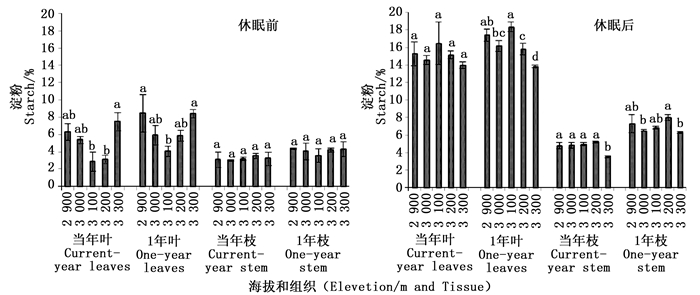
青海云杉各组织中淀粉含量随海拔变化如图 2。当年生叶中淀粉含量随海拔升高先下降,在海拔3 100 m达到最低值,随后又逐渐升高,最高值在海拔3 300 m出现,且显著高于海拔3 100 m和3 200 m的淀粉含量(P<0.05),与低海拔2 900 m没有显著差异(P>0.05)。1年生叶中淀粉含量随海拔变化趋势与当年生叶相同,随海拔升高淀粉含量降低,最低值在3 100 m,显著低于其它各海拔淀粉含量(P<0.05)。当年生枝淀粉含量随海拔升高呈降低-升高-降低的变化趋势,但各海拔间均没有显著差异(P>0.05)。1年生枝中淀粉含量随海拔升高呈先降低后升高的变化趋势,最低值出现在中海拔3 100 m,且各海拔间没有显著差异(P>0.05)。

Figure 2. The contents of starch in different tissue of Picea crassifolia before and after dormancy
休眠后,当年生叶中淀粉含量随海拔升高先降低在升高,最高值在3 100 m,而后随海拔升高淀粉含量又有所降低,海拔间没有显著差异(P>0.05)。1年生叶中淀粉含量随海拔升高呈降低-升高-降低的变化趋势,最高值在中海拔3 100 m,最低值在高海拔3 300 m。方差分析表明,海拔低海拔(2 900 m)、中海拔(3 100 m)的淀粉含量显著高于高海拔(3 200 m和3 300 m)(P<0.05)。当年生枝中淀粉含量随海拔逐渐升高,而在3 300 m淀粉含量呈下降趋势,最低值在海拔3 300 m,显著低于其它各海拔的淀粉含量(P<0.05)。1年生枝中淀粉含量随海拔升高呈降低-升高-升高的变化趋势,最高值在海拔3 200 m,显著高于海拔3 300 m和3 000 m的淀粉含量。比较休眠前后各组织中的淀粉含量发现,休眠后各组织总的淀粉含量均有明显增加。
-
青海云杉非结构性碳水化合物(NSC)含量随海拔变化如图 3。休眠前,当年生叶中NSC含量随海拔升高整体呈上升趋势,最高值在海拔3 300 m,显著高于低海拔和中海拔(P<0.05)。1年生叶中的NSC含量随海拔升高呈先降低在升高的变化趋势,最低值在海拔3 000 m出现,最高值在林线边缘3 300 m,中海拔(3 000 m和3 100 m)的NSC含量显著低于高海拔和低海拔的NSC含量(P<0.05)。当年生枝NSC含量随海拔升高呈上升趋势,但海拔间差异不显著(P>0.05)。1年生枝条中NSC含量随海拔升高先逐渐降低,而后升高海拔3 200 m达最高值,到海拔3 300 m时又有所下降,海拔间没有显著差异(P>0.05)。

Figure 3. The contents of soluble sugar in different tissue of Picea crassifolia before and after dormancy
休眠后,各组织中的NSC含量明显高于休眠前。当年生叶和1年生叶随海拔变化趋势相同,均是随海拔升高呈下降-升高-下降的变化趋势。最高值均出现在中海拔3 100 m,最低值出现在高海拔3 300 m,最高值和最低值间差异显著(P>0.05)。当年生枝中NSC随海拔升高呈先升高在下降的变化趋势,最低值在海拔3 300 m,中海拔(3 000 m和3 100 m)及林线过渡带(3 200 m)的NSC含量显著高于3 300 m和2 900 m。1年生枝条中NSC含量随海拔升高与当年生叶的变化趋势形同,最高值在3 200 m出现,而最低值则在3 000 m,且海拔3 000 mNSC含量显著低于其它各海拔(P<0.05)。
-
休眠前所测各组织中糖/淀粉比值如表 2。当年生叶片中的比值均呈先升高,再降低的变化趋势,最高值在3 200 m处为2.888,显著高于低海拔2 900 m及林线边缘3 300 m的比值。1年生叶糖/淀粉比值的最高值在3 100 m处,随后比值随海拔升高呈降低趋势,3 100 m的比值显著高于其它各海拔。当年生枝条糖/淀粉各海拔比值均1.0~2.0之间,最高值(1.643),在海拔3 300 m,且海拔间没有显著差异。1年生枝糖/淀粉比值最高值(1.781),也是在海拔3 300 m,且海拔间差异不显著。
海拔
Elevation/m当年叶
Current-year leaves1年叶
One-year leaves当年枝
Current-year stems1年枝
One-year stems2 900 0.896a 0.724b 1.197a 0.968a 3 000 1.152a 0.949b 1.249a 1.106a 3 100 3.255a 2.078a 1.403a 1.419a 3 200 4.507a 1.251ab 1.251a 1.260a 3 300 0.834a 1.643b 1.643a 1.781a 不同小写字母表示不同海拔间的差异显著(Duncan多重比较,P<0.05)
Different latters in each column indicate significat differences between elevations(Duncan multiple comparisons at P<0.05)Table 2. The ratio of soluble sugar concentration and starch concentration in tissues of Picea crassifolia at different elevations before dormancy
休眠后多测各组中的糖/淀粉比值如表 3。各组织中的糖/淀粉比值均在0.5~1.5之间,与休眠前相比明显降低。所测各组织中糖/淀粉比值最高值均出现在海拔3 300 m,当年叶(0.578)、1年叶(0.553)、当年枝(1.490)、1年枝(0.906)。当年叶糖/淀粉比值在各海拔间没有显著差异。1年叶、当年生枝、1年生枝在海拔3 300 m的比值显著高于其它各海拔。
海拔
Elevation/m当年叶
Current-year leaves1年叶
One-year leaves当年枝
Current-year stems1年枝
One-year stems2 900 0.516a 0.368b 0.925b 0.659c 3 000 0.518a 0.359b 1.049b 0.614bc 3 100 0.540a 0.390b 1.024b 0.781b 3 200 0.493a 0.425b 0.944b 0.611c 3 300 0.578a 0.553a 1.490a 0.906a 不同小写字母表示不同海拔间的差异显著(Duncan多重比较,P<0.05)
Different latters in each column indicate significat differences between elevations(Duncan multiple comparisons at P<0.05)Table 3. The ratio of soluble sugar concentration and starch concentration in tissues of Picea crassifolia at different elevations after dormancy
休眠前糖/淀粉比值明显高于休眠后,休眠前可溶性糖占得比重高,而休眠后淀粉含量增加,糖/淀粉比值降低。
2.1. 海拔、休眠对青海云杉NSC含量的影响
2.2. 休眠前后青海云杉不同组织中非结构性碳水化合物含量随海拔变化
2.2.1. 可溶性糖含量随海拔变化
2.2.2. 淀粉含量随海拔变化
2.2.3. 非结构性碳水化合物含量随海拔变化
2.2.4. 可溶性糖/淀粉比值随海拔变化
-
本研究发现,休眠前青海云杉当年生叶及1年生叶中NSC在高海拔(3 300 m和3 200 m)的含量不低于中低海拔(3 000 m和3 100 m),而在枝条中高中低海拔间没有显著差异。说明休眠前青海云杉各组织中NSC积累未受到海拔的限制,这与Shi[12]周永斌[6]、王彪[13]等同时期的研究结果一致。研究表明高海拔树种光合作用强度与低海拔没有明显差异[14-15],所以在生长季高山林线树木能够合成足够的光合产物,同时低温环境降低了树木的呼吸消耗使得树木体内积累大量的NSC。这可能是休眠前高海拔青海云杉NSC含量不低于中低海拔的原因之一。利用14C示踪方法对不同组织的NSC研究发现,树木呼吸代谢利用的碳水化合物主要是当年合成的NSC,而1年或2年形成的NSC主要用于年轮的生长和休眠期的代谢[16-17]。因此常绿树种当年生及1年生枝条叶片中的NSC对树木休眠和再生长很重要。可见休眠前青海云杉枝条和叶片中积累的NSC既能保证其安全越冬,也为来年新组织生长提供碳源。
休眠后青海云杉NSC含量表现为:海拔间比较,中海拔(3 100 m)叶中NSC含量著高于高海拔(3 200 m和3 300 m),当年生枝条NSC含量高于高海拔(3 300 m)与低海拔(2 900 m);休眠前后比较,各组织中的NSC含量显著高于休眠前,叶片中尤为明显。造成这种现象的原因一方面中海拔与高海拔相比温度相对较高,与低海拔相比光照充足,所以中海拔更有利于光合产物的形成和积累。造成这种现象的生理机制还有待于结合土壤温度、水分、休眠后各海拔的气温变化以及光照条件深入研究。另一方面休眠后的5月份气温上升,青海云杉针叶开始进行光合作用,而这时土壤温度还很低,根系活动受到限制,所有光合产物的利用率降低,大量的光合产物以淀粉的形式积累在叶片和枝条中,造成了淀粉含量的显著升高。另外,以往的研究表明常绿树在休眠后萌发前,枝条和针叶中淀粉含量会有一个显著的升高过程,保证幼枝在萌发过程中有充足的碳源[18-20]。青海云杉符合针叶树种的这一特性,淀粉含量的增加保证了新的枝叶萌发所需要的碳水化合物。所以,青海云杉休眠前后NSC含量的变化特点是林线树木长期适应高海拔恶劣环境的结果。
-
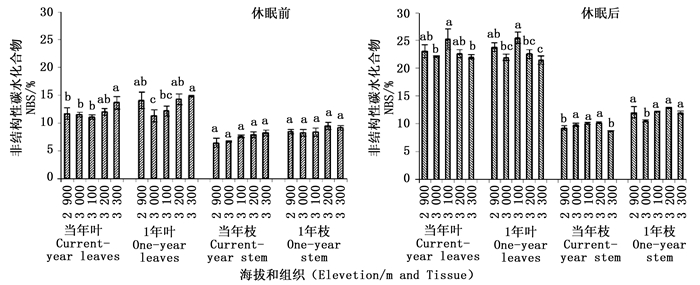
非结构性碳水化合物(NSC)包括淀粉、果糖、葡萄糖、蔗糖及果聚糖等多种成分,未来气候变化不仅会对NSC总量产生一定影响,更重要的是还可能会进一步影响到NSC中各组分的组成和含量[21]。可溶性糖通过提高组织细胞液渗透压,防止细胞内结冰,提高植物抗寒性,顺利度过寒冷季节。休眠前后,各组织在高海拔的可溶性糖含量高于中低海拔或者是没有显著差异。休眠后可溶性糖含量增加,果糖在可溶性糖含量中所占比重上升。造成青海云杉体内较高的可溶性糖含量的原因一方面是为了增加细胞液的浓度防止细胞结冰,安全越冬。另一方面可溶性糖的大量积累可能是林线树木长期进化中对低温环境适应的一种策略。以往的研究也表明,植物中的可溶性糖不仅是能量来源和结构性物质的重要组成元件,还具有信号物质的功能,调节相关基因的表达和酶活性,参与植物的生长发育调节以及应对不良环境[22-24]。迄今已证明,具有重要作用的糖信号主要是二糖和单糖,而单糖主要是葡萄糖和果糖[25-28]。分析认为青海云杉休眠前后可溶性糖组分的变化,可能是葡萄糖和果糖起到了信号物质的作用来调节对低温环境的感知和适应,同时单糖对休眠后青海云杉新生组织的发育及分化也可能起到了信号调节的作用。但是,可溶性糖组分的这种变化究竟又怎样的生理机制,还有待于进一步深入研究。
可溶性糖与淀粉之间的相互转化是植物适应逆境胁迫的一种有效方式,而它们的比例关系在高山植抵御低温中起着关键作用。李迈和等的研究发现,高海拔常绿树木在越冬过程中糖/淀粉比值大于3,才能安全越冬[16]。但本研究的结果发现休眠前后青海云杉在各海拔枝条和叶片中糖/淀粉的比值随海拔升高虽然呈上升趋势但变化范围在0.70~4.5之间,休眠后比值均在0~2之间。这与以往的一些研究结果不同[13, 29]。分析认为由于青海云杉组织中休眠前后积累较多的淀粉使其比值降低,这一方面为应对低温做好可溶性碳水化合物的储备,另一方面为来年新组织的生长积累能源物质。
3.1. 海拔对休眠前后青海云杉不同组织中NSC含量的影响
3.2. 海拔对休眠前后青海云杉不同组织可溶性糖、淀粉含量及其比值的影响
-
本研究通过对休眠前后青海云杉不同组中的非结构性碳水化合物在海拔间变化的研究,发现青海云杉冬季的NSC含量表现为:不同海拔青海云杉休眠前积累足够的NSC,而休眠后光合产物的及时补充,保证了林线树种青海云杉安全越冬,并提供给新生组织发生所需的碳源。这是林线树种青海云杉在低温环境下确保物种延续的一种生存策略。
 DownLoad:
DownLoad: