



-
金属矿山在开采、选矿过程中会产生大量的废弃尾矿砂,金属尾矿由于含有大量高浓度重金属,给周边环境和人类带来污染和危害[1]。重金属在人体较低摄入的情况下,即可对人体产生明显的毒副作用,如镉能蓄积在肝、肾组织中,引起肾脏吸收功能不全;同时重金属极易通过食物链在生物体内积累,并难以通过生物降解而消除,从而对人体健康产生极大危害[2]。尾矿不仅危害人体健康、污染周围环境,而且破坏生态系统,如何在退化的尾矿生态系统中进行植被恢复与植物修复已成为目前环境生态学的研究热点[3]。利用耐性或富集植物对废弃尾矿进行植物修复是当前国际上常用的措施,在美国、英国和澳大利亚等国家,一些对重金属有高耐受性植物的培育已经商业化[4]。
应用柳树(Salix spp.)对重金属污染土壤进行修复的研究与实践在国外已有较多报道[5-9]。近10年来,国内在柳树对重金属的吸收和积累方面的研究也越来越多,但大多数研究是基于室内水培试验的结果[10-11]。目前有关旱柳(S. matsudana Koidz.)、杞柳(S. integra Thunb.)以及金丝柳(Salix × aureo-pendula CL.)等不同柳树品种、无性系在水培条件下对Cd、Zn、Pb等重金属的吸收和积累规律的研究已有很多报道[12-15],然而,应用柳树进行金属尾矿的修复研究还很少。施翔等[16]采用Cu/Zn复合污染的矿砂栽培旱柳等7个柳树无性系,发现柳树叶片对Zn具有较高的转移能力;廖启林等[17]研究了栽种柳树修复镉污染土壤的状况,表明栽种苏柳795、苏柳172对原位修复我国南方农田土壤的镉污染具有广阔的应用前景。
金属尾矿不仅重金属含量高,而且具有肥力低、缺乏土壤微生物等多种限制植物生长的因素,因此,了解植物材料在尾矿中的生长和耐性对评价其在金属尾矿植被恢复中的潜力具有重要意义。全世界柳树约有526种,产于我国的有257种[18],因此,研究柳树不同基因型在金属尾矿的生长响应对开发和利用我国丰富的柳树资源具有重要意义。本研究在前期试验基础上,以旱柳和杞柳无性系为材料,研究了不同柳树无性系在Cd、Zn、Cu 3种重金属复合污染条件下的生长、耐性以及光合作用、叶绿素含量等生理生化响应,为柳树在金属尾矿废弃地的植被恢复提供理论依据。
HTML
-
参试材料分别为旱柳739、470、30和16号,共4个无性系,杞柳‘一枝笔’(S. integra ‘Yizhibi’)和‘大红头’(S. integra ‘Dahongtou’)2个栽培品种,柳树插条均采自浙江省杭州市富阳区新登柳树种质资源收集圃。
试验所用铜矿尾矿砂采自浙江省绍兴平水铜矿,对照采用无土介质泥炭+珍珠岩(3∶1)。污染土样采集自土壤表层(0~30 cm),其物理化学性质见表 1。根据国家标准GB 15618-1995,污染介质中的重金属含量显著高于标准值[16]。
土壤类型
Soil type有效磷
Available Phosphorus/(mg·kg-1)水解氮
Hydrolyzed Nitrogen/(mg·kg-1)有效钾
Available Potassium/(mg·kg-1)镉
Cadmium/(mg·kg-1)铜
Copper/(mg·kg-1)锌
Zinc/(mg·kg-1)pH值 铜尾矿砂
Copper tailings1.05 18.00 57.2 2.28 596.0 824 6.62 对照
Control10.76 0.64 21.0 — 18.5 56 7.01 Table 1. Physical and chemical characteristics of tested soil
-
选择粗细均匀的柳树枝条剪成长6~8 cm的插穗,扦插在沙床上,置于光照良好处,不定期浇水。约90 d后选择生长基本一致的生根苗移植到装有约3 kg试验土壤的圆形塑料盆(口径15 cm,高15 cm)内。试验按无性系与介质两因素裂区,以介质差异为主因素,柳树无性系为副因素,采用随机区组设计,设2个重复区组,每个区组的每个处理又重复3盆,共72盆,随机排列(水平内无性系间)。试验在浙江杭州市富阳区中国林业科学研究院亚热带林业研究所温室大棚进行。试验期间大棚白天温度25~35℃,夜间温度15~20℃。
6月中旬将生根苗移植到塑料盆中,10月中旬收获苗木,试验周期总共120 d。试验期间7—9月,每月进行1次光合作用参数测定。植物收获后测定其叶绿素含量、株高、地径、生物量以及根系形态学参数。
-
植株收获以后,将植株分根、茎、叶3部分,105℃杀青,75℃烘干至恒质量后称其干质量。
-
植物根系去离子水洗净后,用双光源扫描仪扫描。采用WinRHIZO PRO 2007根系分析系统软件(Regent Instruments Inc.,Canada)分析根长、根表面积、根体积以及不同径级根长和表面积等形态学参数。
-
于7—9月,选择晴朗的天气,上午8:00—11:00时,选择3株苗中部的3片功能叶片测定其光合作用。采用Li-6400便携式光合作用系统(Li-COR,USA),主要测定叶片净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)等指标。
-
取处理和对照植株的成熟功能叶片测定叶绿素含量,共4次重复。叶绿素含量测定采用80%丙酮提取法[19]。
-
耐性指数[20]=尾矿砂中生物量/对照组生物量×100%
抑制率(降幅)=(对照-处理)/对照×100%
-
用Microsoft Excel 2003软件进行简单的数据处理,采用DPS7.05软件进行方差分析,用Origin7.5软件进行绘图。采用LSD法对6个柳树无性系在2种土壤介质中的生理反应数据进行差异性比较。主成分分析用SPSS 22.0分析软件完成。
1.1. 试验材料
1.2. 试验设计
1.3. 测定方法
1.3.1. 生物量测定
1.3.2. 根系形态学参数测定
1.3.3. 光合作用测定
1.3.4. 叶绿素含量测定
1.3.5. 耐性指数(TI)及各参数抑制率(降幅)计算
1.4. 统计分析
-
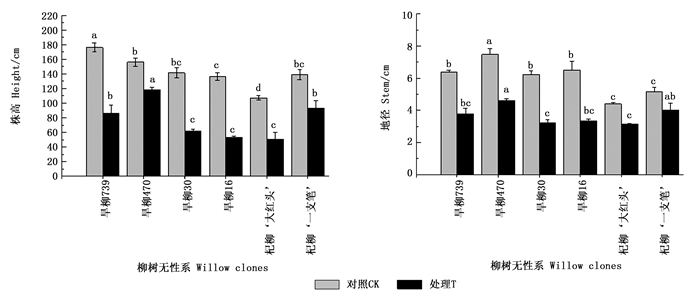
由图 1可知:6个柳树无性系之间的株高和地径生长在各处理间均差异显著(P<0.05),在对照条件下,旱柳739的株高生长明显优于其他无性系,而地径生长则旱柳470最优(图 1)。在铜尾矿砂培养条件下,6个无性系的株高和地径生长均受到不同程度的抑制,其中,株高降幅最小的是旱柳470无性系(24%),地径降幅最小的是杞柳‘一枝笔’(22%)。旱柳30和旱柳16无性系的株高和地径降幅最大(48%~61%),说明在重金属复合污染条件下,这2个无性系的生长受到较严重的抑制作用。

Figure 1. Growth index of six willow clones under two treatments
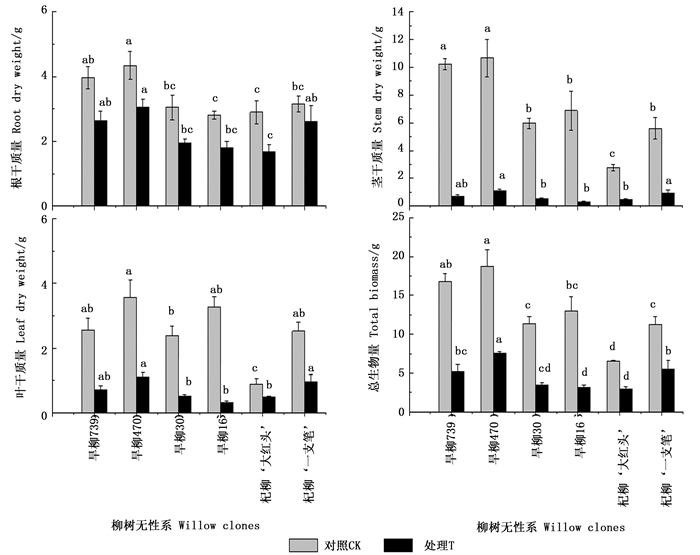
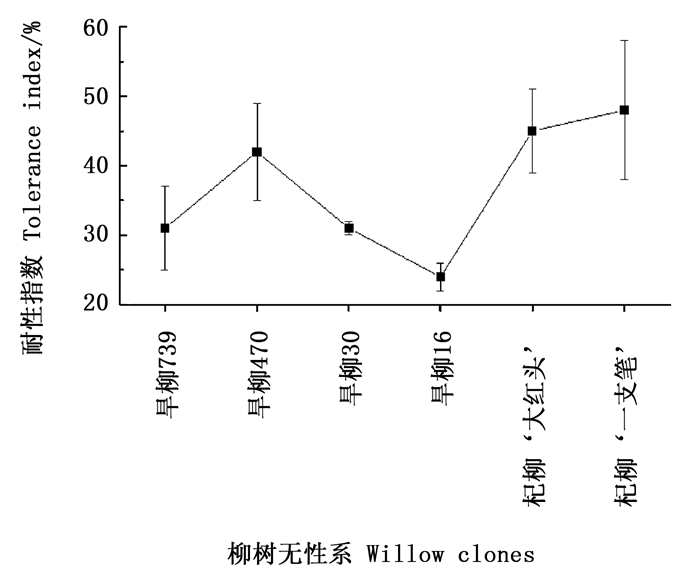
生物量是衡量植物生长状况的重要指标,由图 2可看出:铜尾矿砂中生长的柳树无性系,地上部和根部生物量以及总生物量均显著下降(P<0.05),说明重金属污染显著抑制柳树生物量生产;但铜尾矿砂对不同组织生物量的抑制程度不同,其中,对根部生物量的影响最小,6个无性系的降幅(抑制率)只有16%~42%,降幅大小依次为:杞柳‘大红头’>旱柳30>旱柳16>旱柳739>旱柳470>杞柳‘一枝笔’;铜尾矿砂对叶片和茎干质量的影响最大,其中,对旱柳16的叶片和茎生物量的抑制最高,抑制率分别为91%和86%,表明在重金属污染条件下,地上部生长更容易受到影响。总生物量的下降趋势与茎生物量相似,抑制率为51%~76%,不同无性系抑制率大小依次为:旱柳16>旱柳739=旱柳30>旱柳470>杞柳‘大红头’>杞柳‘一枝笔’。根据不同无性系耐性指数的比较(图 3)发现:杞柳的耐性指数总体而言大于旱柳。

Figure 2. Biomass of six willow clones under two treatments

Figure 3. Tolerance index of six willow clones under mine tailings
进一步对不同无性系的根系形态学参数进行分析,结果(表 2)发现:尽管根部生物量受到重金属污染的影响较小,但与对照相比,各无性系的根长、根表面积、根体积、根尖数在铜尾矿砂中出现下降趋势,说明供试柳树无性系在重金属污染条件下根系的生长发育受到抑制,且抑制程度在不同无性系之间的差异显著(P<0.05)。从各无性系根系形态学参数的下降幅度看,旱柳16无性系的根长、根表面积、根体积和根尖数降幅最大,其中,根体积与对照相比下降了96%,而旱柳470无性系根系各项参数的下降幅度较小。根系总长度、根系表面积、体积等形态学参数是决定根系养分吸收范围、吸收强度的重要指标[21],因此,重金属污染条件下柳树无性系总根长、根体积以及根表面积等参数的下降导致根系对营养物质的吸收范围缩小,吸收强度下降,推测这也是导致其生物量下降的原因之一。
柳树无性系
Clones处理
Treatment根长
Root length/cm根表面积
Root surface area/cm2根体积
Root volume/cm3根尖数
Root tips旱柳739 CK 2 649.59±238.51b 409.14±20.68b 5.11±0.35bc 12 497.86±3 224.71d T 688.50±47.21b 97.64±11.22bc 0.97±0.32b 4 308.54±774.66c 旱柳470 CK 3 320.35±129.16b 503.12±28.31b 5.53±0.99bc 15 125.00±740.77d T 1 639.57±97.17a 202.99±16.76a 1.81±0.40a 9 122.05±513.38bc 旱柳30 CK 2 392.35±149.80b 137.63±7.18b 4.23±0.63bc 15 347.00±1 001.01d T 1 219.58±124.85ab 1 219.98±124.85ab 1.05±0.08b 8 305.95±293.61bc 旱柳16 CK 8 384.48±1 600.15a 1 272.92±246.64a 9.76±3.17a 54 284.10±10 349.77b T 754.76±46.08b 65.48±6.27c 0.39±0.01 c 10 067.85±552.52b 杞柳‘大红头’ CK 3 921.44±400.52b 410.52±146.41b 2.56±1.00c 31 876.80±2 788.60c T 1 702.48±496.53a 126.74±59.89bc 0.77±0.23bc 20 403.69±5 378.36a 杞柳‘一枝笔’ CK 7 491.01±1 955.55a 713.59±364.13b 6.83±3.71ab 83 459.91±18 030.91a T 1 565.74±628.14a 148.73±628.14ab 1.11±0.56b 16 591.00±5 714.33a 注:表中同一列同一处理不同字母表示各无性系根系形态参数差异显著(P<0.05)。
Note: the same column in the same treatment of different letters that the clonal root morphological parameters of significant differences(P<0.05).Table 2. Root morphological parameters of six willow clones under two treatments
-
植物光合作用是植物生长和生存最基础的过程,也是受环境影响最敏感的过程[22-23]。本研究发现,与对照相比,铜尾矿砂介质中生长的柳树叶片净光合速率(Pn)、气孔导度(Gs)以及蒸腾速率(Tr)均呈下降趋势(图 4),而大部分无性系胞间CO2浓度(Ci)则升高(图 4),且同种介质下各无性系光合参数差异显著(P<0.05),说明高重金属含量、低肥力的铜尾矿砂土壤不利于柳树的生长,导致叶片气孔导度下降,从而使叶片胞间CO2蓄积,无法进入细胞内,进而抑制了柳树的光合作用。供试6个无性系在铜尾矿砂中净光合速率(Pn)分别下降了59%(旱柳739)、73%(旱柳470)、75%(旱柳30)、49%(旱柳16)、52%(杞柳‘大红头’)、57%(杞柳‘一枝笔’)(图 4),其中,旱柳16和杞柳‘大红头’净光合速率(Pn)下降较少;然而,旱柳16在铜尾矿砂中的生物量并不理想(图 2),由此推测,尽管净光合速率(Pn)在很大程度上决定了植物干物质的积累[24],但在重金属污染环境中,生物量的积累可能还受到重金属的毒害作用及不同植物基因型的差异等因素影响。

Figure 4. Photosynthetic parameters of six willow clones under two treatment
-
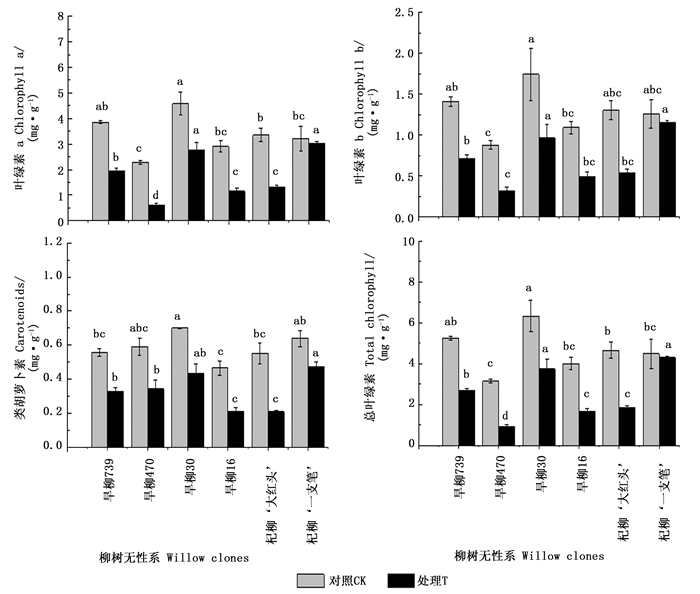
光合色素含量的高低直接影响光合作用的强弱,从而影响植株的代谢和产量。在铜尾矿砂中,6个柳树无性系叶片的平均叶绿素含量与对照相比均显著下降,叶片出现明显的黄化现象(少量叶片在叶缘出现黄化现象),而且无性系之间差异显著(P<0.05)。由图 5可看出:除杞柳‘一枝笔’外,其它5个无性系在铜尾矿砂中的叶绿素a、叶绿素b和总叶绿素含量均出现明显的下降(P<0.05),其中,下降幅度最大的是旱柳470,与对照相比,其叶绿素a、叶绿素b和总叶绿素含量分别下降了74%、64%和71%;3种色素下降幅度最小的是杞柳‘一枝笔’,下降比例仅为4%~8%。6个无性系的类胡萝卜素含量也出现明显的下降(图 5),铜尾矿砂对类胡萝卜素含量的抑制率从大到小依次为:杞柳‘大红头’>旱柳16>旱柳470>旱柳739>旱柳30>杞柳‘一枝笔’。

Figure 5. Photosynthetic pigment contents of six willow clones under two treatments
-
铜尾矿砂介质下柳树无性系不同指标主成分分析(表 3)表明:在所有的主成分构成中,信息主要集中在前3个主成分,其累积贡献率为83.778%,其中第1主成分贡献率为44.372%,第2主成分、第3主成分的贡献率分别为21.974%和17.432%。根据特征根的生物学信息,第1主成分主要是生长性状因子,在第1主成分的特征向量中,特征向量值较高且为正的性状有根体积、茎干质量、叶干质量、总生物量4个因子,说明这4个因子是决定铜尾矿砂中柳树生长状况的关键因子,因此,在进行植被恢复过程中,应选择根系庞大、地上部生长良好的无性系。第2主成分主要是光合色素含量状况,其中,特征向量值较高且为正的性状有叶绿素a、叶绿素b和总叶绿素。第3主成分是光合作用性状因子,其特征向量值较高且为正的性状有胞间CO2浓度。以上3个主成分分析可以看出:第2主成分和第3主成分的贡献率相似,而且都低于第1主成分的贡献率,表明柳树生长性状因子在各因素中起的作用最大,而其它如光合色素含量和光合作用等因子对铜尾矿砂中生长的柳树影响较小。
主成分
Ccomponents第1主成分
The first component第2主成分
The second component第3主成分
The third component指标Index 特征向量Eigenvector 0.617 1 -0.055 7 0.583 7 根长Root length 0.895 7 -0.043 5 0.345 2 根表面积Root surface area 0.971 6 -0.016 8 0.084 6 根体积Root volume 0.065 6 0.050 2 0.523 0 根尖数Root tips 0.890 1 0.088 1 -0.314 8 株高Height 0.907 0 0.094 1 -0.335 8 地茎Ground diameter 0.870 7 0.223 7 -0.341 6 根干质量Root dry weight 0.942 4 0.209 9 -0.095 7 叶干质量Leaf dry weight 0.943 6 -0.058 8 -0.313 2 茎干质量Stem dry weight 0.942 3 0.070 7 -0.301 0 总生物量Total biomass -0.079 9 0.915 2 0.326 9 叶绿素a Chlorophyll a -0.067 8 0.914 1 0.305 3 叶绿素b Chlorophyll b -0.063 8 0.928 3 0.312 6 总叶绿素Total chlorophyll 0.505 0 0.710 8 0.236 9 类胡萝卜素Carotenoids -0.339 9 0.472 2 -0.743 2 净光合速率Net photosynthetic rate -0.510 9 0.492 3 -0.531 5 气孔导度Stomatal conductance 0.081 5 -0.087 4 0.764 1 细胞间CO2浓度Intercellular CO2 concentration -0.403 4 0.561 5 -0.313 2 蒸腾速率Transpiration rate 特征根Eigenvalue 7.987 3.955 3.138 贡献率Contribution rate/% 44.372 21.974 17.432 累积贡献率Accumulative contribution rate/% 44.372 66.346 83.778 因子Factors 生长Growth 光合色素
Photosynthetic pigment/%光合作用
PhotosynthesisTable 3. Principal component analysis of different indicators under copper tailing
对铜尾矿砂介质下的不同指标进行归一化处理并计算得分,6个柳树无性系,得分大小依次为:杞柳‘大红头’(4993)>杞柳‘一枝笔’(4169)>旱柳470(2706)>旱柳16(2441)>旱柳30(2325)>旱柳739(1266)。
2.1. 铜尾矿砂对6个柳树无性系生长和根系发育的影响
2.2. 铜尾矿砂对柳树叶片光合参数的影响

2.3. 铜尾矿砂对柳树叶片光合色素的影响
2.4. 铜尾矿砂中柳树无性系生长性状的主成分分析
-
当土壤重金属浓度达到影响植物生长的临界值时,植物生长就会受到影响,最明显的症状是苗高和叶色的变化,最终反映在植物地上部生物量的显著降低[25]。Macfarlane等[26]研究发现,高浓度的Zn(500 μg·g-1)、Pb(400 μg·g-1)培养时,植株的死亡率增加,生物量降低。刘治昆等[27]研究表明,‘苏柳172’(S. jiangsuensis CL J-172)和垂柳(S. babylonica Linn)受Cu2+胁迫时生物量随浓度升高而减小。本研究也发现,供试6个柳树无性系在铜尾矿砂中生长4个月后,苗高均受到不同程度的抑制,叶色出现深浅不一的黄色,根部、地上部生物量均出现明显的下降,这与大多数研究结果一致[6, 8]。Yang等[28]研究发现,相同浓度的Cu处理下,不同基因型柳树生长表现不同,有些基因型的生长受到促进作用,而有些受到抑制作用,说明柳树对重金属污染响应的基因型差异是普遍存在的。本研究发现,无论是对照组还是处理组,在相同的培养条件下,供试6个柳树无性系生物量均存在明显的基因型差异,其中,旱柳470、旱柳739以及杞柳‘一枝笔’等无性系具有较高的生物量积累。在逆境环境中,植物的生长状况最终体现在生物量方面[29],主成分分析也表明,贡献率最大的第1主成分中总生物量具有较高的因子载荷,因此,在进行重金属污染尾矿植被恢复过程中,应选择具有较高生物量的无性系材料。
研究表明,在重金属污染的土壤中,根系越庞大,向周围空间伸展的能力越强,越有利于扩大营养吸收的范围[29-30],因此,那些根系庞大、细根较多的植物在重金属污染土壤中的竞争力越强。Manciulea等[31]通过对Cd胁迫下印度芥菜(Brassica juncea L.)和莴苣(Lactuca sativa L.)对Cd吸收的差异研究发现,印度芥菜茎部Cd的含量远远大于莴苣,主要原因可能是印度芥菜具有庞大的根系系统。Brunner等[34]研究发现,林木细根积累的重金属最高可达土壤重金属总量的0.2%,对重金属污染的植物固定化修复具有重要意义。本研究主成分分析表明,在贡献率最高的第1主成分中,除了与生长相关的株高、地径、生物量等因子以外,根体积和根表面积也具有较高的因子载荷,说明在铜尾矿砂中,根系的生长发育对柳树的生长也起着关键作用。尽管各无性系在铜尾矿砂中根系的生长发育均受到明显的抑制作用,但根据对不同无性系根系形态的观察,杞柳在铜尾矿砂中根系的发育总体上优于旱柳。对不同无性系耐性指数的计算也发现,杞柳的耐性指数达49%~50%,而旱柳4个无性系的耐性指数仅为28.93%~40.28%,特别是杞柳‘一枝笔’,不仅具有较高的耐性指数,而且也具有较高的生物量积累,因此,其可望用于铜尾矿废弃地的植被恢复。
光合作用是所有绿色植物生长和生命活动的基础,而逆境条件下光合系统的可塑性对提高植物的生存能力和生长潜力具有重要意义,因此,基于树木光合速率的选育也是寻找理想的植物修复材料的重要过程[8-9]。本研究发现,不同柳树无性系净光合速率(Pn)存在明显的基因型差异,其中,旱柳739、旱柳16以及杞柳‘一枝笔’等无性系在各处理均具有较高的净光合速率(Pn),各无性系在重金属污染条件下的净光合速率(Pn)均出现明显的下降,说明重金属污染抑制了柳树的光合作用。Pajevic′等[8]在研究白柳(S. alba L.)无性系对复合重金属污染的响应时,也发现重金属污染严重抑制了白柳的光合作用; 然而,也有研究发现[5],重金属污染条件下,蒿柳(S. viminalis L.)在重金属污染环境中的净光合速率并没有降低,反而增加,从而使其生物量变化不大,这也说明重金属污染对柳树光合作用的影响是双向的,而出现这种双向调节的原因,一方面可能是由于重金属污染程度不同,另一方面可能是由于不同基因型之间的内在遗传机制造成的[32]。
重金属污染对植物光合作用的影响主要是由于重金属影响了植物光合色素的生物合成、电子传递以及二氧化碳还原、气孔开合等过程[9]。本研究也发现,铜尾矿砂介质导致不同柳树无性系气孔导度(Gs)下降,而且下降程度存在明显的基因型差异(P<0.05),其中,对杞柳‘大红头’气孔导度的影响最小,在铜尾矿砂中生长的植株气孔导度与对照中生长的植株几乎相同;同时,对蒸腾速率(Tr)的研究也发现了类似的规律。叶绿素含量降低是重金属对植物毒害的普遍现象,刘春生等[33]研究发现,过量铜可极显著地降低苹果(Malus pumila Mill.)叶片叶绿素含量。本研究也发现,铜尾矿砂引起了不同基因型柳树叶绿素的下降,其中,杞柳‘一枝笔’叶绿素含量下降程度最小,另外,旱柳739的叶绿素含量变化也不明显。由此说明,重金属污染不仅导致了柳树叶片气孔的部分关闭,限制了进入叶肉细胞的二氧化碳,而且影响了柳树叶绿素的合成,使叶绿素含量下降,由此导致净光合速率(Pn)不同程度的下降; 然而,逆境下气孔的关闭除了抑制光合作用以外,对减少植株的蒸腾作用也具有重要意义,而这一点,对于减少逆境下植物体内的水分流失同样重要。
-
6个柳树无性系在铜尾矿砂中的生长和生物量积累具有明显的基因型差异,根据对生物量、耐性指数以及光合作用等生理生化相关指标的综合分析,杞柳‘一支笔’和‘大红头’在铜尾矿砂中的耐性高于旱柳无性系,光合色素的合成受影响较小,而且也具有较好的生物量积累能力,在铜尾矿的植被恢复过程中是值得推荐的品种;但由于本研究未考虑不同无性系对铜尾矿重金属的积累能力,因此,杞柳无性系对铜尾矿砂中重金属的提取能力有待于进一步的研究。
 DownLoad:
DownLoad: