




-
研究森林群落种群的结构和动态对阐明森林的形成与维持、种群的个体配置情况、种群数量动态以及群落的发展趋势具有重要意义[1-4]。以往针对群落特征及种群结构的研究主要关注珍稀濒危物种[5-7]。狭域特有种是分布范围相对狭窄的物种,一般由于对生境要求较严格,只能生长于山区或者极端特殊生境区域[8]。部分狭域特有种被列入中国物种红色名录,如金沙江下游干热河谷局部区域的狭域特有植物毛蕊三角车(Rinorea erianthera C. Y. Wu et C. Ho)、石灰岩狭域特有植物报春苣苔(Primulina tabacum Hance)、东阿拉善-西鄂尔多斯狭域特有种四合木(Tetraena mongolica Maxim)等[9]。随着人类活动的加剧、生境改变以及自身遗传限制等因素的影响,狭域特有种尤其是未被引起足够关注的物种的野生种群面临着严重的威胁,对其进行群落学和种群学的研究并为其制定相应的保护和保育策略是非常紧迫的。
元江蚬木(Burretiodendron kydiifolium Y. C. Hsu & R. Zhuge),又名元江柄翅果,椴树科(Tiliaceae)柄翅果属(Burretiodendron Rehd.)落叶乔木,是云南元江干热河谷局部区域的狭域特有植物[10]。元江蚬木是元江干热河谷的土著树种,具有耐旱性和对干热河谷较强的适应能力,该物种木材坚硬,可作为用材树种,具有重要的保护价值、利用价值和研究价值[11]。以往除唐亚等学者对元江蚬木的系统学地位进行过探讨外[12-14],有关元江蚬木群落特征及种群结构等保护生物学方面的研究未见报道。本研究基于元江蚬木野外生存现状的实地调查,分析该物种的群落特征及种群结构,为该物种的生物多样性保护以及干热河谷狭域特有种的可持续发展提供科学依据。
HTML
-
研究区域位于云南省元江哈尼族彝族傣族自治县(101°39′~102°21′ E,23°18′ ~23°54′ N)的元江干热河谷,这是我国西南典型的干热河谷[15]。由于西南方向的哀牢山及无量山阻挡来自孟加拉湾的西南暖湿气流,形成了突出的河谷焚风效应,气候十分干热,元江河谷(海拔400 m)的平均温度23.9℃,极端高温42.6℃,年均降水量748.7 mm,年均蒸发量2 754.8 mm,干湿季分明,降水集中,旱季长达8个月,为半干旱暖冬高原季风气候[16-17]。土壤主要是在石灰岩和砂岩上发育的燥红壤[16]。海拔800 m以下地区的植被类型为干热河谷稀树灌木草丛、灌草丛及肉质多刺灌丛,局部沟谷残存季雨林[16, 18-19]。
-
通过查阅文献及访问调查,对元江干热河谷区域元江蚬木集中分布地段采用典型抽样法,于2014年4月、11月和2015年1月、4月建立了5块20 m×20 m的固定样地和4条累计长度为8.95 km的调查样线,记录各样地及样线两侧20 m范围内出现的元江蚬木植株的GPS坐标,并记录生境特征及人为干扰情况。分层调查了样地内所有维管植物,记录木本植物的高度、胸径,同时记录样线上元江蚬木植株的高度、胸径(表 1,表 2)。
样地
Sample plots地点
Sites海拔
Altitude/m坡向
Slope坡位
Slope position坡度
Slopes/°岩石类型
Rock type土壤类型
Soil type人为影响
Anthropogenic disturbance1 普漂 595 西偏南30° 中下部 25 石灰岩裸露25% 燥红壤 放牧 2 普漂 595 西偏南30° 中下部 15 石灰岩裸露30% 燥红壤 放牧 3 元洼路 629 西坡 中部 30 砂岩 燥红壤 放牧、砍伐 4 元洼路 790 西北坡 中部 35 砂岩 燥红壤 放牧、砍伐 5 白沟深 454 南坡 中下部 25 砂岩 燥红壤 放牧、砍伐 Table 1. Basic information of the samples plots
样线
Sample lines地点
Sites长度
Distance
/km岩石类型
Rock type土壤类型
Soil type人为影响
Anthropogenic
disturbance1 普漂 2.14 石灰岩 燥红壤 放牧 2 元洼路 4.35 石灰岩 燥红壤 放牧、砍伐 3 鲁业冲 1.57 石灰岩 燥红壤 放牧、砍伐 4 施坻冲 0.89 / 燥红壤 放牧、砍伐 Table 2. Basic information of the sample lines
-
按照丹麦学者Raunkiaer的生活型分类系统统计并绘制生活型谱[20]。
-
分别计算乔木层、灌木层和草本层的重要值[21],重要值=(相对频度+相对显著度+相对密度)/3,其中:相对频度=某个种的频度/所有种频度之和×100;相对显著度=某个种的胸高断面积/样地中全部个体胸高断面积总和×100;相对密度=某个种的个体数/全部植物的个体数×100。
乔木层的相对显著度用胸高断面积进行计算;由于灌木层和草本层的物种无法计算胸高断面积,用盖度来计算物种的显著度。
-
元江蚬木数量稀少又无解析木资料,采用“空间代替时间”的方法,用大小级结构分析法代替年龄结构分析元江蚬木种群结构特征。许多学者提出过不同的划分标准[22-25],对元江蚬木种群大小级结构按以下标准进行划分:Ⅰ幼苗(H≤150 cm);Ⅱ幼树(H>150 cm,DBH < 5 cm),采用上限排外法,以后各级胸径(DBH)每隔5 cm为1级,Ⅲ(5 cm≤DBH < 10 cm);Ⅳ(10 cm≤DBH < 15 cm);Ⅴ(15 cm≤DBH < 20 cm);Ⅵ(DBH≥20 cm)。
-
根据干扰类型以及调查地点种群受到干扰的威胁状况,将干扰强度分为轻度、中度和重度,详细的划分原则见表 3。
干扰强度
Disturbance intensity样地/样线
Sample plots/lines干扰类型及受威胁状况
Disturbance type and threatened conditions轻度 样地1、样地2、样线1 放牧,位于元江国家级自然保护区 中度 样地5 放牧、砍伐 重度 样地3、样地4、样线2、样线3、样线4 放牧、砍伐,位于道路两侧 Table 3. Definition and description of disturbance intensity
1.1. 研究区域概况
1.2. 野外调查
1.3. 分析方法
1.3.1. 生活型
1.3.2. 重要值计算
1.3.3. 种群大小级划分
1.3.4. 干扰强度的划分
-
在总面积为2 000 m2的5个调查样地中,记录到维管束植物共计83种,隶属于39科69属,其中,蕨类植物1种,属于卷柏科卷柏属;被子植物38科68属82种,其中双子叶植物29科51属64种,单子叶植物9科17属18种。各样地维管束植物在22~35之间,平均为28.6。
-
元江蚬木群落生活型谱(图 1)可以反应该群落的环境特征,高位芽植物占优势,有50种,所占比例高达60.24%,又以矮高位芽为主,小高位芽次之,比例分别为28.92%、19.28%,藤本高位芽比例为12.04%,缺乏大高位芽和中高位芽植物,元江蚬木群落以高位芽植物占绝大多数,反应了该群落植物生长季节具有温热多湿的环境特征;地上芽植物居第二,有16种,比例为19.28%,反应了该群落植物生长季节具有一定的年均降雨量较小,极端最低温度较高的环境特征[26];地面芽植物和地下芽植物分别有10种和6种,比例为12.05%、7.23%;一年生植物占总物种比例的1.2%,只有鸭跖草(Commelina communis L.)一种。

Figure 1. Life form spectrum of Burretiodendron kydiifolium community
-
元江蚬木所处群落为季雨林(植被型)中的落叶季雨林(植被亚型),季雨林的外貌由于上层树种多数由落叶、半落叶的树种组成,因此具有明显的季相变化,而落叶季雨林的季节变化更为突出,该类型主要由落叶树种组成,群落结构简单,优势种比较显著,并具有许多喜阳耐旱的生态适应反映[15, 27]。根据主要种类的不同,元江蚬木群落分为3个群系:元江蚬木+厚皮树林(Form. Burretiodendron kydiifolium +Lannea coromandelica)、元江蚬木+清香木林(Form. Burretiodendron kydiifolium+Pistacia weinmannifolia)以及元江蚬木+白皮乌口树林(Form. Burretiodendron kydiifolium+Tarenna depauperata)。
元江蚬木群落乔、灌木树种有55种,以落叶树为主,比例高达90.9%,常绿物种只有5种,包括:直脉榕(Ficus orthoneura Levl. et Vant.)、清香木(Pistacia weinmannifolia J. Poisson ex Franch.)、厚皮香(Ternstroemia gymnanthera (Wight et Arn.) Beddome)、九里香(Murraya exotica L.)、霸王鞭(Euphorbia royleana Boiss.)。
群落总盖度60%~90%,分为乔木层、灌木层、草本层和层间层(表 4)。乔木层盖度0~50%,植物种类19种,个体数量达108株,高度5~14 m,胸径4.8~32 cm,包括元江蚬木、厚皮树(Lannea coromandelica (Houtt.) Merr.)、异序乌桕(Sapium insigne (Royle) Benth. ex Hook. f.)、清香木、蒙自合欢(Albizia bracteata Dunn)、直脉榕、白头树(Garuga forrestii W. W. Smith)、金合欢(Acacia farnesiana (L.) Willd.)、槐(Sophora japonica L.)、老人皮(Polyalthia cerasoides Roxb)、心叶木(Haldina cordifolia (Roxb.) Ridsd.)等。
物种Species 相对频度
Relative frequency相对显著度
Relative significance相对密度
Relative density重要值
Important value乔木层 元江蚬木Burretiodendron kydiifolium Y. C. Hsu & R. Zhuge 16 45.34 50.00 37.11 厚皮树Lannea coromandelica (Houtt.) Merr. 8 21.45 14.81 14.76 异序乌桕Sapium insigne (Royle) Benth. ex Hook. f. 8 6.73 7.31 7.38 清香木Pistacia weinmannifolia J. Poisson ex Franch. 8 6.17 3.70 5.96 蒙自合欢Albizia bracteata Dunn 8 4.62 2.78 5.13 灌木层 元江蚬木Burretiodendron kydiifolium Y. C. Hsu & R. Zhuge 5.62 35.15 33.24 24.67 霸王鞭Euphorbia royleana Boiss. 4.49 9.77 16.71 10.33 疏序黄荆Vitex negundo L. 5.62 8.35 9.25 7.74 白皮乌口树Tarenna depauperata Hutchins. 2.25 5.67 5.14 4.35 元江素馨Jasminum yuanjiangense P. Y. Bai 3.37 5.71 3.21 4.10 草本层 卷柏Selaginella pulvinata (Hook. et Grev.) Maxim. 18.18 32.88 16.77 22.61 扭黄茅Heteropogon contortus (L.) Beauv. ex Roem. et Schult. 4.55 14.95 26.69 15.39 鸭趾草Commelina communis L. 9.09 10.46 5.34 8.30 沿阶草Ophiopogon bodinieri Levl 9.09 7.77 6.94 7.93 芸香草Cymbopogon distans (Nees ex Steud.) Wats. 4.55 7.47 7.62 6.55 层间层 土密藤Bridelia stipularis (L.) Bl. 16.67 30.30 55.87 34.28 翅果藤Myriopteron extensum (Wight) K. Schum 11.11 31.32 18.25 20.39 云南牛奶菜Marsdenia balansae Cost. 11.11 13.64 12.85 12.53 昆明马兜铃Aristolochia kunmingensis C. Y. Chen et J. S. Ma 5.56 7.58 6.21 6.45 虫豆Cajanus crassus (Prain ex King) Van der Maesen 11.11 4.55 1.12 5.59 Table 4. The top five species of important values in Burretiodendron kydiifolium community
灌木层盖度30%~50%,高0.2~7.0 m,植物种类47种,包括乔木幼树及灌木。乔木幼树22种,比例达46.81%;灌木25种,比例53.19%。乔木幼树主要有元江蚬木、厚皮树、老人皮、朴树(Celtis sinensis Pers.)、直脉榕、清香木、红椿(Toona ciliate Roem.)、刺桑(Streblus ilicifolius (Vidal) Corner)、槐等;灌木有霸王鞭、疏序黄荆(Vitex negundo L.)、白皮乌口树(Tarenna depauperata Hutchins.)、元江素馨(Jasminum yuanjiangense P. Y. Bai)、一叶萩(Flueggea suffruticosa (Pallas) Baillon)、瘤果三宝木(Trigonostemon tuberculatus F. Du et Ju He)、假杜鹃(Barleria cristata L.)、越南叶下珠(Phyllanthus cochinchinensis (Lour.) Spreng.)、细梗美登木(Maytenus graciliramula S. J. Pei et Y. H. Li)等。
草本层盖度5%~10%,主要有卷柏(Selaginella pulvinata (Hook. et Grev.)Maxim.)、扭黄茅(Heteropogon contortus (L.) Beauv. ex Roem. et Schult.)、鸭趾草、沿阶草(Ophiopogon bodinieri Levl)、芸香草(Cymbopogon distans (Nees ex Steud.) Wats.)、细柄草(Capillipedium parviflorum (R. Br.) Stapf)、铃铛草(Themeda caudata (Nees) A. Camus)、线纹香茶菜(Rabdosia lophanthoides (Buch.-Ham. ex D. Don) Hara)、飞机草(Chromolaena odorata (L.) R. M. King & H. Rob.)等。
层间植物以藤本植物为主,种类有土密藤(Bridelia stipularis (L.) Bl.)、翅果藤(Myriopteron extensum (Wight) K. Schum)、云南牛奶菜(Marsdenia balansae Cost.)、昆明马兜铃(Aristolochia kunmingensis C. Y. Chen et J. S. Ma)、虫豆(Cajanus crassus (Prain ex King) Van der Maesen)、翅子藤(Loeseneriella merrilliana A. C. Smith)、叉蕊薯蓣(Dioscorea collettii Hook. f.)等。
-
元江蚬木分布最低海拔454 m,最高海拔812 m,呈零星或片断分布,斑块面积非常小,生境破碎化十分严重。调查记录到元江蚬木412株,其中乔木层234株,高度范围5~25 m,胸径3~45 cm;灌木层178株,高度0.05~4.5 m,最大胸径达30 cm(表 5)。分布区内元江蚬木的平均密度约22.76株·hm-2。
样地/样线
Sample plots
/lines所处层次
Layers数量
Numbers高度
Height
/m胸径
DBH/cm海拔
Altitude
/m样地1 乔木层 10 5~10 4.9~29.15 595 灌木层 2 1.2~3.5 0.4~3.5 样地2 乔木层 11 5~9 7~13.3 595 灌木层 7 0.05~3.5 ≤5.8 样地3 乔木层 16 5~14 4.3~24.5 629 灌木层 23 0.7~4.5 1.2~5.7 样地4 乔木层 15 5~15 3~25 796 灌木层 50 0.3~4 ≤11.5 样地5 乔木层 0 / / 454 灌木层 29 0.4~4.5 ≤6.2 样线1 乔木层 88 5~15 5~25.8 519~623 灌木层 36 0.1~4.5 ≤10 样线2 乔木层 36 5~12 6~30 603~812 灌木层 9 2~4.5 230 样线3 乔木层 23 6~25 8~45 596~727 灌木层 4 2.5~4 ≤10 样线4 乔木层 35 5~15 6~25 579~681 灌木层 18 2.5~4.5 3~22 合计 乔木层 234 5~25 3~45 454~812 灌木层 178 0.05~4.5 ≤30 Table 5. The number of Burretiodendron kydiifolium individuals in sample plots/lines
-
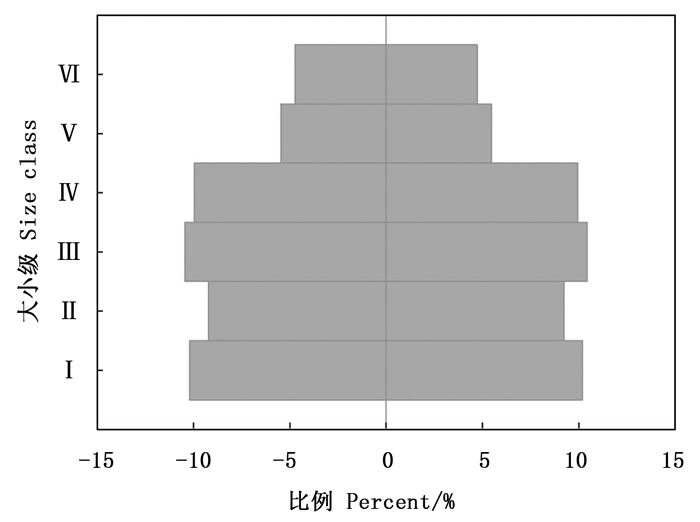
元江蚬木种群Ⅵ级木(20 cm≤DBH)数量最少,占总株数的9.47%,其次是Ⅴ级(15 cm≤DBH < 20 cm),占总株数的10.92%。Ⅰ、Ⅱ、Ⅲ、Ⅳ级数量占总株数的比例差异较小,分别为20.39%、18.45%、20.87%以及19.9%,种群结构整体呈稳定型(图 2)。

Figure 2. Population structure of Burretiodendron kydiifolium
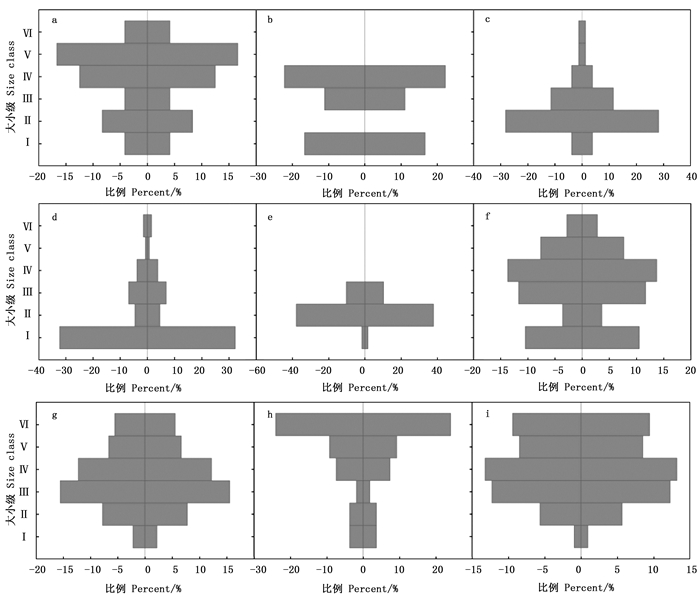
对比各样地和样线元江蚬木种群结构(图 3),发现样地2和样线3元江蚬木种群结构呈现衰退型(图b、h),样地1、样线1、样线2和样线4呈稳定型(图a、f、g、i),而样地3、样地4、样地5种群结构呈增长型(图c、d、e)。

Figure 3. Population structure of Burretiodendron kydiifolium in sample plots/lines
-
野外调查发现由于历史干扰因素的影响,元江蚬木种群含有一定的萌枝,数量38株,比例9.22%,萌枝出现在样地5、样线2和样线4(表 6),样地5和样线4的萌枝处于Ⅰ级幼苗和Ⅱ级幼树的比例分别为75.86%和1.89%。可见,除实生苗外,枝条萌生也是元江蚬木种群更新的方式。
样地/样线
Sample plots/lines萌枝比例
Percent of sprout
branches/%Ⅰ级幼苗和Ⅱ级幼树的比例
Percent of seedlings
and young trees/%样地5 96.55 75.86 样线2 8.89 0 样线4 16.98 1.89 Table 6. The regeneration of sprout branches of Burretiodendron kydiifolium
对比不同干扰程度下各样地和样线Ⅰ级幼苗和Ⅱ级幼树的数量(图 4),发现不同干扰强度下不同地点元江蚬木种群的更新情况存在较大差异。干扰程度是影响种群更新的原因之一,处于轻度干扰的样线1幼苗和幼树分别为26株、9株(f);样线2、样线3和样线4受到重度干扰的影响(g、h、i),幼苗和幼树数量较少,均在7株以下,最少只记录到1株。而样地1和样地2虽然受到轻度的干扰(a、b),但是由于样地岩石裸露,生境极其干燥,不利于种子萌发和幼苗的生长;处于中度干扰的样地5由于受到放牧和砍伐的影响(e),元江蚬木群落具有明显的次生性质,植株多呈灌木状,高度均在5 m以下,虽然幼树个体数量较多,但多是萌生枝条,实生幼苗只有1株;样地3和样地4虽然受到重度干扰的影响(c、d),但生境相对湿润,样地3成年树较少,保留了更多幼树的生长;而样地4群落郁闭度大,有利于种子萌发和幼苗生长。

Figure 4. The difference of the number of seedlings and young trees of Burretiodendron kydiifolium at different disturbance intensity levels in sample plots/lines
2.1. 群落特征
2.1.1. 物种组成
2.1.2. 生活型
2.1.3. 群落外貌特征
2.2. 种群结构
2.2.1. 种群数量
2.2.2. 种群结构
2.2.3. 更新状况

-
元江蚬木分布范围狭窄,为元江干热河谷狭域特有植物,是云南省北热带干热河谷地区原生季雨林的建群种,也是元江河谷植被恢复中的重要标志树种。野外调查时间是1月、4月、11月,基本处于旱季,部分1年生草本植物处于枯死的状态,可能会导致草本种类记录不完整。详细的群落研究还需增加不同季节的野外调查。
元江蚬木种群整体结构呈现稳定型,但不同分布地点元江蚬木种群存在增长型、稳定型和衰退型3种结构,其中样地2和样线3种群结构呈衰退型。虽然样地1和样地2均受到轻度干扰和生境干旱的双重影响,而且Ⅰ级幼苗和Ⅱ级幼树的数量均在6株以下,但样地2存在大小级缺失的现象,未记录到Ⅱ级幼树、Ⅴ级和Ⅵ级的个体,且Ⅳ级个体数量显著大于Ⅰ级幼苗,种群结构呈现出衰退型。样线3 Ⅵ级个体数量13株,比例达48.15%,并记录到整个种群胸径最大的一株,达45 cm,由于受到重度的人为干扰的影响,幼苗和幼树数量均在2株以下,种群结构呈典型的衰退型。
元江蚬木种群更新方式主要包括实生苗和萌枝,萌枝是木本植物受到自然或人为干扰以及剧烈环境变化而做出的响应[28],这种更新策略在许多物种的种群研究中也同样存在[29-30]。样地5元江蚬木种群结构中的部分大小级缺失,只有Ⅰ幼苗、Ⅱ幼树和Ⅲ级,且幼树萌枝占整个样地元江蚬木种群数量的75.86%,种群结构呈增长型。不同学者对萌枝的方式划分存在差异,其中Del Tredici[31]将温带落叶树种萌枝的方式划分为根茎萌枝、木质瘤萌枝、根出条萌枝和机会萌枝,而张松等[32]认为除了以上的萌枝方式,还应增加树冠萌枝和残桩萌枝。有研究发现萌枝萌发的位置对将来物种的发展状况具有决定作用,如:残桩的萌枝生长过程中所需要的营养、水分主要依赖于主干,容易受到病虫害的侵扰而随着主干的衰退而逐渐衰退[33],若萌生枝是从地面或者地面以下萌发,其基部可以直接与土壤接触,而有利于植株的生长发育[34]。有关元江蚬木的种群研究还需进一步确认其萌枝的方式,以此更加准确的分析未来种群的发展动态。
元江蚬木种群更新受到生境特征和人为干扰的影响,生境干旱及重度的人为干扰都不利于种子萌发和幼苗、幼树的生长。生境状况对植物生活史具有重要作用,对植物数量、存活和更新具有直接影响[35-37],同时,野外观测到元江蚬木结实率很高,抽样统计样地1和样地2的种子数量分别为14 856粒、10 411粒,而实生苗数量很少,可能与元江蚬木自身生物学特性有关,具体原因有待进一步研究。
位于元江县普漂的元江蚬木种群处于元江国家级自然保护区的核心区,而其他分布点,都不在保护区内,元江蚬木群落均受到不同程度的人类活动的干扰。要加强对元江蚬木的保护,关键是杜绝人为破坏;以就地保护为主,保护原生生境,促进天然更新;同时通过人工采种繁育幼苗,定植到原生境,复壮野外种群;此外,还需加强对种子到幼苗动态变化的研究,探索种群更新的内在影响机制。
-
根据野外调查及文献资料记录,元江蚬木属于典型的元江干热河谷局部地段的狭域特有种,是云南省北热带干热河谷地区原生季雨林的建群种,呈零星或小片断状分布。元江蚬木群落维管束植物83种,隶属39科69属,群落季相变化明显,落叶树种较多,生活型以高位芽植物为主,其次是地上芽植物,植被类型为季雨林中的落叶季雨林,群系划分为元江蚬木-厚皮树林、元江蚬木-清香木林以及元江蚬木-白皮乌口树林。元江蚬木种群整体结构呈稳定型,但是,增长型、稳定型和衰退型结构在不同分布地点均存在。实生苗和萌枝是元江蚬木种群更新的主要方式,不同分布地点幼苗和幼树数量差异较大,生境干旱和人为干扰是影响种群更新的主要原因。
 DownLoad:
DownLoad: