



-
树冠是森林进行气体交换最主要和最直接的部分[1],因此,冠层光合生理参数的变化对森林生产力形成具有重要影响。树木不同冠层及方位太阳辐射[2-5]、水分蒸腾[6-9]、比叶面积[7, 10]、叶绿素含量[11-12]、营养元素含量[13-14]的差异都将导致冠层光合参数的空间异质性。前人对于光合参数的研究大多针对幼苗或冠层某一部位,而这些研究很难反映整个冠层的光合特性,以这些测定数据来估算冠层生产力时可能会产生较大的误差。因此,认知大树冠层光合参数的空间变化规律及其影响因素有助于准确估算冠层生产力。
落叶松(Larix spp.)天然分布于温带山区、寒温带平原及高山气候区,枝条有长短枝之分,叶倒披针状线型,在长枝上螺旋状散生,在短枝上簇生。此外,落叶松具有适应性强、早期速生、成林快、病虫害少、材质优良的特点,是我国东北、西北、华北及南方亚高山地区的重要纸浆材及建筑材树种,也是退耕还林及防护林工程的主要造林树种[15]。国内许晨璐等[16]、刘宁等[17]、毛子军等[18]和郭盛磊等[19]分别研究了落叶松种间光合特性的差异、光照变化、氮(N)及CO2增加对光合特性的影响,但这些研究均是针对幼苗或幼树的某一部位进行,很少涉及大树不同冠层和方位光合作用的研究[20-21]。因此,本研究以7年生和19年生日本落叶松(Larix kaempferi (Lamb.) Carr.)为研究对象,定量研究不同冠层和方位的光合生理参数差异,揭示冠层光合特性的空间变化规律,旨在为精确估算冠层水平生产力及进一步构建日本落叶松冠层生产力模型提供参考。
HTML
-
研究地位于甘肃省小陇山林业实验局林业科学研究所沙坝实验基地(105°42′~106°00′E,34°07′~34°24′N),地处秦岭西段北坡,海拔1 500~2 100 m。该地位于我国暖温带南缘向北亚热带过渡的地带,属暖温带湿润气候区,年均气温7.2℃,≥10℃年均积温2 480℃,年均降水量757 mm,年均蒸发量1 012 mm,年均相对湿度78%,年均日照时数1 553 h,全年无霜期120~218 d。
研究样地分别设置在沙坝实验基地164和168林班,林龄分别为7、19 a,在每个林分中设置3块20 m×30 m的样地,对样地内树木进行每木检尺,并从中选择平均木作为测定样木。落叶松样地概况见表 1。
林龄
Stand age/a胸径
DBH/cm树高
Height/m林分密度
Stand density/(tree·hm-2)坡向
Slope orientation坡度
Slope/(°)海拔
Elevation/m土壤类型
Soil type7 6.9 7.4 3 700 西南 31 1 687 褐色森林土 19 13.5 16.1 2 160 西 20 1 712 褐色森林土 Table 1. Survey of Larix kaempferi plantation plots
-
在样木四周利用钢管搭设与样木等高的架子作为观测平台,根据冠层外缘针叶接受光照的差异和树冠高度,将树冠等分为上、中、下3层,利用罗盘将每一冠层分为东、西、南、北4个方位。在上、中、下冠层的每一方位均选择靠近冠层外缘的非当年生枝条作为测定样本,在枝条上选择相邻的1~2簇针叶,使用便携式光合作用测定系统(Li-6400XT;Li-Cor Inc.,Lincoln,NE,USA)连接可控光簇状叶室(6400-22L)测定所选针叶的光响应曲线与CO2响应曲线。在非阴雨天对样枝光合响应曲线进行测定,每月对各样枝均测定1次,测定日期分别为2014年7月22—25日和8月23—26日,测定时间为8:30—12:30。为保证不同部位针叶光合响应参数的可比性,在响应曲线测定前,将叶室内光照强度设置为1 000 μmol·m-2·s-1,CO2浓度为380 μmol·mol-1,温度为25℃,流速设置为500 μmol·s-1,对测定针叶进行30 min诱导。
-
调用光合测定系统中的自动测定程序进行光响应曲线和CO2响应曲线的测定,在响应曲线自动测定程序中,稳定时间设置为180 s。在测定光响应曲线时,CO2浓度设为380 μmol·mol-1,温度设为25℃,光强由强至弱设为2 000、1 700、1 300、1 000、800、600、400、200、150、100、50、20、0 μmol·m-2·s-1。在测定CO2响应曲线时,将饱和光强设为1 000 μmol·m-2·s-1,温度设为25℃,利用光合作用测定系统自带的CO2注入系统调控叶室内CO2浓度,CO2浓度梯度依次为380、250、150、60、120、200、400、600、900、1 200、1 500、1 800、2 000 μmol·mol-1。
-
采用分光光度法测定光合色素含量[22],根据针叶含水率将单位鲜质量的光合色素含量转化为单位干质量光合色素含量。
-
利用光合助手软件(Dundee Scientific,Dundee,U.K.)计算光响应参数和CO2响应参数[13],采用非直角双曲线模型[23]对光响应曲线进行拟合,估算出最大净光合速率(Amax)、表观量子效率(AQY)、光饱和点(LSP)、光补偿点(LCP)和暗呼吸速率(Rd);利用Farquar生化模型[24-25]拟合CO2响应曲线,估算出最大羧化速率(Vcmax)、最大电子传递速率(Jmax)和磷酸丙糖利用率(TPU)。将胞间CO2浓度(Ci)<200 μmol·mol-1时与其对应的净光合速率(Pn)进行直线回归[26],回归方程为:
式中:Rp为光下呼吸速率,CE为羧化效率。当Pn=0时,Ci即为CO2补偿点(CCP)。由于光下暗呼吸很小,因此,可以把光下呼吸速率近似地作为光呼吸[27]。
-
对测定数据进行正态性检验,将非正态分布数据进行自然对数转换使其呈正态分布。利用双因素方差分析比较不同冠层、方位及二者的交互作用对各光合生理参数的影响程度,当差异显著时(P < 0.05),采用S-N-K多重比较对光合生理参数进行分析,比较各光合生理参数在不同冠层和方位的差异。
2.1. 光响应曲线与CO2响应曲线的测定
2.2. 光合色素的测定
2.3. 光响应曲线和CO2曲线参数估计
2.4. 数据分析
-
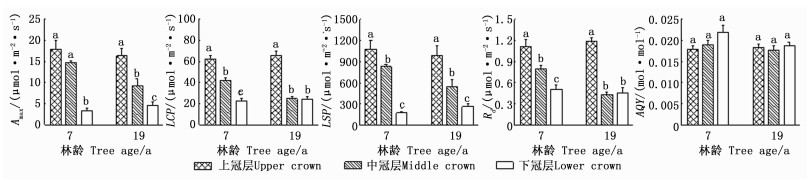
经方差分析可知:除AQY外,冠层对2林龄日本落叶松光响应参数影响显著,而方位及冠层与方位的交互作用对所有光响应参数的影响均不显著(表 2)。由图 1可知:除AQY外,光响应参数在冠层间差异显著。7年生样木的Amax、LCP、LSP和Rd随冠层的下降而减小,最大值分别为17.91、61.99、107.12、1.11 μmol·m-2·s-1,下冠层相应的光响应参数分别是上冠层的18.8%、37.0%、16.4%、45.8%;19年生样木的Amax、LCP、LSP、Rd也随冠层的下降而减小,最大值分别为16.27、65.55、993.37、1.18 μmol·m-2·s-1,下冠层相应的光响应参数分别是上冠层的28.4%、36.5%、26.9%、38.8%;7、19年生样木的AQY均表现为下冠层最大,最大值分别为0.022、0.019 mol·mol-1;这表明日本落叶松冠层间多数光响应参数具有较大的空间异质性。方差分析发现,全冠层光响应参数均值在林龄间差异不显著。
光合生理参数
Photosynthetic and physiological parameters7年生7-year-old 19年生19-year-old 冠层
Crown方位
Orientation冠层×方位
Crown×Orientation冠层
Crown方位
Orientation冠层×方位
Crown×OrientationAQY 2.40 1.46 0.50 0.2 0.15 0.71 Amax 289.18*** 2.14 1.12 11.73** 0.37 0.64 LCP 56.67*** 2.25 0.90 23.58*** 2.15 0.40 LSP 39.15*** 1.56 0.47 11.65** 0.51 0.80 Rd 17.83*** 2.63 0.61 25.84*** 2.59 0.72 Vcmax 13.79*** 0.50 1.04 15.54*** 3.09 1.15 Jmax 5.70* 0.36 0.60 2.24 0.75 0.64 TPU 2.28 0.15 0.65 0.45 0.66 0.78 CE 24.53*** 0.41 0.56 17.83*** 2.06 0.54 CCP 19.54*** 0.56 0.47 13.32*** 0.56 0.57 Chla 18.19*** 0.68 0.16 65.59*** 1.03 1.64 Chlb 14.93*** 0.20 0.12 48.43*** 0.35 1.92 Car 30.42*** 2.06 0.35 7.68** 0.19 0.20 Chl(a+b) 17.36*** 0.49 0.14 87.44*** 1.41 1.87 Chl a/b 5.84* 0.84 1.26 30.85*** 1.08 0.21 注Note:*:P < 0.05;**:P < 0.01;***:P < 0.001。AQY:表观量子效率 Apparent quantum yield;Amax:最大净光合速率 Maximum net photosynthetic rate;LCP:光补偿点 Light compensation point;LSP:光饱和点 Light saturation point;Rd:暗呼吸速率 Dark respiration rate;Vcmax:最大羧化速率 Maximum carboxylation rate;Jmax:最大电子传递速率 Maximum electron transport rate;TPU:磷酸丙糖利用率 Triose phosphate utilization;CE:羧化效率 Carboxylation efficiency;CCP:CO2补偿点 CO2 compensation point;Chla:叶绿素a Chlorophyll a;Chlb:叶绿素b Chlorophyll b;Car:类胡萝卜素 Carotenoid;Chl(a+b):叶绿素a与叶绿素b的和 Sum of chlorophyll a and b;Chl(a/b):叶绿素a与叶绿素b的比值 Ratio of chlorophyll a and b;下同 The same bellow. Table 2. The effects of canopy, orientation and their interactive effect on photosynthetic and physiological parameters of 7- and 19-year-old Larix kaempferi (F values)

Figure 1. The comparison of light response parameters in different crowns of Larix kaempferi (Mean±SE)
由图 2可看出:方位对光响应参数的影响均不显著。除LSP外,7年生样木的光响应参数均为西向最小,Amax、LCP、LSP、Rd和AQY的变幅分别为10.63~14.26、37.23~47.87、590.83~831.63、0.64~0.91 μmol·m-2·s-1和0.017~0.021 mol·mol-1;19年生样木的光响应参数均为北向最大,除AQY外,均为西向最小,Amax、LCP、LSP、Rd和AQY的变幅分别为8.38~10.99、34.08~45.81、505.17~660.08、0.62~0.84 μmol·m-2·s-1和0.018~0.019 mol·mol-1;这表明日本落叶松不同方位光响应参数存在一定程度的差异性,但差异不显著。7、19年生样木的多数光响应参数冠层均值分别与南向和东向相近,因此,可用南向和东向光响应参数分别代表 7、19年生样木全冠层均值。

Figure 2. The comparison of light response parameters in different orientations of Larix kaempferi (Mean±SE)
-
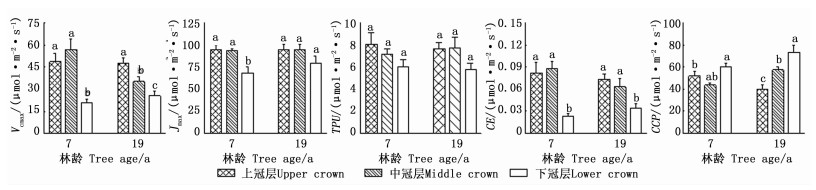
冠层对2林龄样木大多数CO2响应参数影响显著,方位及冠层与方位的交互作用对所有CO2响应参数的影响不显著(表 2)。由图 3可知:除TPU和19年生Jmax外,CO2响应参数在冠层间存在显著差异。7年生样木的Vcmax、Jmax、TPU和CE表现为上、中冠层高于下冠层,最大值分别为56.89、95.01、8.09、0.088 μmol·m-2·s-1,下冠层相应的CO2响应参数分别是最大值的37.0%、72.2%、74.8%、26.4%;19年生样木的Vcmax、Jmax、TPU和CE也表现为上、中冠层大于下冠层,最大值分别为47.45、94.97、7.76、0.074 μmol·m-2·s-1,下冠层相应的CO2响应参数分别是最大值的54.1%、83.5%、74.2%、46.9%;而7、19年生样木的CCP均表现为下冠层的最大,最大值分别为60.52、73.20 μmol·m-2·s-1。这表明,日本落叶松冠层间多数CO2响应参数也具有空间异质性。方差分析表明,全冠层CO2响应参数均值在林龄间差异不显著。

Figure 3. The comparison of CO2 response parameters in different crowns of Larix kaempferi (Mean±SE)
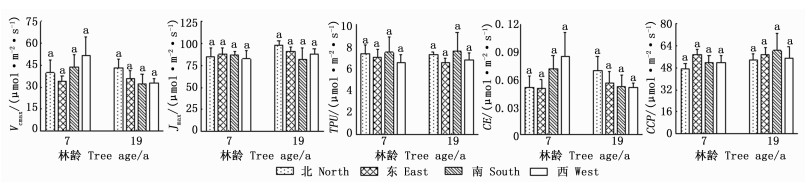
由图 4可看出:方位对CO2响应参数的影响均不显著。7年生样木的CO2响应参数无明显的变化规律,Vcmax、Jmax、TPU、CCP和CE变幅分别为33.85~51.54、82.48~87.69、6.52~7.48、47.20~57.52和0.050~0.085 μmol·m-2·s-1;19年生样木的Vcmax、Jmax、CE均表现为北向最大,TPU和CCP表现为南向最大,Vcmax、Jmax、TPU、CCP和CE变幅分别为32.39~43.27、81.53~97.99、6.54~7.62、53.71~60.57和0.051~0.070 μmol·m-2·s-1。这表明,日本落叶松方位间CO2响应参数存在差异,但差异不显著。7年生和19年生样木多数CO2响应参数冠层均值分别与南向和东向相近,因此,可用南向和东向CO2响应参数分别代表 7年生和19年生样木全冠层均值。

Figure 4. The comparison of CO2 response parameters in different orientations of Larix kaempferi (Mean±SE)
-
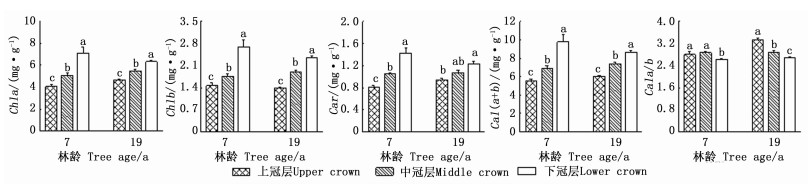
冠层对2林龄样木光合色素含量影响显著,而方位及冠层与方位的交互作用对光合色素的影响不显著(表 2)。由图 5可知:光合色素含量在冠层间存在显著差异。7年生和19年生样木的Chla、Chlb、Car、Chl(a+b)均随冠层下降而升高,7年生的最大值分别为7.09、2.70、1.43、9.79 mg·g-1,下冠层相应光合色素分别是上冠层的1.74、1.84、1.76、1.77倍;19年生的最大值分别为6.32、2.35、1.24、8.67 mg·g-1,下冠层相应光合色素分别是上冠层的1.36、1.69、1.31、1.44倍;而7年生和19年生样木Chla/b的最大值分别出现在中冠层和上冠层,分别为2.88和3.35。这说明,日本落叶松冠层光合色素含量具有显著的空间变异性。方差分析发现,全冠层光合色素含量在林龄间差异不显著。

Figure 5. The comparison of photosynthetic pigments content in different crowns of Larix kaempferi (Mean±SE)
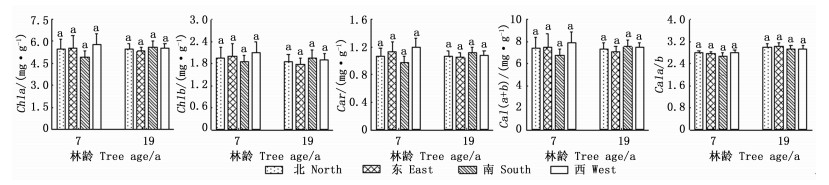
由图 6可看出:光合色素含量在方位间差异不显著。7年生样木的光合色素含量均为西向最大,南向最小,Chla、Chlb、Car、Chl(a+b)和Chla/b变幅分别为4.93~5.79、1.85~2.09、0.98~1.20、6.78~7.88 mg·g-1和2.68~2.81;除Chla/b外,19年生样木的光合色素含量均为南向最大,东向最小,Chla、Chlb、Car、Chl(a+b)和Chla/b变幅分别为5.32~5.60、1.79~1.96、1.05~1.12、7.11~7.55 mg·g-1和2.94~3.03。这表明,日本落叶松不同方位光合色素含量变化较小。

Figure 6. The comparison of photosynthetic pigments content in different orientations of Larix kaempferi (Mean±SE)
3.1. 日本落叶松光响应参数比较

3.2. 日本落叶松CO2响应参数比较
3.3. 日本落叶松光合色素含量比较
-
树木冠层微环境具有空间异质性,冠层内叶的大小、方位及分布制约着冠层内光照的变化,光照强度对树叶的发育、形态、能量平衡、水分利用效率和光合作用具有重要影响,进而导致光合参数产生垂直变化[2, 4, 26]。通过对光响应曲线的分析,有助于理解植物光合特性与光照强度的关系。本研究中,2种林龄样木的Amax、LCP、LSP和Rd均随冠层的升高而增大,而AQY均为下冠层的最大,与刘强等[28]对长白落叶松(L. olgensis Henry)的研究相一致,且Amax最大值略高于长白落叶松。这说明,日本落叶松冠层光合能力、对强光的利用能力及光合产物的消耗速率随冠层的上升而增加,对弱光的利用能力则表现为相反的变化趋势,且下冠层对弱光的转化效率高于上、中冠层,这可能是冠层针叶对所处环境长期适应的结果。因为测定样木所处林分密度较大,上冠层光照充足,下冠层则长期处于弱光照环境。光响应参数相似的变化规律在其他树种中也得到了验证[6, 21, 26, 29-30],张小全等[29]、肖文发等[6]对杉木(Cunninghamia lanceolata (Lamb.) Hook.)的研究也认为,针叶对光环境的长期适应是造成光响应参数变化的主要原因。另外,不同冠层针叶叶肉导度、气孔导度和比叶质量的改变也可能是影响Amax垂直变化的因素[2, 26]。
CO2响应曲线是分析植物利用CO2效率的有力工具,有助于理解植物叶片的生理状况及CO2浓度与净光合速率的关系[31]。本研究中,2种林龄样木上、中冠层的Vcmax、Jmax、TPU和CE均大于下冠层,而CCP为下冠层最大,表明日本落叶松上、中冠层光合能力高于下冠层;在低CO2浓度时,上、中冠层活化的核酮糖-1,5-二磷酸羧化酶/加氧酶(Rubisco)较多,光合速率较大,且对低CO2浓度的利用能力较强,具有较强的环境适应能力。Saito等[26]对圆齿水青冈(Fagus crenata Blume)及Han等[32]对赤松(Pinus densiflora Sieb. et Zucc.)冠层CO2响应参数的研究也发现了类似的规律。Han等[11, 32]对赤松的研究发现,冠层N含量与Vcmax、Jmax正相关,且N含量随冠层的下降而减少;Vcmax与比叶面积负相关,而比叶面积则随冠层的下降而升高。Qu等[33]报道了日本落叶松及其杂种落叶松(L. gmelinii (Rupr.) Kuzen. × L. kaempferi (Lamb.) Carr.)幼苗遮光12周后,其Vcmax显著低于未遮光处理。所以,不同冠层叶N含量、比叶面积与光照强度的变化是冠层CO2响应参数空间异质性的重要因素[34]。
叶绿体是植物进行光合作用的主要器官,其光合色素含量影响着植物的光合作用。本研究发现,2种林龄样木的Chla、Chlb、Chl(a+b)和Car均随冠层的升高而降低,Chla/b则表现为相反的变化趋势,表明日本落叶松下冠层针叶对太阳能的转化效率和对弱光的利用能力较高。在弱光胁迫下,日本落叶松中、下冠层的Car增加,可能会使之与更多的色素蛋白复合体结合,形成辅助色素,增加由类胡萝卜素向叶绿素能量的传递,从而维持光胁迫条件下的光合速率。相似的变化规律在刨花楠(Machilus pauhoi Kanehira)、华东润楠(M. leptophylla Hand.-Mazz.)和糖槭(Acer saccharum Marsh.)的研究中得到了验证[2, 12]。
树冠不同方位叶片在同一时间内受到的光照强度不同[4, 35],叶片对于不同光照强度的长期适应造就了其形态、解剖结构及生物化学特性的差异[3],一般来说,阳生叶的光合能力是阴生叶的24倍[4]。李六林等[36]对杂种榛(Corylus heterophylla Fisch. × C. avellana L.)的研究结果表明,方位对平均净光合速率影响显著;胡耀升等[37]对辽东栎(Quercus wutaishanica Mayr)的研究发现,方位对光合生理特性影响显著;赵曦阳等[38]对毛白杨(Populus tomentosa Carr.)无性系光合特性的研究发现,其瞬时光合速率在方位间差异不显著。本研究中,不同方位光合参数及光合色素含量均差异不显著,这可能是因为林分密度较大,树木冠层间相互遮挡,导致方位间光照强度差异较小。
林木光合能力一般随林龄的增长而减小[8, 39],因为林木气孔导度、N含量随林龄的增长而下降,比叶面积随林龄的增长而减小[39],气孔导度和比叶面积的下降会引起叶片扩散阻力的增加,限制大气中CO2向羧化位点移动,而N含量的减小将限制光合器官的形成。本研究中,全冠层光合响应参数在林龄间无显著变化,而臧润国等[40]对不同龄级天山云杉(Picea schrenkiana var. tianshanica (Rupr.) Cheng et Fu.)和刘海涛等[41]对不同林龄乌柳(Salix cheilophila Schneid.)光合特性的研究均发现,光合能力随林龄的增长表现为先增加后降低的变化规律,可能是因为林木生长发育初期需要投入较多的资源来应对各种环境胁迫,而用于提高光合能力的资源较少;当林木进入快速生长期时,逐渐适应环境,投入到光合生长方面的资源逐渐增加,表现为相对较高的光合能力和生长速率;当林木成熟后,较多的资源被用于维持自身的各种代谢过程,导致光合能力下降。
-
冠层对日本落叶松光合色素含量和多数光合参数影响显著,方位和林龄对以上参数的影响不显著。除AQY和CCP外,光合参数均随冠层的升高而增加,光合色素含量则随冠层的升高而下降;方位间光合生理参数的变化不显著,南向和东向光合参数均值分别可以代表 7年生和19年生样木全冠层光合特性。因此,在构建日本落叶松冠层生产力模型和估算冠层生产力时,应详细考虑冠层光合作用的垂直变化。
 DownLoad:
DownLoad: