



-
杨树是目前世界各国最主要的造林及用材树种之一,属于杨柳科(Salicaceae)杨属(Populus),全世界大约100余种,属下通常分5个派,即白杨派(Sect. Leuce Duby)、大叶杨派(Sect. Leucoides)、青杨派(Sect. Tacamahaca)、黑杨派(Sect. Aigeiros)和胡杨派(Sect. Turanga)[1-2]。由于雌雄异株的生物学特性使得杨属的各派在长期的进化过程中不断进行杂交从而产生许多派内及派间的天然杂种,虽然其大大丰富了杨属的遗传多样性,但同时也为如何有效区分和鉴别杨树种类造成了严重的障碍。同样,也正因为杨属这种不断杂交所形成的网状进化特点使得提供较少信息位点的标记方法如形态标记和同功酶等标记方法很难区分辨别杨属种间关系,因此,寻求更加稳定有效的标记方法成为研究杨属树种系统发育关系的重要内容。随着分子生物学技术的发展,已陆续研发出数十种DNA分子标记技术,使得在基因组水平上探究杨属派间、派内种间及无性系间的遗传变异成为可能。与形态标记等相比,DNA分子标记因具有直接比较遗传物质,不受环境因素的干扰,提供信息中性及内容更丰富等特点,使其与其它的标记方法相比更能体现出植物种内和种间的遗传变异,揭示物种演化[3-6]。因此,DNA分子标记被认为是进行遗传变异评价的理想标记,受到越来越多研究学者的青睐。
迄今为止,已经有多种分子标记诸如RAPD[7-8]、ISSR[9]、SSR[10]和AFLP[11-13]等用于杨属派间、派内种间以及种内无性系间的遗传变异的相关研究,其中,AFLP标记由于引物随机,提供的信息量大、重复性好等优点被国内外众多学者作为研究杨属遗传的主要分子标记类型;然而,它却存在着成本高,操作步骤繁琐并且技术要求较高等缺点。SRAP(sequence related amplified polymorphism)是由Li等[14]开发的一种新型分子标记系统,因为没有酶切、预扩及选扩等繁琐步骤,仅利用根据基因中外显子、内含子及启动子碱基含量不同的特点设计的特异性引物进行简单的PCR扩增,所以对技术要求非常简单,并且还可以显著降低成本,节省时间,具有操作简单、高效、引物利用率高、重复性好、费用低等优点[15]。不单如此,它还能提供丰富的遗传信息,据刘艳萍等[16]在利用SRAP对新疆地区的15个杨属树种遗传多样性研究中发现,27对引物竟产生853条多态性片段,多态性比率高达96.8%。SRAP标记广泛应用于遗传变异和亲缘关系分析、遗传多样性分析和指纹图谱构建[5-6]、品种鉴定[17-18]、遗传连锁图的构建和基因克隆和定位[19]以及反映物种进化历程[20-21]的研究中,显示出越来越多的优越性。
本研究以杨属中的白杨派、青杨派、黑杨派和胡杨派22个重要的树种,共33个样本为试验材料,采用SRAP分子标记技术研究杨树的遗传变异和亲缘关系水平,希望借助于研究所揭示的杨树遗传基础,为研究杨属树种的分类鉴定、系统进化以及遗传育种等奠定重要的理论基础。
HTML
-
白杨派、青杨派、黑杨派和胡杨派的22个重要的树种分别在不同地区采样,共33个样本,每个样本采集3个无性系,详细信息见表 1。
树种Species 传统分类派别Sections 采集地Collection place 毛白杨 P.tomentosa♀ ♂ 白杨派Leuce Duby 北京Bejing 银白杨 P.alba♀ ♂ 白杨派Leuce Duby 北京Beijing 中国山杨P.davidiana ♀ ♂ 白杨派Leuce Duby 北京Beijing 美洲山杨P.tremuloides ♀ ♂ 白杨派Leuce Duby 美国U.S.A 河北杨P.hopeiensis ♀ 白杨派Leuce Duby 北京Beijing 新疆杨P.bolleana ♂ 白杨派Leuce Duby 北京Beijing 毛新杨P.tomentosa×P.bolleana ♀ 白杨派Leuce Duby 北京Beijing 银腺杨P.alba×P.glandulosa ♀ ♂ 白杨派Leuce Duby 山东Shandong 美洲黑杨P.deltoides ♀ ♂ 黑杨派Aigeiros 河南Henan 欧美杨P.×euramericana ♀ ♂ 黑杨派Aigeiros 河南Henan 山海关杨 P.canadensis cv. ‘Shanhaiguanensis’ ♀ 黑杨派Aigeiros 河北Hebei 加拿大杨P.canadensis ♂ 黑杨派Aigeiros 内蒙古Inner Mongolia 小叶杨P.simonii ♀ ♂ 青杨派Tacamahaca 内蒙古Inner Mongolia 青杨P.cathayana ♀ ♂ 青杨派Tacamahaca 四川Sichuan 大青杨P.ussuriensis ♀ ♂ 青杨派Tacamahaca 吉林Jilin 三脉青杨P.trinervis ♀ 青杨派Tacamahaca 四川Sichuan 冬瓜杨P.purdomii 青杨派Tacamahaca 青海Qinghai 滇杨P.yunnanensis 青杨派Tacamahaca 云南Yunnan 藏川杨P.szechuanica var. tibetica 青杨派Tacamahaca 四川Sichuan 毛果杨P.trichocarpa 青杨派Tacamahaca 美国U. S. A 辽杨P.maxuimowiczii 青杨派Tacamahaca 北京Beijing 胡杨P.euphratica ♀ ♂ 胡杨派Turanga 内蒙古Inner Mongolia Table 1. The information of materials
-
总DNA的提取按照DNeasy Plant Mini Kits(Qiagen, Inc., Valencia, CA, USA)说明书描述的方法进行。待1%琼脂糖凝胶电泳检测DNA质量后,稀释至所需浓度(20 ng·L-1),-20℃保存备用。
-
SRAP反应体系采用经均匀设计试验两轮优化后的体系[22],即在25 μL反应体系中包含MgCl2 3.5 mmol·L-1,dNTPs 0.20 nmol·L-1,引物0.44 μmol·L-1,TaqDNA聚合酶1.50 U,模板28 ng·L-1,其中,MgCl2、dNTPs和Taq DNA聚合酶均购自天根公司。正反向引物的序列信息见表 2,引物由上海生工公司合成,正反向引物随机配对后形成72对引物组合。
编号Code 正向引物Forward primer 编号code 反向引物Reverse primer Mel 5’-TGAGTCCAAACCGGATA-3’ Eml 5’-GACTGCGTACGAATTAAT-3’ Me2 5’-TGAGTCCAAACCGGAGC-3’ Em2 5’-GACTGCGTACGAATTTGC-3’ Me3 5’-TGAGTCCAAACCGGAAT-3’ Em3 5’-GACTGCGTACGAATTGAC-3’ Me4 5’-TGAGTCCAAACCGGACC-3’ Em4 5’-GACTGCGTACGAATTTGA-3’ Me5 5’-TGAGTCCAAACCGGAAG-3’ Em5 5’-GACTGCGTACGAATTAAC-3’ Me6 5’-TGAGTCCAAACCGGTAA-3’ Em6 5’-GACTGCGTACGAATTGCA-3’ Me7 5’-TGAGTCCAAACCGGTCC-3’ Em7 5’-GACTGCGTACGAATTGAG-3’ Me8 5’-TGAGTCCAAACCGGTGC-3’ Em8 5’-GACTGCGTACGAATTGCC-3’ Em9 5’-GACTGCGTACGAATTTCA-3’ Table 2. The primers sequences used in SRAP amplification system for Populus
PCR扩增在PTC-200 PCR仪上进行。扩增程序:94℃预变性5 min;前5个循环为94℃变性50 s,35℃复性50 s,72℃延伸90 s;后30个循环将复性温度升为50℃;最后72℃延伸10 min,并于10℃保存。
-
扩增得到的PCR产物用1.8%琼脂糖凝胶电泳检测。统计各个样本不同引物组合在相同片段位置上产生的谱带,以“1”和“0”统计各位点条带的有无,有条带记为1,无条带记为0。
-
多态性指扩增得到的多态性条带数占总扩增条带数的比例。遗传相似系数(GS)和遗传距离(GD)采用Nei等[23]的计算公式:
式中:Nxy表示样本x和y的公共带数,Nx和Ny分别是样本x和y的带数。
利用NTSYS-pc2.1软件和非加权配对算术平均法(UPGMA)进行聚类。
1.1. 试验材料
1.2. 方法
1.2.1. DNA提取
1.2.2. SPRA-PCR反应
1.2.3. PCR产物扩增结果检测与记录
1.2.4. 数据统计与分析
-
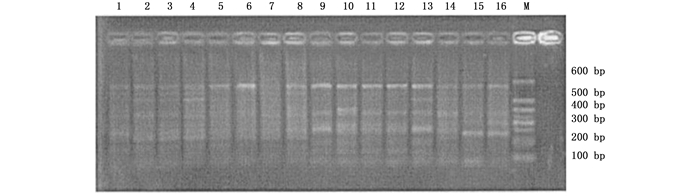
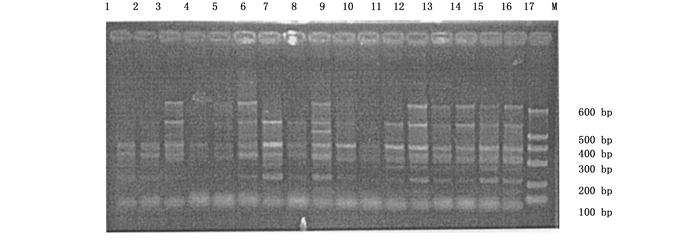
本试验首先采用随机配对的72对SRAP引物组合对采集自不同地区的22个杨属树种共33个样本进行扩增,最后筛选出40对可获得清晰、重复性好、具有多态性条带的引物组合。40对引物共产生487条比较清晰的条带,片断长度集中在100~600 bp之间,其中,表现出多态性的条带有441条,多态性条带比率为90.55%。平均每对引物扩增出12.1条带,多态性带11.0条。代表性图谱见图 1和图 2。
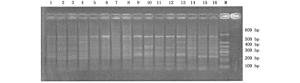
Figure 1. Amplification results of the primer combination "Me8 Em2" in parts of the accessions
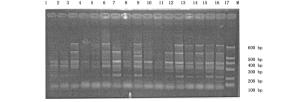
Figure 2. Amplification results of the primer combination "Me6 Em1" in parts of the accessions
-
对采集自不同地区的33个杨属样本的派内种间遗传变异和亲缘关系分析表明:在扩增得到的487条带中,12个青杨派树种扩增出189条多态性条带,各样本间遗传相似系数为0.515 6~0.893 3,遗传距离为0.106 7~0.484 4,其中,遗传距离最大的是辽杨和三脉青杨雌株,最小的是大青杨雄株和大青杨雌株(表 3、4);13个白杨派树种扩增出183条多态性条带,各样本之间遗传相似系数为0.529 4~0.886 6,遗传距离为0.113 4~0.470 6,其中,遗传距离最大的是毛白杨雄株和银白杨雌株,最小的是中国山杨雌株和中国山杨雄株;6个黑杨派树种扩增出91条多态性条带,各样本之间遗传相似系数为0.678 8~0.886 2,遗传距离为0.113 8~0.321 2,其中,遗传距离最大的是加拿大杨雄株和欧美杨雄株,最小的是欧美杨雌株和欧美杨雄株;2个胡杨派树种扩增出24条多态性条带,遗传相似系数为0.910 3,遗传距离为0.089 7(表 4)。派内种间遗传相似性最大的是胡杨雌株和胡杨雄株,可能表明二者之间具有较小的遗传差异。
大青杨雌 大青杨雄 青杨雌 青杨雄 毛果杨 小叶杨雄 冬瓜杨 小叶杨雌 三脉青杨雌 藏川杨 辽杨 滇杨 大青杨雌 1 大青杨雄 0.893 3 1 青杨雌 0.812 5 0.869 0 1 青杨雄 0.805 6 0.763 2 0.839 5 1 毛果杨 0.641 8 0.662 0 0.671 1 0.676 5 1 小叶杨雄 0.700 0 0.761 9 0.775 3 0.753 1 0.763 2 1 冬瓜杨 0.726 1 0.775 8 0.777 1 0.729 6 0.644 3 0.800 0 1 小叶杨雌 0.709 7 0.736 2 0.786 1 0.751 6 0.680 3 0.855 5 0.823 5 1 三脉青杨雌 0.625 0 0.644 7 0.642 0 0.643 8 0.676 5 0.777 8 0.679 2 0.751 6 1 藏川杨 0.705 9 0.745 3 0.771 9 0.748 4 0.648 3 0.760 2 0.773 8 0.759 0 0.683 9 1 辽杨 0.698 4 0.656 7 0.638 9 0.640 6 0.576 3 0.611 1 0.638 3 0.633 1 0.515 6 0.656 9 1 滇杨 0.707 5 0.735 5 0.715 2 0.698 0 0.661 9 0.727 3 0.777 8 0.737 5 0.657 7 0.810 1 0.748 1 1 Table 3. Genetic similarity coefficient in Sect. Tacamahaca
派别
Sections遗传距离的变幅
Range of genetic distance均值
Mean of genetic distance青杨派Tacamahaca 0.106 7~0.484 4 0.280 9 白杨派Leuce Duby 0.113 4~0.470 6 0.301 8 黑杨派Aigeiros 0.113 8~0.321 2 0.224 2 胡杨派Turanga 0.089 7 0.089 7 Table 4. The range and mean of genetic distance in sections of Populus
-
对杨属4个派的33个样本的派间遗传变异和亲缘关系分析表明:各杨树样本间的遗传相似系数为0.448 6~0.910 3,其中,不同派的样本间遗传相似性最小的是辽杨和毛白杨雄株,其相似系数为0.448 6,遗传距离为0.551 4;不同派的样本间遗传相似性最大的是青杨雌株和中国山杨雄株,为0.780 7,遗传距离为0.219 3。杨属派间的平均遗传距离最大的是白杨派和胡杨派,为0.377 7;其次是白杨派和黑杨派(0.367 1)、青杨派和胡杨派(0.364 5)及青杨派和白杨派(0.363 8),这三组的遗传距离中等;之后是青杨派和黑杨派(0.336 0),而遗传距离最小的是黑杨派和胡杨派,为0.333 5(表 5)。
派别Section 青杨派Tacamahaca 白杨派Leuce Duby 黑杨派Aigeiros 胡杨派Turanga 青杨派Tacamahaca 0.363 8 0.336 0 0.364 5 白杨派Leuce Duby 0.219 3~0.551 4 0.367 1 0.377 7 黑杨派Aigeiros 0.232 6~0.485 3 0.236 6~0.472 9 0.333 5 胡杨派Turanga 0.302 6~0.461 5 0.294 8~0.496 2 0.289 5~0.372 5 Table 5. The range and mean of genetic distance among sections in Populus
-
根据各杨树种间的遗传相似系数,采用NTSYS-pc2.1版分析软件,利用非加权配对算术平均法(UPGMA)进行聚类分析,构建了33个杨属样本的亲缘关系树状图。由聚类图(图 3)可知:33个杨树材料在遗传相似系数0.608的位置可以分成2大类群,即黑杨派、胡杨派和青杨派为一个分支,白杨派单独形成另一个分支,可能暗示黑杨派、胡杨派和青杨派间的亲缘关系更近。以0.628的相似系数为阈值,可将材料分为4大类:第Ⅰ类包括9个树种的12个样本,均为青杨派树种;第Ⅱ类有4个树种的6个样本,均为黑杨派树种;第Ⅲ类有1个树种的2个样本,为胡杨派树种;第Ⅳ类有8个树种的13个样本,为白杨派树种。

Figure 3. Dendrogram of 33 Populus genotypes from cluster analysis based on SRAP markers
2.1. SRAP标记的多态性
2.2. 杨属派内种间的遗传变异和亲缘关系分析
2.3. 杨属派间的遗传变异和亲缘关系分析
2.4. 聚类分析

-
青杨派树种种类众多,且分布的地理跨度较广,是研究杨属派内种间遗传变异和亲缘关系的理想对象。由本研究构建的UPGMA系统发育树(图 3)可以看出:除了毛果杨和三脉青杨形成一个单独分支之外,青杨派的其它种包括小叶杨、滇杨等聚类形成另一个分支。青杨派中藏川杨的归属是比较有争议的,藏川杨属于川杨(P.szechuanica)的一个变种。Hamzeh等[24]研究发现,川杨与青杨派有相似的核糖体ITS序列,但同时也有与黑杨派相似的叶绿体TrnT-F序列,说明川杨是杂种起源。对西南地区杨属青杨派乡土树种的研究表明,藏川杨与昌都杨(P.qamdoensis)、德钦杨(P.haoana)等亲缘关系很近[25-26];而本研究中藏川杨与滇杨、辽杨聚类在一起,这与李善文等[12]和史全良等[27]研究结果相一致,推测藏川杨属于青杨派。滇杨作为少有的低纬度高海拔杨属树种,其雌雄花蕊的长度与蒴果的瓣裂数与青杨派某些其他树种存在差异,滇杨的分类地位也存在着较大的争议。史全良等[27]利用IST序列构建的系统发育树认为,滇杨与白杨派的银白杨和欧洲山杨亲缘关系较近;陈珂[28]研究发现,在SSR和ISSR的聚类分析中,滇杨明显与青杨派其它树种分离而独立形成一组,而在AFLP和cpDNA的聚类分析中,滇杨与小叶杨、三脉青杨聚类形成一支;Cervera等[11]和李善文等[12]研究结果表明,滇杨和小叶杨的亲缘关系较近。本研究中,滇杨与藏川杨和辽杨聚为一类,且与小叶杨等其他青杨派物种形成一个大的分支,可能暗示滇杨与青杨派其他树种的亲缘关系较近,推测滇杨同样属于青杨派树种。
-
传统上白杨派可分为山杨亚派和白杨亚派,山杨亚派主要包括河北杨、欧洲山杨(P.tremula)、美洲山杨、中国山杨和响叶杨(P.adenopoda)等,银白杨、新疆杨、毛白杨等是白杨亚派的代表树种。但由本研究构建的UPGMA系统发育树(图 3)可以看出:山杨亚派的中国山杨、河北杨、美洲山杨和白杨亚派的银白杨、新疆杨在遗传相似系数在0.638水平时聚类形成一个分支,而其它的种包括由银白杨与腺毛杨(P.glandulosa)杂交形成的银腺杨、由毛白杨与新疆杨杂交产生的毛新杨以及白杨亚派的毛白杨则形成另外一个分支。由此结果可以看出,首先河北杨与中国山杨最终聚类成一个分支,推测这二者亲缘关系比较近,符合传统分类上河北杨属于山杨亚派的归类。其次,由聚类结果看,山杨亚派并不能清晰地与白杨亚派区分开,可能的原因之一是中国山杨、欧洲山杨和美洲山杨在归类上存在分歧。史全良等[27]研究显示,欧洲山杨与银白杨和银灰杨形成一个单独分支,而且白杨派内部各树种间的关系形成一个多歧平行的结构。Wang等[29]研究发现,欧洲山杨更接近于中国山杨,美洲山杨与大齿杨(P.grandidentata)形成另一个分支,因此,可能传统上对美洲山杨和中国山杨的分类存在疑问。另一个可能的原因是白杨派的很多树种都是杂种起源,王东升[30]综合核基因和cpDNA系统发育树以及Structure结果显示,河北杨的起源方式是由父本银白杨向母本中国山杨单向授粉形成的杂种,而毛白杨形成比较复杂,在cpDNA树中毛白杨分成了2种类型,毛白杨Ⅰ可能是由父本银白杨向母本响叶杨单向授粉形成的杂种,而毛白杨Ⅱ推测可能是(中国山杨♀×响叶杨♂)×银白杨的复合杂种,而且,杂种起源也导致种内不同类型及单株之间遗传变异很大。因此,由于白杨派树种复杂的起源方式,可能传统的白杨和山杨2个亚派的分类方式并不准确。
-
黑杨派树种在世界范围内广泛分布,具有速生、易繁殖等特点,是重要的用材树种。由本研究构建的UPGMA系统发育树(图 3)可以看出:6个黑杨派树种能聚类成一个单独分支,之后加拿大杨和山海关杨相继分化,而美洲黑杨和欧美杨在遗传相似系数为0.766水平上才出现分化,这可能说明美洲黑杨和欧美杨之间亲缘关系较近,其遗传相似系数为0.780 5~0.832 3。本研究中,加拿大杨是美洲黑杨和欧洲黑杨的杂交种,山海关杨是欧美杨的栽培变种,加拿大杨和山海关杨的遗传相似系数为0.752 9,最终各自聚类为一个分支;相对于加拿大杨,山海关杨与美洲黑杨和欧美杨在遗传相似系数为0.734时才出现分化,可能暗示山海关杨与美洲黑杨和欧美杨亲缘关系相对更近。不同的杂交种与父母本的遗传距离存在差异,可能是由于不同的杂交组合所产生的杂种优势差异导致的。
-
通过系统发育树可以看出,在遗传相似系数0.608水平上,白杨派首先单独形成一个分支,而黑杨派、胡杨派和青杨派形成另一分支;在遗传相似系数0.618水平上,青杨派才与黑杨派、胡杨派分开形成另一分支。这一结果可能暗示白杨派与其他各派的亲缘关系较远,青杨派与黑杨派和胡杨派的亲缘关系则比较近,黑杨派和胡杨派关系则最为紧密。
白杨派、黑杨派和青杨派之间的聚类结果与先前采用的杂交试验和同工酶试验的结果相似[31-32]。张绮纹等[33]对杨属各派花粉的扫描电镜结果显示,青杨派和黑杨派树种花粉粒表面均有云片状雕纹,雕纹上有小的乳凸和不规则小孔,虽有不同但基本特征是一致的;白杨派的山杨亚派和白杨亚派树种的花粉表面纹饰相似,但有区别,且与其他各派均不同。杨属树种派间杂交试验显示,黑杨派和青杨派树种杂交表现出较好的亲和性,易产生杂种,白杨派与其它派之间存在明显的杂交障碍。对杨属不同派间树种做同工酶试验结果也显示白杨派与青杨派和黑杨派的同工酶谱带相似性低,亲缘关系较远。因此,从杂交试验、同工酶分析、DNA多态性3个方面分析的结果可以推测黑杨派与青杨派之间的亲缘关系较近,与白杨派亲缘关系较远。远缘杂交试验表明[31, 34-35],青杨派与白杨派的派间杂交亲和力强于黑杨派与白杨派,相对容易产生杂种。本研究中,青杨派和白杨派的遗传距离为0.363 8,小于白杨派和黑杨派的遗传距离(0.367 1),推测青杨派和白杨派的亲缘关系相对更近,与前人研究结论相一致。
胡杨派的聚类方向却比较有争议。胡杨派共3个种,即胡杨、灰胡杨(P.pruinosa)和肯尼亚胡杨(P.ilicifolia)。胡杨派树种叶片呈浅裂形或者深裂形,对干旱具有较强的抗性,具备很多杨柳科的古老特征,很多学者认为其是杨属物种最原始的类群[36]。Wang等[36]利用单拷贝核基因和cpDNA研究发现,在cpDNA系统发育树上胡杨派和青杨派亲缘关系较近,核基因树表明胡杨派是相对古老的类群。李善文等[12]发现,胡杨派可以形成一个单独的分支,独立于黑杨派、青杨派和白杨派之外。本研究中发现,胡杨派同黑杨派最终聚类形成一支,与史全良等[27]的研究结果相一致,可能暗示胡杨派和黑杨派关系比较紧密。但研究显示[33],胡杨派树种花粉粒表面呈分立的疣状凸起,与杨属其他各派花粉相比比较特殊;育种实践也证明,若不经过特殊处理,胡杨派同其它派之间几乎不可配[1]。对于出现胡杨派聚类方向与育种实践不一致的情况,分析可能的原因是实验中胡杨派树种只选取了胡杨,取样品种不足,容易因个体间的差异等因素影响其聚类方向。
3.1. 杨属派内种间的遗传变异和亲缘关系
3.1.1. 青杨派
3.1.2. 白杨派
3.1.3. 黑杨派
3.2. 杨属派间的遗传关系
-
采用SRAP分子标记技术,对采集自不同地区的22个杨属树种33个样本进行遗传变异和亲缘关系分析,共扩增出487条清晰条带,多态性条带比率为90.55%。研究结果显示,不同的杨树品种在SRAP位点有较高的多态性。聚类分析结果表明,33个样本可分为4大类,这一聚类结果与传统的分类结果一致,且白杨派单独形成一个分支,黑杨派、胡杨派和青杨派形成另一分支,可能暗示白杨派与其他各派的亲缘关系较远,青杨派与黑杨派和胡杨派的亲缘关系则比较近。本研究表明,SRAP分子标记适用于分析杨属的遗传变异和亲缘关系,研究结果为杨属树种的分类鉴定、系统进化以及遗传育种等提供了一定的理论依据。
 DownLoad:
DownLoad: