



-
楸树(Catalpa bungei C.)属紫葳科(Bignoniaceae)梓属(Catalpa)落叶大乔木,是我国独有的珍贵用材树种,古有“木王”美称。楸树材性优良,干形端直,节间较少,出材率高;木材纹理通直,木质结构紧密;木材物理性能好,抗弯、抗压性能高;化学性质稳定,耐腐蚀耐水湿。不仅如此,楸树适应性强,在我国分布广泛,但由于人们长期滥砍滥伐,导致楸树数量急剧下降。楸树用材缺口巨大,价格颇高,因此亟待选育出速生优良品种以满足楸树珍贵木材供应[1-2]。
研究表明,林木生长动态存在一定的规律,依据林木生长状况采用有效的数学模型对其生长过程进行探索,可以更深入地了解林木种间和种内竞争关系以及其对有机物的累积和消耗规律。以此为基础,可对现行经营模式、经营密度下林分单株树木的生长潜力和生长状况进行科学评价与判断,并制定培育措施和管理方案[3]。如曹健康等[4]通过对光皮桦(Betulaluminifera H. Winkl.)人工幼林的物候期和生长节律的研究,采取了相应的抚育管理及施肥措施以提高光皮桦全年生长量。此外,林木生长过程还可为确定以经济效益为主的速生用材林的合理收获年限,以降低因过早或过迟砍伐造成的经济损失。如木麻黄(Casuarina equisetifolia Forst.)无性系轮伐期为造林后6~9年,相比较于福建实生苗人工林25年数量成熟龄早许多,为海南地区木麻黄无性系最佳轮伐期提供了参考依据[5]。
另一方面,对林木生长过程的研究可剖析其生长特性,对林木遗传改良有指导作用[6-8]。如青钱柳(Cyclocarya paliurus (Batal.) Iljinsk.)种源[9]、南方红豆杉(Taxus wallichiana var. mairei (Lemee et Levl.) Cheng et L. K. Fu)家系[10]和杨树(Populus L.)无性系[6]等林木不同水平上均存在显著的生长节律变异,主要体现在如生长速率、快速生长持续的时期长短等方面,而这种差异正好也为人工选育提供了新的思路。以往的遗传改良研究大多只是评价不同基因型目标性状的优劣[11],忽视了其性状优良的内在原因。鉴于此,本研究以32个楸树无性系为试验材料,从楸树无性系单株材积的早期生长过程出发,探究其单株材积变化的规律,提高楸树优良无性系选择的准确性,同时为楸树早期管理提供更加科学的理论指导。
HTML
-
试验地位于河南省南阳市潦河镇潦东村(32°.93′ N, 112°.41′ E),属季风大陆湿润半湿润气候,四季分明。海拔145 m,土壤类型为黄棕壤土,自然肥力较高。年平均气温14.4~15.7℃,7月平均气温26.9~28.0℃,1月平均气温0.5~2.4℃。年降水量703.6~1 173.4 mm,年日照时数1 897.9~2 120.9 h,年无霜期220~245 d。于2009年春季营建楸树杂种无性系试验林,试验采用完全随机区组设计,每小区2株,5个区组。株行距2.5 m×4 m。
-
2009—2015年每年末在树木生长停滞期时对全林进行每木检尺,测定其树高(H)和胸径(DBH),并据此估算单株材积。连年生长量为当年性状测定值减去上一年性状值,平均生长量为当年性状大小除以树龄。
单株材积估算公式[12]:
-
采用Logistic模型对楸树无性系进行生长曲线拟合,通过SPSS 22.0的非线性回归方法求出模型各参数值和决定系数R2,然后运用模型计算楸树无性系的单株材积生长特征值。生长曲线的Logistic模型拟合详细方法见参考文献[13-14]。
-
使用SPSS 22.0以小区均值为单位进行性状的方差分析,以检验楸树无性系各性状是否具有显著差异,并分析树高和胸径遗传参数的树龄变化规律。
聚类分析、Pearson相关分析均通过R语言软件的“ape”和“psych”程序包完成,聚类分析采用层次聚类法,以欧氏距离为划分标准。
方差分析线性模型为:
式中:yijk为第i无性系第j区组第k小区观察值的平均值;μ为群体平均效应;Ci为第i无性系效应;Bj为第j区组效应;eijk为机误。
重复力公式[15]:
式中:R为重复力,F为方差分析F检验值。
变异系数(cv)公式[16]:
式中:σ为标准差,X为性状均值。
1.1. 试验材料与试验地概况
1.2. 生长性状测定
1.3. 生长曲线拟合
1.4. 数据处理
-
楸树无性系生长性状方差分析结果(表 1)表明:无性系间1 a树高、胸径和单株材积差异不显著(P>0.05),其余年份各生长性状皆存在极显著差异,这表明造林2 a后楸树无性系生长性状表现出显著差异,遗传变异丰富,具有较好的遗传改良基础。不同区组中,楸树无性系生长性状在第1年差异不显著,7 a树高的区组效应亦不显著,其余年份不同生长性状存在极显著的区组效应(P < 0.01),说明包括试验地土壤肥力、土壤含水量等方面的环境效应对无性系生长具有显著影响,因此,不能忽视对楸树生长环境优化的人工管理。
性状
Traits树龄/a
Tree ages无性系Clone 区组Block 均方MS F值F value P值P value 均方MS F值F value P值P value 树高Height 1 0.15 1.39 0.11 0.04 0.37 0.83 2 0.42 2.37 0.00 0.58 3.28 0.01 3 0.55 4.46 0.00 0.82 6.70 0.00 4 0.71 3.37 0.00 1.35 6.40 0.00 5 0.74 3.75 0.00 0.96 4.85 0.00 6 0.56 2.93 0.00 1.59 8.33 0.00 7 0.49 2.15 0.00 0.15 0.68 0.61 胸径DBH 1 0.20 1.29 0.17 0.11 0.68 0.60 2 0.56 3.90 0.00 1.22 8.47 0.00 3 1.42 5.29 0.00 5.59 20.77 0.00 4 3.33 4.64 0.00 20.63 28.79 0.00 5 4.22 3.96 0.00 18.79 17.63 0.00 6 5.85 5.27 0.00 15.42 13.87 0.00 7 7.63 5.41 0.00 6.15 4.37 0.00 单株材积Individual volume 1 0.000 0 1.20 0.24 0.000 0 0.69 0.60 2 0.000 0 3.38 0.00 0.000 1 6.39 0.00 3 0.000 1 5.22 0.00 0.000 5 18.65 0.00 4 0.000 7 4.17 0.00 0.003 6 21.96 0.00 5 0.001 7 2.80 0.00 0.006 6 10.59 0.00 6 0.002 7 4.03 0.00 0.008 5 12.59 0.00 7 0.003 7 3.99 0.00 0.002 6 2.79 0.03 Table 1. The variance analysis of growth characters of C. bungei
-
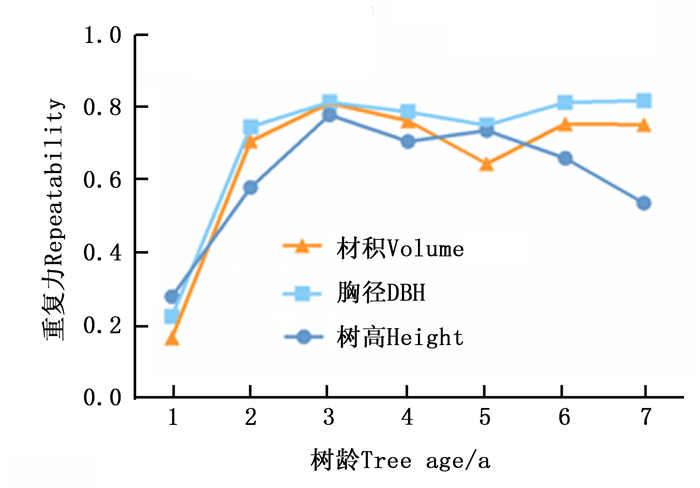
从图 1可看出:树高、胸径和单株材积的无性系重复力随林龄的变化趋势大致相同,第1年的重复力均较小,随着年龄的增加,重复力逐渐增大,在第3年达到一个高峰,随后又略有回落。自第5年起,胸径和单株材积的无性系重复力呈上升趋势而树高的无性系重复力呈下降趋势。1 a三种性状的无性系重复力均较低,这与苗木栽植对土壤等环境还未完全适应有较大关系,此阶段苗木生长受环境主导,而这种状况也大大降低了重复力的估算精度。自第2年始至第7年,性状重复力较高,变幅减小,遗传效应对生长调控占据了主要作用。在3个生长性状中,胸径的重复力最高(除第1年外),稳定性最强。6 a和7 a单株材积的重复力保持较高值且相对稳定,这意味着楸树单株材积受遗传控制较强。

Figure 1. Repeatabilities of tree height and DBH at different age stages in C.bungei
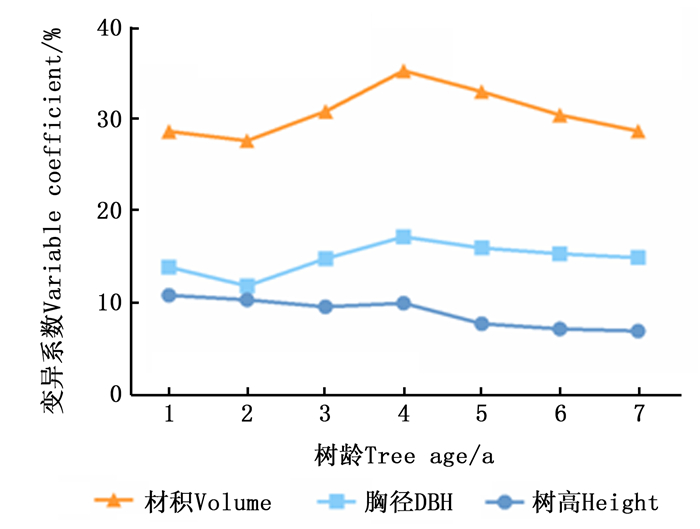
变异系数可作为衡量观测值变异程度的指标,反映无性系该性状可利用的改良潜力。由图 2可以看出:楸树无性系单株材积、胸径和树高变异系数在不同年份存在一定差异,其中,单株材积的变异系数最大,胸径、树高次之。单株材积与胸径的变异系数有着相似的变化趋势,在1~2 a,变异系数呈较为稳定趋势,变化不大;在2~7 a,有先升高后降低的过程,均在4 a达到变异最大值。树高的变异系数较小且变化范围也较小。说明楸树无性系的单株材积具有丰富的遗传变异,遗传改良潜力大。相对的,树高改良价值较小。

Figure 2. Variation parameters of different traits at different age stages in C.bungei clones
-
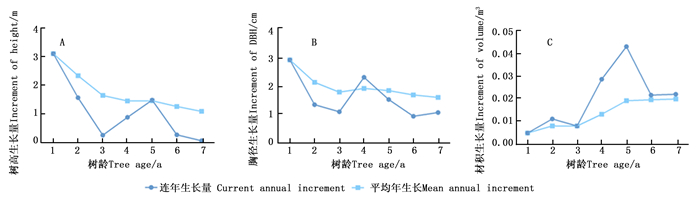
对楸树无性系早期生长过程进行分析,树高和胸径有着相似的生长规律,1 a平均年生长量和连年生长量达到最大,生长速度最快(图 3A、B)。树高和胸径平均年生长量自1 a始逐年降低。1~3 a连年生长量逐年下降,3~6 a进入先增加后降低的过程,分别在5、4 a达到高峰,6~7 a树高连年生长量继续下降,而胸径有所回升。由图 3可知:单株材积平均年生长量在不断增加,1~3 a增加缓慢,3~5 a增加迅速,自第5 a年后始终保持较高增长水平(图 3C)。单株材积连年生长量1~5 a持续增加,在5 a达到巅峰,随后下降,在6~7 a保持稳定增长速度,说明楸树单株材积生长主要集中在后期。单株材积的平均年生长量与连年生长量并未相交,说明楸树单株材积生长未达到数量成熟龄。

Figure 3. Different growth traits curve of C.bungei clones
-
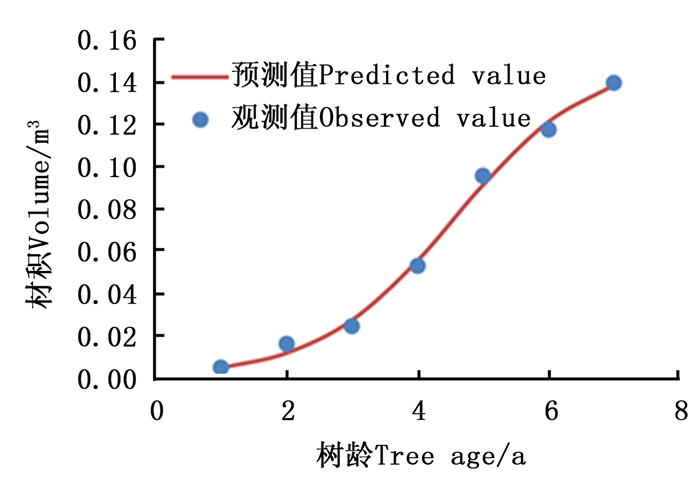
为剖析楸树的早期生长过程,对楸树32个无性系单株材积生长量指标早期生长过程采取Logistic拟合。拟合结果(表 2)表明:Logistic曲线线性回归相关系数均达到99%,达到极显著相关,楸树无性系拟合曲线拟合度很高,说明Logistic可以较好地说明楸树无性系单株材积的早期生长过程。由图 4可知:楸树7年生无性系单株材积生长呈现出“S”型曲线趋势,实际值与预测值比较接近,不同林龄时的单株材积残差值均较小,拟合的生长预估模型较科学, 预测值能够较好地反应林分生长情况。
无性系
Clones模型拟合参数
Model parametersR2 单株材积生长特征值
Characteristic value of individual volumeK/m3 a b tm/a t1/a t2/a LGT/a MGR/(m3·a-1) LGR/(m3 ·a-1) LGI/m3 1-1 0.21 66.28 0.79 0.994 5.31 3.64 6.98 3.34 0.041 9 0.034 9 0.122 6 12-09 0.14 155.09 1.06 0.997 4.76 3.52 6.00 2.49 0.036 9 0.030 8 0.080 5 12-13 0.11 121.29 1.07 0.993 4.47 3.24 5.69 2.45 0.028 3 0.023 6 0.060 8 13-05 0.21 60.86 0.76 0.991 5.40 3.67 7.13 3.46 0.039 9 0.033 3 0.121 1 13-06 0.18 93.64 0.95 0.995 4.79 3.40 6.18 2.78 0.042 0 0.035 0 0.102 4 16-01 0.16 54.95 0.86 0.995 4.68 3.14 6.22 3.08 0.035 2 0.029 3 0.094 8 16-04 0.16 80.75 0.99 0.996 4.46 3.12 5.79 2.67 0.038 6 0.032 1 0.090 4 16-05 0.15 104.92 1.13 0.994 4.10 2.94 5.26 2.32 0.041 7 0.034 7 0.084 8 16-07 0.16 53.89 0.89 0.996 4.48 3.00 5.96 2.96 0.036 2 0.030 1 0.093 8 16-10 0.19 51.15 0.75 0.989 5.22 3.47 6.96 3.49 0.036 2 0.030 2 0.110 9 17-05 0.09 117.03 1.03 0.988 4.62 3.34 5.89 2.55 0.024 4 0.020 3 0.054 7 17-06 0.11 73.06 0.99 0.994 4.32 2.99 5.64 2.65 0.028 0 0.023 3 0.065 1 18-09 0.13 72.32 0.91 0.997 4.71 3.26 6.16 2.90 0.030 2 0.025 2 0.076 7 19-01 0.22 69.35 0.87 0.991 4.86 3.35 6.36 3.02 0.047 4 0.039 5 0.125 3 19-12 0.13 98.03 1.05 0.993 4.36 3.11 5.62 2.51 0.034 4 0.028 7 0.075 6 19-27 0.19 105.26 1.01 0.996 4.60 3.30 5.90 2.60 0.048 1 0.040 1 0.109 7 20-01 0.16 179.84 1.25 0.987 4.16 3.10 5.21 2.11 0.049 0 0.040 8 0.090 6 20-02 0.21 62.95 0.73 0.991 5.66 3.86 7.46 3.60 0.039 1 0.032 6 0.123 3 20-06 0.15 105.08 1.14 0.994 4.09 2.93 5.25 2.31 0.041 8 0.034 8 0.084 8 21-02 0.11 119.26 1.12 0.990 4.27 3.10 5.45 2.35 0.031 8 0.026 5 0.065 6 21-03 0.13 66.59 0.86 0.997 4.91 3.37 6.45 3.08 0.027 5 0.022 9 0.074 2 22-01 0.18 87.36 0.95 0.991 4.68 3.30 6.06 2.76 0.042 7 0.035 6 0.103 3 22-03 0.20 88.02 1.00 0.998 4.47 3.15 5.78 2.63 0.050 0 0.041 6 0.115 0 22-05 0.17 76.57 0.90 0.994 4.81 3.35 6.26 2.92 0.037 9 0.031 6 0.097 0 22-07 0.19 75.16 0.90 0.997 4.81 3.34 6.28 2.93 0.042 9 0.035 7 0.110 3 22-08 0.16 149.59 1.18 0.993 4.23 3.12 5.35 2.23 0.045 9 0.038 2 0.089 6 22-10 0.14 43.46 0.79 0.988 4.75 3.09 6.40 3.31 0.027 3 0.022 8 0.079 4 23-05 0.10 111.92 1.14 0.983 4.14 2.98 5.29 2.31 0.029 3 0.024 4 0.059 4 6-05 0.17 152.16 1.13 0.992 4.44 3.28 5.61 2.33 0.048 2 0.040 1 0.098 3 7-01 0.08 86.02 0.99 0.988 4.52 3.18 5.85 2.67 0.019 1 0.015 9 0.044 7 9-05 0.14 138.93 1.12 0.994 4.39 3.22 5.57 2.35 0.039 6 0.033 0 0.081 4 9-1 0.16 47.54 0.79 0.992 4.88 3.22 6.55 3.33 0.031 5 0.026 3 0.092 1 均值Mean 0.16 92.76 0.97 0.993 4.64 3.25 6.02 2.76 0.037 3 0.031 1 0.089 9 注:t1:单株材积快速增长期起始时间;t2:单株材积快速增长期终止时间;tm:单株材积最大相对生长速率出现时间;MGR:最大生长速率;LGR:线性生长速率;LGI:线性生长量;LGT:线性生长持续时间。
Note:t1:The initial time of rapid growth of individual volume;t2:The end of rapid growth of individual volume;tm:The occurrence time of Maximum relative growth rate of individual volume;MGR: The maximum growth rate;LGR: Linear growth rate;LGI: Linear growth increment;LGT: Linear growth time。Table 2. Parameters of volume Logistic fitting curve and Characteristic value of individual volume of C. bungei clones

Figure 4. Individual volume Logistic fitting curve of C. bungei clones
生长参数K值是反应模型拟合条件下无性系单株材积生长理论能达到的最大值,楸树无性系间K值差异较大,无性系19-01的K值最大(0.22),而7-01的K值最小(0.08),前者为后者的2.75倍(表 2)。楸树无性系约从3 a开始进入单株材积快速生长期,持续2~4 a左右,约在4.64 a时连年生长量达到最大值,并在5~7 a结束快速生长,只有无性系20-02和13-05的快速生长持续时间超过了7 a。无性系20-06的t1和tm值最小,最快开始单株材积线性生长,也最早达到连年生长量最大值,但是线性阶段生长量并不高(0.084 8 m3)。20-02的t1、t2、tm、LGT值最大,整个线性生长期较其他无性系均晚,无性系20-06和20-02 LGR和MGR值差异不大,但20-20的线性阶段生长量较20-06高(45.40%),说明线性生长过早并不有利于线性生长期单株材积的累积。无性系22-03的LGR和MGR值在所有试验的无性系中最大(0.05 m3·a-1、0.041 6 m3·a-1),反之无性系7-01的最小(0.019 1 m3·a-1、0.015 9 m3·a-1),二者的线性生长时期时间点参数差异不大,但前者线性阶段生长量为后者的2.6倍,说明线性阶段生长速率对单株材积的生长量影响巨大。19-01线性阶段持续时间较长(3.02 a),线性阶段生长速率也比较高(LGR= 0.039 5 m3·a-1、MGR=0.047 4 m3·a-1),故而线性阶段生长量最大(0.125 3 m3),说明LGR、MGR、LGI较高均有利于楸树无性系单株材积的增长。7年生楸树无性系单株材积与生长节律参数相关性分析结果(图 5)发现:7 a单株材积生长量与最大生长速率(MGR)、线性生长速率(LGR)、线性生长量(LGI)呈极显著正相关(P<0.01),7 a单株材积生长量与tm、t1、t2呈现低度正相关。与上文无性系的结果一致,这表明单株材积生长量受线性生长时间参数影响较小,而无性系的生长速率才是决定其单株材积增长的决定性因素,速生期的生长速率越大其最终单株材积量也越大。

Figure 5. Pearson correlation analysis of individual volume and growth parameters of 7-year-old C. bungei clones
-
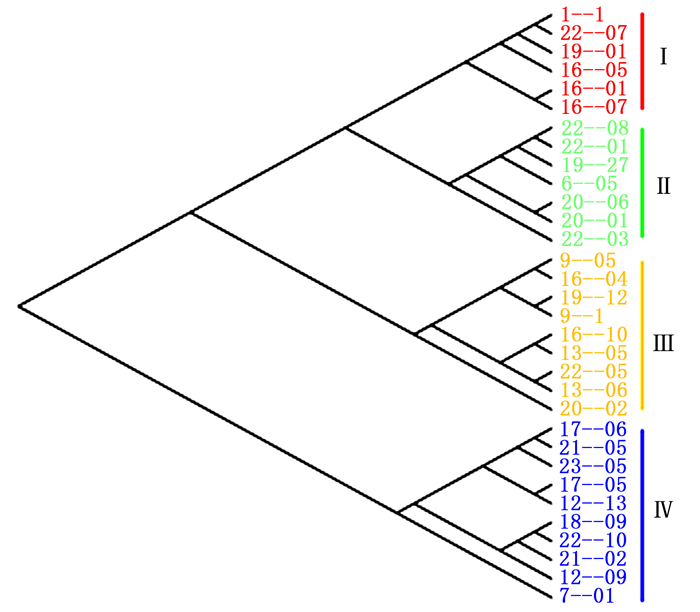
采用层次聚类法对32个楸树无性系进行类群的划分,由图 6可以看出:32个楸树无性系可分为4大类,其中,第Ⅰ类型包括1-1、22-07、19-01、16-05、16-01和16-07共6个无性系;第Ⅱ个类型包括22-08、22-01、19-27、6-05、20-06、20-01和22-03共7个无性系;第Ⅲ类型包括9-05、16-04、19-12、9-1、16-10、13-05、22-05、13-06和20-02共9个无性系;第Ⅳ类型包括17-06、21-03、23-05、17-05、12-13、18-09、22-10、21-02、12-09和7-01共10个无性系。对上述4大类楸树无性系的MGR、LGR、LGI、不同年份的单株材积进行比较(表 3)可知:第Ⅰ类无性系线性阶段生长量(LGI值)最大,第Ⅱ和Ⅲ类型次之,第Ⅳ类无性系最小,第Ⅰ类是第Ⅳ类的1.86倍。第Ⅰ类无性系1~3 a单株材积生长量高于其他类型单株材积生长量。第Ⅱ类的最大生长速率(MGR)和线性生长速率(LGR)最大,4~7 a单株材积生长量最大。第Ⅱ类7 a的单株材积生长量与第Ⅰ和Ⅱ类型差异不显著,比第Ⅳ类7 a单株材积生长量高80.7%。综上所述,第Ⅰ和Ⅱ类型生长表现较类似,生长速度较快,7 a时单株材积累积都达到较高水平。第Ⅲ类无性系生长表现总体低于第Ⅰ和Ⅱ类型;第Ⅳ类型的线性生长阶段生长量参数和不同年份单株材积均最低,生长表现最差。

Figure 6. Cluster Analysis of C. bungei clones
类群
GoupsMGR/
(m3·a-1)LGR/
(m3·a-1)LGI/
m3单株材积Individual volume / m3 1 a 2 a 3 a 4 a 5 a 6 a 7 a Ⅰ 0.040 9 0.034 0 0.105 3 0.006 0 0.019 1 0.029 6 0.061 8 0.105 9 0.134 0 0.160 1 Ⅱ 0.046 5 0.038 8 0.098 8 0.005 4 0.018 6 0.028 6 0.064 2 0.118 2 0.142 6 0.160 6 Ⅲ 0.037 7 0.031 4 0.099 3 0.005 1 0.016 1 0.024 4 0.053 4 0.096 0 0.117 2 0.147 6 Ⅳ 0.024 2 0.020 2 0.056 6 0.004 4 0.011 9 0.016 4 0.033 0 0.063 1 0.078 7 0.088 9 Table 3. Comparison on individual volume and part growth rhythm parameters of four groups of C. bungei clones
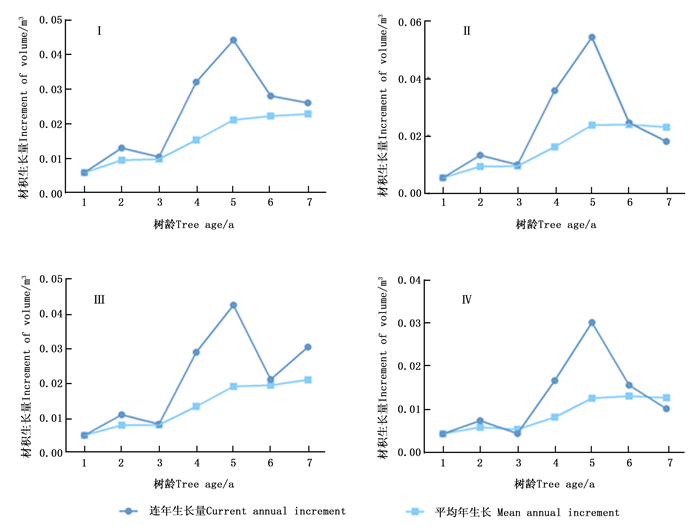
为了解不同类型无性系的单株材积生长规律,分析了其多年的单株材积平均生长量和连年生长量(图 7)。4类无性系单株材积平均年生长量近似“S”型持续增加,3~5 a增长速度近似线性,5~7 a增长速度放缓。4类无性系单株材积连年生长量1~5 a单株材积连年生长量整体增加,在3 a时均有小幅度下降,在5 a达到最大值,其中,第Ⅱ类型连年生长量最大值高于其他类型。特别的是,第Ⅰ和第Ⅲ类无性系在测定的年份中连年单株材积生长量与平均单株材积生长量未有交叉点,而第Ⅱ和Ⅳ类无性系两指标具有交叉点。这表明Ⅰ和Ⅲ类型成熟龄较晚,Ⅱ和Ⅳ类型在第6~7 a左右达到数量成熟龄阶段。
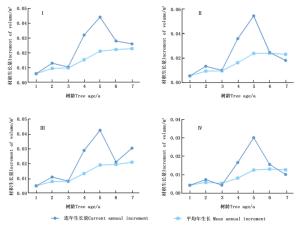
Figure 7. Mean annual increment and current annual increment curves of individual volume of different types of C. bungei clones
2.1. 楸树无性系生长性状方差分析
2.2. 楸树无性系生长性状遗传参数估算
2.3. 楸树无性系连年生长曲线分析
2.4. 楸树无性系单株材积生长曲线拟合及生长特征值相关分析

2.5. 楸树无性系的聚类分析及优良无性系评价
-
遗传与变异是林木遗传改良的基础,高变异和高重复力有利于无性系的选择[17-18]。本研究方差分析结果显示,楸树无性系在不同年份生长量差异显著,说明以无性系水平对楸树生长进行遗传改良具有可行性。楸树胸径和单株材积重复力在6 a之后保持较高值且趋于稳定,赵曦阳等也发现,包括生长性状在内的10个楸树无性系性状重复力超过0.80[19]。与此同时,楸树单株材积变异系数始终保持较高水平(27.84%~35.56%)。
-
本研究通过Logistic曲线对楸树无性系早期单株材积生长进行了拟合,解析了其生长节律参数,结果表明,无性系间生长节律参数存在较大差异。相关分析表明,楸树无性系单株材积生长量与最大生长速率(MGR)、线性生长速率(LGR)、线性生长量(LGI)皆呈极显著的正相关(P<0.01)。同时,聚类分析结果也显示了MGR、LGR和LGI较大的第Ⅰ类、第Ⅱ类无性系7 a单株材积生长量较其他两类无性系单株材积生长量高。这与南方红豆杉[10]、木荷[20]的研究结果类似,揭示了苗木高生长量与线性生长持续时间无关,主要取决于生长速率。而张宋智等[21]对幼龄楸树生长规律进行研究,发现速生期持续时间长短与生物量形成有密切关系。类似的结果还在浙江楠(Phoebe chekiangensis C. B. Shang)[22]、香椿(Toona sinensis (A. Juss.) Roem.)[23]生长节律报道中出现。
数量成熟龄指的是林木在生长过程中,单株材积平均生长量达到最大状态的年龄。一般来讲,在正常的林木生长过程中,单株材积平均生长量与连年生长量都是先上升,达到数量成熟后,缓慢下降[24-25]。楸树单株材积平均年生长量呈“S”型曲线趋势,连年生长量先上升后下降,在5 a达到高峰,截止到本次调查时还未达到数量成熟。在此期间可以加强抚育管理,尽量缩短数量成熟龄到来时间,从而缩短轮伐期,获得较高的经济效益[26]。本研究供试的32个楸树无性系可分为4大类,第Ⅱ类(22-08、22-01、19-27、6-05、20-06、20-01和22-03)和Ⅳ类(17-06、21-03、23-05、17-05、12-13、18-09、22-10、21-02、12-09和7-01)生长较快,依据早期生长数据进行预测,发现在第6~7年达到数量成熟龄,适合于速生用材。第Ⅰ和Ⅲ类在本次研究结束时,单株材积连年生长量和平均生长量尚未出现交点,且在6~7 a生长趋势较高,在未来可能还会出现生长高峰。但考虑到此次分析为楸树无性系早期单株材积生长节律,预测拟合的效果不具较强实际生产指导意义,长周期的生长节律变化有待后期进一步观察研究。
3.1. 楸树无性系生长性状遗传参数估算
3.2. 楸树无性系单株材积早期生长过程分析
-
楸树无性系存在丰富的表型变异,具有较高的选择潜力,而此变异受较强的遗传因素控制,受环境因素影响较小,有利于遗传改良。
楸树无性系间生长节律参数存在较大差异。不同物种、基因型及环境条件是影响林木生长节律的关键因子。因此,揭示林木生长节律,做好速生期间的抚育管理和水肥管理,促进其快速生长尤为重要。
综合各项生长参数,第Ⅱ类楸树虽单株材积生长总量、MGR、LGR、4~7 a单株材积生长量均最大,但早期生长过程分析发现,其单株材积增量呈现出下降趋势,可能后期单株材积累积可能并不理想,不能作为优良的用材无性系;而第Ⅰ类的无性系单株材积总量较大,具有较强的后期生长潜力。由此筛选出6个无性系包括1-1、22-07、19-01、16-05、16-01和16-07。
 DownLoad:
DownLoad: