




-
开花结实是植物进入成熟衰老状态的标志,竹子亦是如此,只是自然界中大多数竹类植物营养生长的时间比较长,要经过数年或数十年甚至更久才会进入性成熟阶段[1-2]。并且竹类植物开花后发生大面积的枯死、竹林进入衰败死亡等开花特性一直威胁着竹类物种的繁衍,而且破坏了大熊猫等一些珍稀濒危动植物的生境,严重威胁着它们的生存[3],同时,也对某些经济竹林造成了毁灭性的打击。目前,水稻(Oyza sativa L.)、小麦(Triticum aestvum L.)、玉米(Zea mays L.)等禾本科其他植物的生殖生物学研究已越来越深入[4-6],而竹子因开花周期漫长,且大多数植株花后成片枯死,结实的情况更为罕见,使得试验材料难以获取,大大限制了其开花结实的生殖生物学的研究,同时使得开花竹林利用以及无性系选育等很多方面的研究进展迟缓[7]。
竹类植物在开花之前有明显的预兆,例如新出土的竹笋矮小或不发笋,竹林整体枯黄。乔士义[8]等研究表明,竹株开花前叶片有发黄的趋势,开花小枝上发生变异生出变态小叶。根据对麻竹(Dendrocalamus latiflorus Munro)[9]植株开花前后动态观察发现,开花的枝条上会出现变态型小叶,代表竹株进入生殖生长的阶段,原应发育为营养轴的潜伏芽转而分化成花序轴;何奇江等[10]对雷竹(Ph. violascens (Carr.) A.et C.Riv.)开花生物学特性进行了研究发现,开花雷竹的花穗绝大多数无叶,可能是开花竹株处于生殖生长时而抑制了营养叶的生长,并且最终雷竹林营养消耗过度,花后竹林衰败死亡。有关竹类植物开花方面的研究主要集中在花期生物学特性[11]、花粉生活力[12]、花粉形态[13]、花粉管的生长[14]等研究上,还有一些关于胚胎学[15]方面的研究。
翠竹(Pleioblastus pygmaeus (Miq.) Nakai)属于地被类竹种,叶片翠绿,极耐修剪,经常运用于园林布置中。20世纪80年代翠竹引入南京林业大学竹种园[16],直至2015年还未有关于其开花结实的相关记载或报道,但近几年竹种园中的翠竹在自然环境中连年开花,为开展竹类植物生殖生物学的研究提供了珍贵的试验材料。本研究以南京林业大学竹类植物园中开花的翠竹为研究对象,综合利用野外定株调查、形态观察以及扫描电镜等方法,系统研究了翠竹花期生物学特性以及翠竹花粉形态等,以期全面了解翠竹有性繁殖的特性,加强竹子开花的基础生物学研究,为竹类植物个体发育过程积累原始材料,也为下一步深入开展竹类植物开花分子机理、遗传转化等深入研究提供参考。
HTML
-
试验地位于江苏省南京市南京林业大学竹种园(118º48′17″ E,32º4′44″ N),土壤属于下蜀系黄壤,属亚热带季风气候,四季分明,春秋短、冬夏长。雨量充沛,平均降水量为971.7 mm,全年积温(10 ℃以上)为 4 897 ℃,年平均气温为15.7 ℃,全年无霜期233 d。年温差较大,7月份平均28 ℃,1月份平均2.2 ℃[17]。
-
在翠竹林中选定3个1 m×1 m样方,连续3年观察调查开花竹林的构成,以了解翠竹开花物候,并对翠竹的开花期进行统计分析,主要记录以下几个时间段:开花初期(25%以下的竹株开花)、开花盛花期(25%~75%的竹株开花)、开花后期(25%以下的竹株开花)、开花末期(少于10%的竹株开花)。
-
2016—2018年连续3年观察翠竹的开花动态,了解翠竹竹株开花和死亡的基本情况。开花期间,定期记录小花开放、花粉散出、内外稃打开与闭合开始和持续的时间。在盛花期任选30根花枝,观察并记录花器官各结构的长度、颜色等花部形态特征以及内、外稃打开时间,并在LEICA S6D体视显微镜下拍照。
-
当翠竹处于盛花期时,采集开花竹株上成熟且未散粉的花药作为材料,先用70% FAA固定花药,扫描电镜制样后,移入Quanta 200环境扫描电镜下观察并照相。
1.1. 研究地点
1.2. 研究方法
1.2.1. 翠竹开花竹林观察
1.2.2. 花序形态特征
1.2.3. 花粉形态观察
-
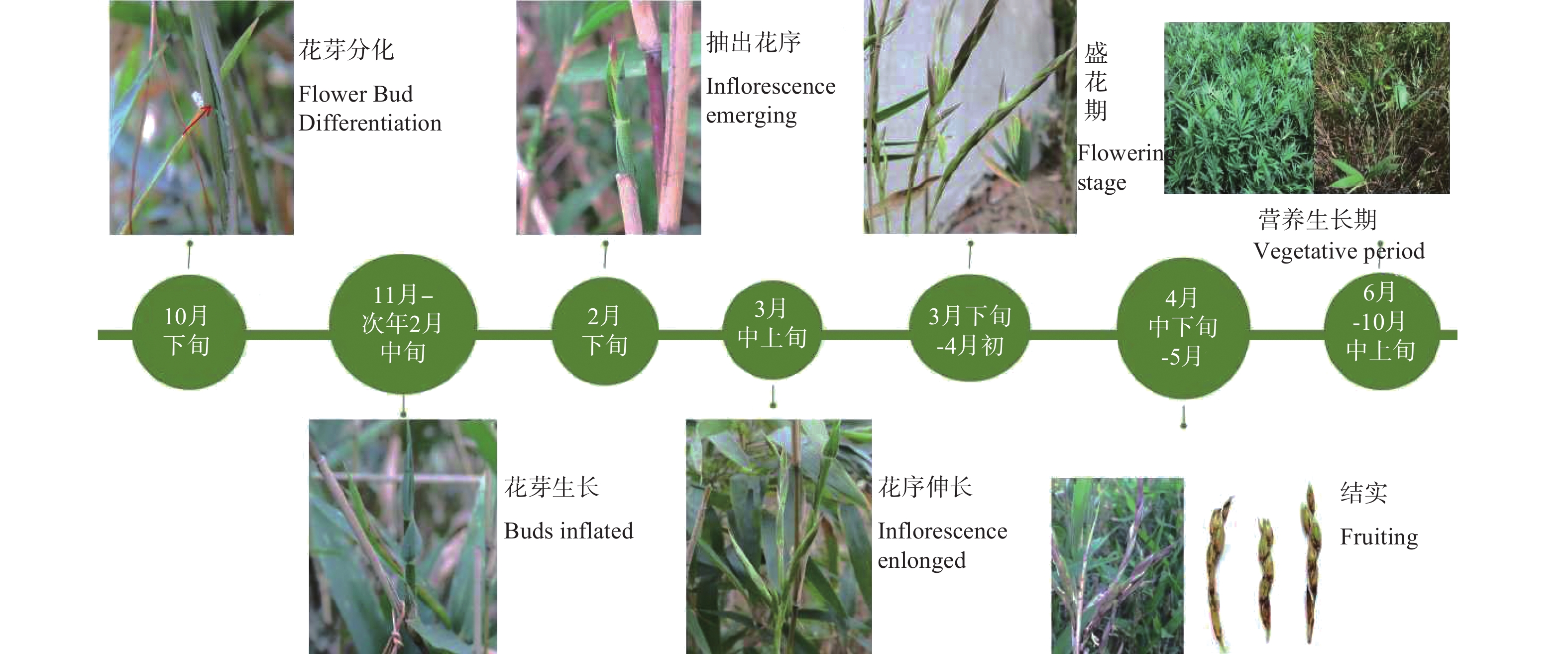
翠竹开花时间较为一致,属于成片开花类型(图1-1)。开花的竹林整体呈枯黄色,每年开花过后部分竹秆死亡,所以在竹林中既有开花的竹子,又有正常不开花的竹子(图1-2),还有已开过花的枯死竹(图1-3)。3月初翠竹竹鞭上会长出新芽,4月份新笋出土长成新竹,6—10月整个翠竹竹林出现返绿现象。

Figure 1. The formation process of inflorescence of Pleioblastus pygmaeus
翠竹花芽的形成到开花跨越两个年度,一般于10月下旬形成花芽(图1-4),次年3月下旬开花。花芽一般是从枝条的下部开始出现,然后从下而上,整个枝条各节都发育为花芽。在花芽发育过程中,前期发育缓慢,即从花芽分化到次年2月气温较低的情况下,花芽发育非常迟缓(图1-5)。3月中上旬,随着温度的升高以及光照的增加,花芽发育速度加快(图1-6,7),到3月中下旬可陆续抽出小穗以及花序,直至花序形态建成(图1-8)。3月下旬至4月上旬为翠竹的盛花期(图1-9),竹林大面积成片开花,并且翠竹盛花期时间比较短,持续时间为2周左右。1个月之后,种子陆续成熟,随机脱落。
翠竹1年之中林相变化(图2)可归纳为:翠竹竹林在10月下旬发生开花转变,开始孕育花芽,竹秆变为枯黄,枝条顶部叶片变黄脱落,直到次年春天3月花序抽出生成小穗,小花形态建成。3月中下旬至4月初为翠竹盛花期,竹林中小花集中开放,4月中上旬大部分小穗开花结束,但在开花期间仍有新生的枝条,其后断断续续有一两个小穗开花,而不开花的枝条上逐渐抽出新叶。1个月之后,种子陆续成熟,果实为颖果。4月份新笋出土长成新竹,整个竹林重新进入营养生长期。
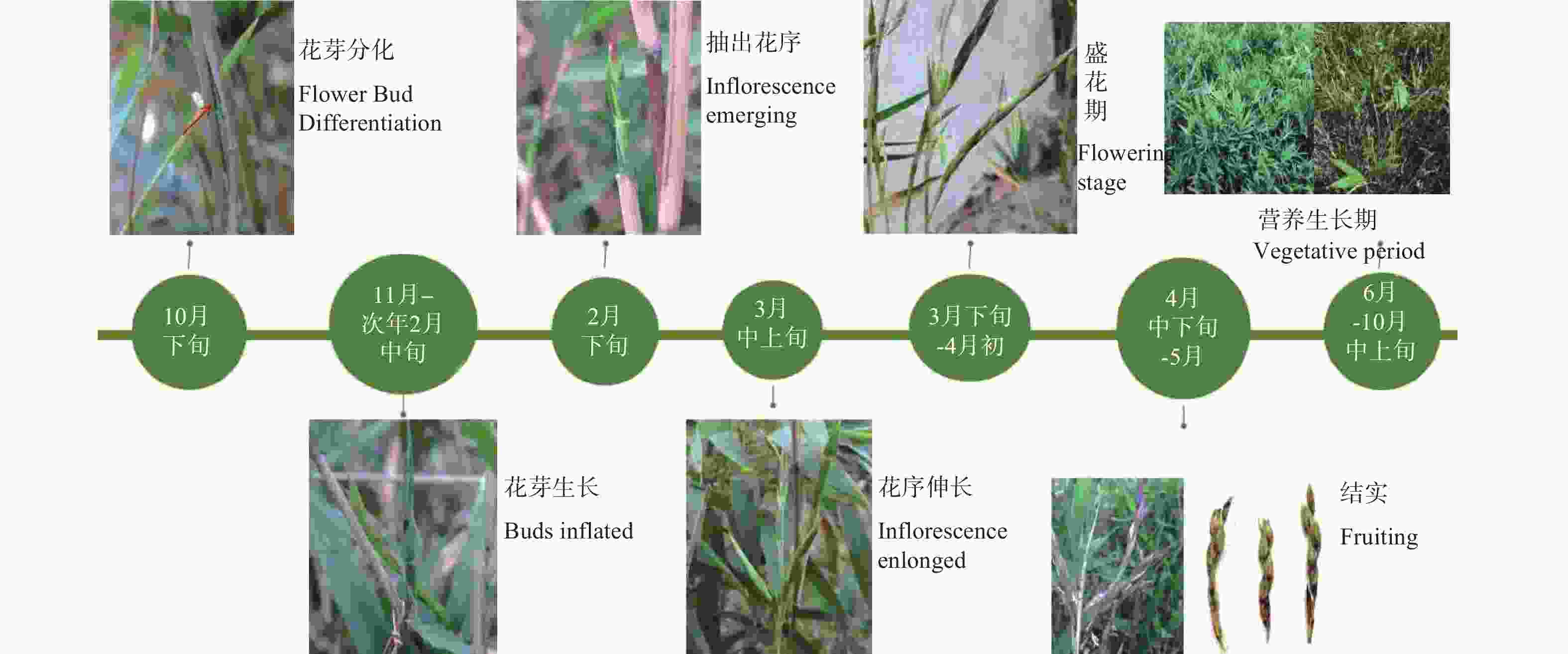
Figure 2. The morphology changes of Pleioblastus pygmaeus in a year
-
翠竹花后死亡与否和开花程度相关,一般开花时无叶或少叶,或过多开花的竹株更容易死亡。根据开花特点,可将开花翠竹竹株分为4个类型:(1)老竹全株开花型;(2)老竹部分开花型;(3)新竹全株开花型;(4)笋芽开花型:
(1)老竹全株开花型:翠竹开花时花芽顶端生出变态小叶,整个植株长满花序,老竹枝条的顶部仅零星分布少量老叶,成片集中开花。花后竹秆呈枯黄色,植株进入半死亡状态。
(2)老竹部分开花型:竹林中这部分翠竹进入生殖年龄后,竹株上只有部分枝条开花,开花后竹株一般不死亡,次年可继续开花。
(3)新竹全株开花型:新竹只有一级侧枝,比正常竹株矮小,新竹成竹展枝时先发出细小竹叶,主枝和一级侧枝的枝条前端都形成花序,外侧的叶片比正常叶片要小。在每年新发竹笋中,都有不开花竹株长成,是竹林自然更新的一种方式。新长成的竹株相对而言较矮小,但是在整个竹林中新竹的比例越来越多,整个竹林逐渐开花停止,从而达到更新复壮。
(4)笋芽开花型:开花翠竹林中,花芽直接从地下冒出,多株簇生在一起,形成无叶型的花序,开花后即死亡(图1-10,11)。根据对地下竹鞭探查,发现从地下竹鞭上的本应发育为笋芽的芽转而形成了花芽,从而形成当年生花序。
-
从表1中可以看出在2016—2018的观察期间,翠竹的花期历时1个月左右,但是盛花期持续较短,一般集中在1星期内完成开花。不同年份的花期略有不同,总体没有明显的差别。
观测项目 Observation items 2016 2017 2018 个别竹株开花
Sporadic bamboos blossom18/03 19/03 15/03 25%竹株开花
25% of bamboos blossom21/03 25/03 24/03 50%及其以上竹株开花
≥50% of bamboos blossom25/03 27/03 26/03 <25%竹株尚在开花
<25% of bamboos blossom29/03 31/03 30/03 开花末期,<10%竹株开花
End of flowering, <10% of bamboos blossom03/04 04/04 01/04 第一朵开花日期
First flowering date13/03 12/03 15/03 盛花期(开花最多的日期)Flowering
stage (Date with the largest number of blossoms)25/03 27/03 26/03 Table 1. The flowering process of Pleioblastus pygmaeus (Date/Month)
-
于2016年3月27日(天气晴朗)标记20株翠竹小花,3个重复,每隔1小时进行定株观测。由表2可以看出翠竹在1天之内随时都可以开花,尤以上午8:00—12:00这段时间开花最多。小花开放数受外界环境影响显著,一般是晴朗的天气开花数较阴雨天多,并且温度越高,开花持续的时间及散粉的速度越快。
时间
Time将要开花的小花数
Number of florets
to Blossom花药已下垂小花数
Number of anther
drooping florets已散粉小花数
Number of scattered
pollen florets7:00—8:00 7±2 2±1 0 8:00—9:00 8±3 3±2 0 9:00—10:00 6±4 6±2 2±1 10:00—11:00 8±4 9±3 4±1 11:00—12:00 6±3 13±4 11±3 12:00—13:00 1±2 10±3 9±3 13:00—14:00 3±2 2±1 6±2 14:00—15:00 4±1 3±1 6±2 15:00—16:00 2±2 1±1 3±1 16:00—17:00 3±1 2±1 2±1 17:00—18:00 2±1 1±1 0 Table 2. The opening process of florets
小花刚开的时候,内、外稃微微张开,只略微看见花药的顶部(图3-1);随着时间的延长,内、外稃张开幅度变大,此时花药露出大半(图3-2);浆片吸水膨大将内外稃完全推开,可见花药全部露出(图3-3);随后花丝不断伸长将花药送出稃片之外,花药逐渐呈下垂状态(图3-4,5),中间露出雌蕊柱头;花药完全下垂时,包裹在内部的柱头完全露出,花药顶部开裂(图3-6,7,8),随后借助风力传粉。花药散粉后,内外稃重新闭合(图3-9)。翠竹单花持续的时间一般只有几小时,花药从稃片包围到伸出稃片之外的时间为4~8 h,花药完成开裂散粉大约需要1~2 h。
于2016年3月28日(天气晴朗)标记了3朵小花,记录小花从片完全闭合到完成散粉所需的时间,开花进程如下:
① 8:50花药微微露出—9:50花药露出大半—10:50花药全部露出—11:00花药趋于下垂—2:50花药全部下垂—13:30花药开裂散粉。
② 8:55花药微微露出—9:55花药露出大半—10:55花药全部露出—11:55花药趋于下垂—12:55花药全部下垂—14:15花药开裂散粉。
③ 9:00花药微微露出—10:00花药露出大半—12:00花药全部露出—13:00花药趋于下垂—14:00花药全部下垂—15:20花药开裂散粉。

Figure 3. The flowering process of Pleioblastus pygmaeus
-
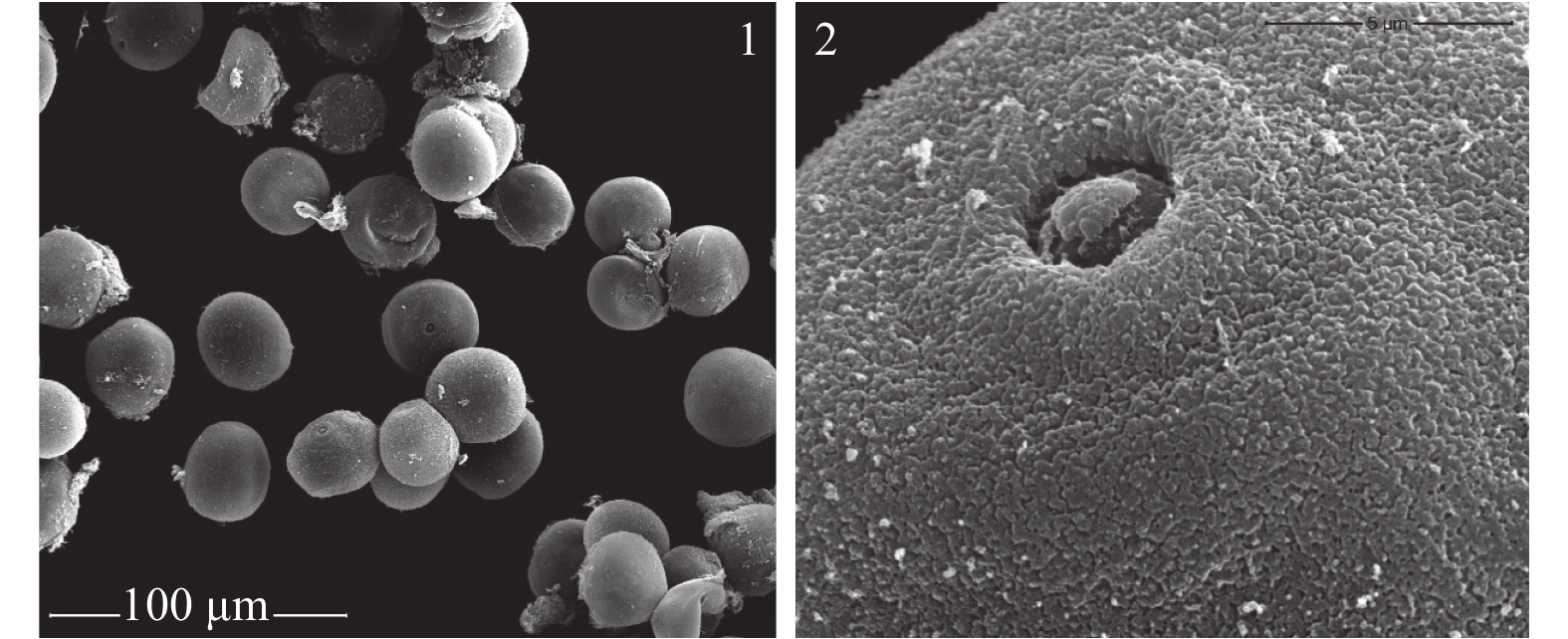
翠竹花粉粒近圆球形(图4-1),单个萌发孔,孔明显,近圆形,萌发孔内凹,孔盖下限(图4-2)。外壁纹饰细颗粒状,外壁极薄,易皱瘪破裂。花粉粒的直径约为30.6~35.4 μm,平均值为33.0 μm。
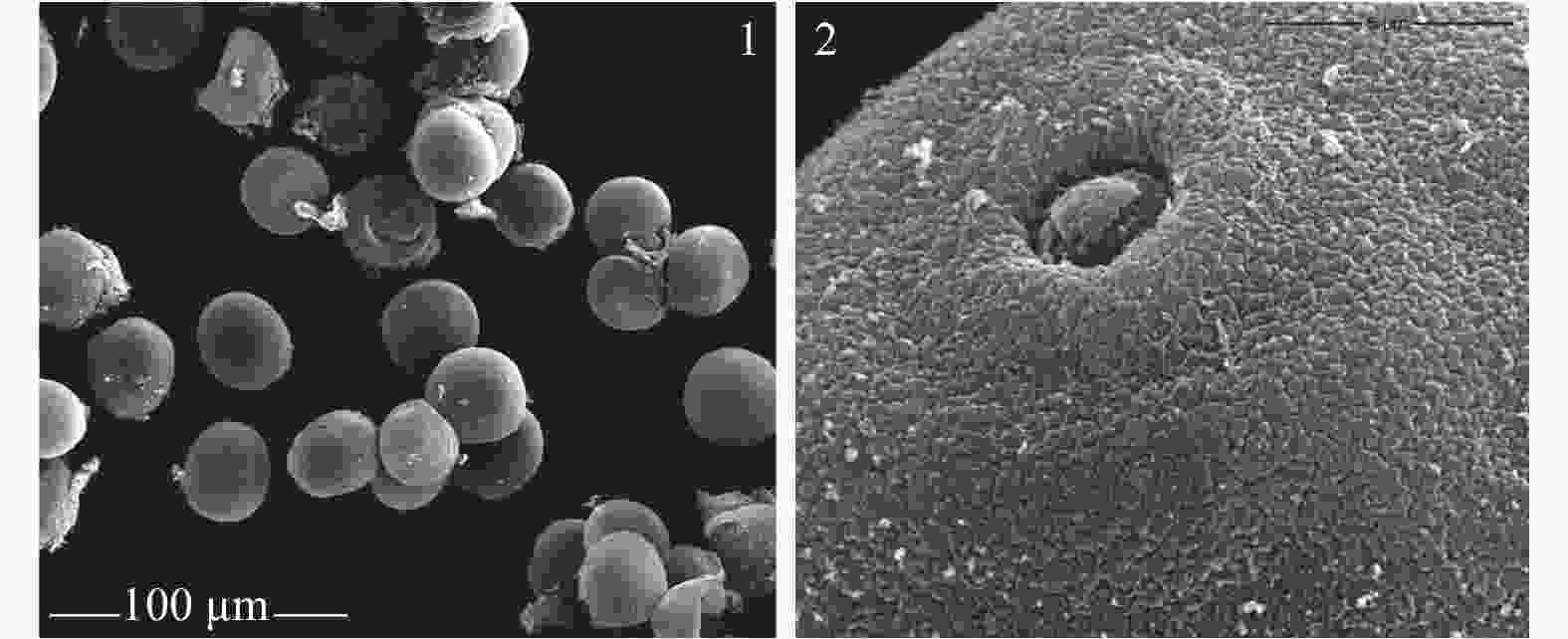
Figure 4. The morphology and germination of pollen of Pleioblastus pygmaeus
2.1. 翠竹开花林相变化
2.2. 开花竹株类型
2.3. 开花动态
2.3.1. 竹林的开花动态
2.3.2. 小花的开花动态

2.4. 花粉形态特征
-
根据对翠竹开花林相的观察发现,翠竹属于成片开花类型,与刚竹属的淡竹 [18]、水竹(Ph. heteroclada Oliver)[19]等竹种开花行为一致,部分半花半叶的竹株可以连续几年开花,全株开花的植株花后即死亡。而丛生竹例如孝顺竹,在开花的竹林中,发现大部分竹丛都是正常生长的,只有少数几株零星开花[20],其他丛生竹林如巨龙竹(Dendrocalamus sinicus Chia et J. L. Sun)[49]、慈竹(Bambusa emeiensis Chia et H.L.Fung)[21]等竹种都表现为零星开花,花期能维持10~15年左右。
-
张文燕等[22]以花柱伸出的程度为依据将竹子分为长花柱型和短花柱型,长花柱型竹种开花时,内外稃片微启不全部张开,柱头细长凸出,能够伸出稃外以接受花粉,如早竹、粉绿竹(Ph. viridiglaucescens (Carr.) A. et C. Riv.)、黄槽石绿竹(Ph. arcana f. luteosulcata C.D.Chu et C.S.Chao)等[17]竹种;后者花柱较短,开花时浆片吸水将稃片撑开露出柱头,如福建茶秆竹(Pseudosasa amabilis var. convexa Z.P.Wang et G.H.Ye)、短穗竹(Semiarundinaria densiflora (Rendle) Wen)、异叶苦竹、月月竹等竹种。翠竹亦属于短花柱型,开花时浆片吸水膨胀,将内外稃推开,露出包裹在内的3裂羽毛状柱头,可以接受飘散的花粉,一段时间后内、外稃重新闭合。
McClure教授根据竹子的小穗基部有无芽的存在发展了竹子的无限制与有限制花序的理论[23]。有限花序基部无休眠芽,开花是一次性的,而无限花序的竹子开花时其小穗基部保留生长点,不断产生新的休眠芽,萌发出新的假小穗。竹子开花时是一次发生或续次发生在分类学上具有重要意义,花序的发育特性是竹子划分超族的依据[24]。2017年林树燕等[16]通过详细研究,认为翠竹花序顶生或侧生,一小穗轴具少数至多枚小穗,属于混合花序。就整个翠竹同一花序而言,一般是位于花序轴顶端的小穗中的小花最先开放,是从顶端开始向基部开放的顺序;而就翠竹同一花小穗而言,一般是小穗基部的小花先开放,然后渐次向顶端开放,开花顺序是从基部开始的。因此,不能将翠竹的开花顺序简单说成单一的从上而下或从下而上。
-
翠竹的花粉近球形,单个萌发孔,孔明显,具明显的孔环和盖,与大部分竹类植物花粉形态描述一致。翠竹的外壁纹饰为细颗粒状,与林树燕[17]报道的短穗竹、异叶苦竹、鹅毛竹等竹种的外壁表面纹饰一致。不同植物的花粉在发育过程中花粉形态都比较稳定,进化上最具保守性,因此了解翠竹花粉形态对于竹子的分类研究具有重要意义[25]。测得翠竹的花粉粒直径为33.0 μm,小于毛竹的(61.0 μm)和鹅毛竹的(40.22 μm),但是与异叶苦竹的(27.86 μm)相近。刚竹属竹种花粉属于大型花粉,平均直径均在50 μm以上;而苦竹属的花粉属于中型花粉,平均直径多在50 μm以下[26]。
-
翠竹原产日本,最初的学名由荷兰植物学家 Friedrich Anton Wilhelm Miquel 1866年命名,其基本名是Bambusa pygmaea Miq[27];由于该竹子不是丛生竹,英国植物学家Algernon Bertram Freeman-Mitford于1896年将翠竹移至散生竹的青篱竹属,学名为Arundinaria pygmaea (Miq.) Mitf;1902年由Ascherson,Paul Friedrich August等人更名为Arundinaria pygmaea (Miq.) Asch. & Graebn[28];1913年法国植物学家E. G. Camus认为该竹子的分枝特性等性状不同于青篱竹属,而将翠竹移至赤竹属,学名为Sasa pygmaea (Miq.) E. G. Camus[29],1932年日本植物分类学家Nakai将翠竹移至苦竹属(也称大明竹属),学名为Pleioblastus pygmaeus (Miq.) Nakai[30];1942年,Nakai 发表了新属Nipponocalamus,将翠竹移至该新属,学名为Nipponocalamus pygmaeus (Miq.)Nakai[31];1978年,日本植物学家认为该新属不成立,于是将翠竹又移至苦竹属,学名为Pleioblastus pygmaeus (Miq.) Nakai,该竹种被引种到我国后,先后被置于赤竹属[32-33]和青篱竹属[34]。鉴于东亚是否有青篱竹属竹种的分布,国内学术界还存在很大争议。2017年林树燕[16]认为赤竹属的竹种是6个雄蕊,而翠竹只有3个雄蕊,于是将翠竹移至苦竹属(也称大明竹属),学名为Pleioblastus pygmaeus (Miq.) Nakai。
3.1. 开花现象以及开花类型
3.2. 花序的特点
3.3. 花粉形态
3.4. 翠竹分类学的讨论
-
翠竹开花时间一致,属于成片开花类型,翠竹花芽的形成到开花跨越2个年度,一般于10月下旬形成花芽,次年3月下旬开花,3月下旬至4月上旬为翠竹的盛花期,持续时间为1周左右;翠竹在1天之内随时都可以开花,上午(上午8:00—12:00)开花最多且受温度影响显著。1个月之后,种子陆续成熟,随机脱落,每年开花过后部分竹秆死亡。翠竹花后死亡与否和开花程度相关,一般开花时无叶或少叶,或过多开花的竹株更容易死亡。根据开花特点,可将开花翠竹竹株分为4个类型:(1)老竹全株开花型;(2)老竹部分开花型;(3)新竹全株开花型;(4)笋芽开花型。翠竹单花持续的时间一般只有几小时,花药从稃片包围到伸出稃片之外的时间为4~8 h,花药完成开裂散粉大约需要1~2 h。翠竹花粉粒近圆球形,单个萌发孔,孔明显,近圆形,萌发孔内凹,孔盖下限。外壁纹饰细颗粒状,外壁极薄,易皱瘪破裂。花粉粒的直径约为30.6~35.4 μm,平均值为33.0 μm.
翠竹花后,能收获一定数量的种子,在自然状态下的开花翠竹林复壮能力目前还在观察。
 DownLoad:
DownLoad: