



-
马尾松(Pinus massoniana Lamb.)在我国松属树种中分布最广泛,具有适应性强、经济价值高等特点。它分泌的大量以萜类化合物为主的次生代谢物称为松脂,主要由单萜、倍半萜和二萜组成,贮存在针叶树的根、茎、叶和球果中[1]。松脂是松香、松节油工业的基础原料,松香、松节油深加工产品又可作为其他相关工业的中间原料,广泛用于油墨、胶黏剂、香料、涂料、纺织、电子印染和医药等工业生物产品中[2]。
萜类化合物是植物次生代谢产物中最大的一类天然化合物,广泛存在于动植物体内,目前已研究报道过的萜类化合物有5万多种。萜类化合物有重要的生物学功能和应用价值,在植物体内,单萜和倍半萜可看作挥发油的成分,大多数具有香气,是食品、医药、化妆品的重要原料。超过20个碳的萜类物质一般为非挥发性物质,树脂酸和赤霉素是二萜中最常见的物质,其中,树脂酸在马尾松中占所有代谢物的70%以上[3]。1-脱氧-d-木酮糖-5-磷酸还原异构酶(DXR)是马尾松萜类化合物合成途径中的重要限速酶,它催化1-脱氧-d-木酮糖-5-磷酸(DXP)发生还原反应生成2-C-甲基-D-赤藓糖醇-4-磷酸(MEP)[4],同时DXP也是维生素B1和B6的前体物质[5-6],故该催化反应被认为是MEP途径中的关键调控位点。目前,已从多种植物中克隆出DXR基因,并对其功能作了相关研究;但关于DXR基因密码子偏好性的研究较少,仅在豆科植物中有报道,李凌烜等发现豆科植物DXR基因对A或U结尾的密码子表现出较强的偏好性[7]。
密码子是自然界中遗传信息准确识别和传递的基本载体,在生物遗传和变异中起着关键作用。构成天然蛋白质的20种氨基酸中除甲硫氨酸和色氨酸外,其余氨基酸都对应2~6种密码子,编码同一种氨基酸的密码子被称为同义密码子[8]。研究发现,不同生物体在编码氨基酸时对密码子的选择并不是完全随机的,而是偏好使用某些特定的同义密码子[9];另外,同种生物中不同基因对密码子的使用频率也存在差异,这种现象即为密码子使用偏好性[10]。同义密码子的选择使用不改变氨基酸序列,有利于翻译的准确性,但可能会对蛋白表达水平产生影响[11-12]。密码子偏好性会影响蛋白质翻译的速度和折叠、mRNA水平的转录调控及外源基因的表达量[13-14]。如果外源基因密码子与宿主系统密码子的使用频率相比有明显差异,将会影响外源基因的翻译与表达[15]。因此,分析密码子的偏好性对研究基因功能、蛋白结构和提高外源基因表达效率有重要意义。目前,科学界尚无关于马尾松PmDXR基因在密码子偏好性方面的报道,对其相关功能尚不清楚。本研究将分析马尾松PmDXR基因密码子使用偏好性,讨论其形成的主要因素,再与拟南芥(Arabidopsis thaliana (L.) Heynh.)、烟草(Nicotiana tabacum L.)、酿酒酵母(Saccharomyces cerevisiae Meyen. ex Hansen.)和大肠杆菌(Escherichia coli (Migula.) Castellani & Chalmers.)基因组密码子进行比较分析,以期为马尾松PmDXR功能验证相关研究提供基础数据。
HTML
-
马尾松PmDXR基因序列由本实验室克隆获得,该序列全长1 994 bp(GenBank登录号为MK969119.1),其开放阅读框(ORF)为1 584 bp,编码527个氨基酸。银杏(Ginkgo biloba L.)、赤松(Pinus densiflora Sieb. et Zucc.)、火炬松(Pinus taeda L.)等38个物种的DXR基因CDS序列来自GenBank(
https://www.ncbi.nlm.nih.gov/ ),登录号见表1。本文所采用的拟南芥、烟草、酿酒酵母和大肠杆菌的基因组密码子使用频率数据来自密码子使用数据库(http://www.kazusa.or.jp/codon )。类别
Classes物种
Species登录号
Genbank
accession
numberumberA3s C3s G3s U3s ENc CAI GC3s/
%GC/
%裸子植物 银杏 Ginkgo biloba AY494186.1 0.350 0.177 0.256 0.435 51.05 0.200 36.82 45.26 赤松 Pinus densiflora EU439294.1 0.390 0.178 0.231 0.425 49.32 0.192 34.58 44.17 火炬松 Pinus taeda EU862299.1 0.396 0.171 0.226 0.426 47.89 0.188 33.75 44.24 马尾松 Pinus massoniana MK969119.1 0.410 0.186 0.222 0.402 48.93 0.184 34.47 43.75 东北红豆杉 Taxus cuspidata Sieb. et Zucc. AY575140.1 0.383 0.176 0.242 0.420 47.99 0.216 35.56 44.70 单子叶植物 枫茅 Cymbopogon winterianus Jowitt. KJ749651.1 0.346 0.228 0.269 0.374 55.97 0.203 41.60 47.69 盾叶薯蓣 Dioscorea zingiberensis C. H. Wright. KY131955.1 0.326 0.257 0.252 0.384 53.92 0.208 42.89 47.49 浦竹仔 Indosasa hispida McClure. KX819309.1 0.319 0.229 0.301 0.380 53.41 0.235 43.64 47.88 水稻 Oryza sativa L. AF367205.1 0.330 0.240 0.279 0.375 54.75 0.214 43.46 48.03 双子叶植物 砂仁 Amomum villosum Lour. FJ459894.1 0.349 0.221 0.245 0.415 52.84 0.193 38.90 46.23 金鱼草 Antirrhinum majus L. AY770406.1 0.280 0.224 0.281 0.450 54.68 0.227 41.53 46.12 沉香 Aquilaria sinensis (Lour.) Spreng KF498704.1 0.264 0.221 0.291 0.450 52.71 0.227 42.17 47.11 拟南芥 Arabidopsis thaliana AF148852.1 0.326 0.178 0.250 0.473 49.92 0.212 35.98 44.56 黄花蒿 Artemisia annua L. AF182287.2 0.313 0.190 0.241 0.474 51.65 0.229 36.36 44.05 艾草 Artemisia argyi Lévl. et Van. MG780999.1 0.323 0.203 0.235 0.455 50.48 0.228 37.00 44.33 苍术 Atractylodes lancea (Thunb.) DC. MG859909.1 0.263 0.265 0.282 0.421 54.13 0.244 44.94 47.19 胭脂树 Bixa orellana L. KT358987.1 0.279 0.205 0.261 0.483 52.04 0.233 38.89 45.62 积雪草 Centella asiatica (L.) Urban. JQ965955.1 0.298 0.214 0.255 0.461 50.52 0.217 38.95 45.54 樟树 Cinnamomum camphora (L.) Presl KU886266.1 0.297 0.220 0.296 0.413 53.46 0.208 42.68 47.13 枇杷 Eriobotrya japonica (Thunb.) Lindl. JX089590.1 0.295 0.217 0.256 0.446 55.01 0.216 39.66 46.84 栀子花 Gardenia jasminoides Ellis. KY631919.1 0.326 0.233 0.229 0.439 52.50 0.211 38.57 45.56 滇龙胆草 Gentiana rigescens Franch. ex Hemsl. KF941189.1 0.276 0.196 0.282 0.475 50.44 0.227 39.37 44.98 橡胶树 Hevea brasiliensis (Willd. ex A. Juss.) Muell. Arg. DQ437514.1 0.323 0.180 0.229 0.490 52.05 0.200 34.53 44.42 薰衣草 Lavandula angustifolia Mill. JX630151.1 0.217 0.307 0.329 0.382 53.97 0.209 51.68 49.23 番茄 Lycopersicon esculentum Mill. AF331705.2 0.309 0.202 0.264 0.444 49.67 0.191 38.87 45.59 桑 Morus alba L. KX387390.1 0.297 0.262 0.258 0.403 55.14 0.218 43.43 47.53 水仙 Narcissus tazetta var. chinensis Roem. GU574805.1 0.313 0.196 0.273 0.439 51.40 0.204 39.28 46.28 木樨 Osmanthus fragrans (Thunb.) Lour. KX400843.1 0.313 0.218 0.240 0.458 53.99 0.213 38.11 45.19 夏枯草 Prunella vulgaris L. MK272811.1 0.269 0.309 0.276 0.372 59.49 0.235 48.52 49.16 西洋梨 Pyrus communis L. KM030333.1 0.290 0.213 0.273 0.442 54.71 0.210 40.55 46.99 萝芙木 Rauvolfia verticillata (Lour.) Baill. DQ779286.1 0.321 0.203 0.233 0.475 50.94 0.219 36.21 44.14 地黄 Rehmannia glutinosa (Gaert.) Libosch. ex Fisch. et Mey. KX058461.1 0.289 0.215 0.273 0.453 55.23 0.206 40.42 45.89 玫瑰 Rosa rugosa Thunb. JX518616.1 0.324 0.190 0.248 0.452 52.32 0.201 37.00 45.88 丹参 Salvia miltiorrhiza Bunge. DQ991431.1 0.274 0.269 0.309 0.388 55.65 0.205 47.37 48.35 半枝莲 Scutellaria barbata D.Don. MK035042.1 0.265 0.251 0.285 0.425 56.63 0.209 44.26 47.69 阿育魏实 Trachyspermum ammi (L.) Sprague MG762013.1 0.320 0.175 0.235 0.500 48.84 0.216 34.11 43.65 雷公藤 Tripterygium wilfordii Hook. f. KJ174341.1 0.301 0.179 0.261 0.496 53.61 0.232 36.38 44.40 睡茄 Withania somnifera (L.) Dunal GQ921844.1 0.297 0.199 0.274 0.446 51.30 0.197 39.58 45.96 Table 1. Preference related parameters of DXR gene codons among different species
-
整理38个物种DXR基因的CDS序列,借助CodonW1.4.2软件计算密码子的使用特性参数,包括A3s、C3s、U3s、G3s、相对同义密码子使用度(RSCU)、密码子适应指数(CAI)和有效密码子数(ENc)等。运用EMBOSS中的CUSP和CHIP在线程序计算38个物种DXR基因密码子G和C总含量(GC)、密码子第3位上的GC含量(GC3s)、使用频率(Frequency)等;同时运用SPSS20.0软件分析38个物种DXR基因密码子使用特性参数的相关性,再进一步对DXR基因密码子进行中性绘图分析、奇偶偏好性(PR2-plot)分析以及ENc-plot分析。
CAI值范围为0~1,越接近1表明偏好性越强,而越接近0则表明偏好性越弱[16]。ENc值范围为20~61,可反映特定基因密码子家族中同义密码子非均衡使用的偏好程度[17]。RSCU为某一密码子的使用频率与其无偏性时期望频率的比值。一般,若RSCU值小于1,表明该密码子使用偏好性较低;若其值等于1,表明该密码子无偏好性;若其值大于1,则表明该密码子使用偏好性较强[18]。
-
通过MEGA5.1软件中的邻接法(Neighbor-Joining),基于38个物种DXR基因的CDS序列构建系统进化树,检验方法为Bootstrap method,检验次数为1 000,Gaps处理方法为Complete deletion。并利用在线绘图软件EvolView (
https://evolgenius.info//evolview-v2/#login )可视化展示进化树。将每个物种的DXR基因作为一个对象,以59个同义密码子的(去除终止密码子TAA、TAG和TGA,起始密码子ATG和编码甲硫氨酸的密码子TGG)的RSCU值为变量对密码子使用偏好性进行分析。
1.1. 序列来源
1.2. 数据分析方法
1.2.1. DXR基因同义密码子偏好性分析
1.2.2. 基于DXR基因CDS序列和密码子使用偏好性的聚类分析
-
利用CondonW程序分析马尾松PmDXR基因密码子的CAI值、ENc值,同时运用EMBOSS中的CUSP在线程序计算总GC含量以及GC3s有效密码子数。结果(表1)显示:PmDXR基因中密码子的GC和GC3s分别为43.75%和34.47%,说明其密码子比较偏好以A/T结尾。ENc值为35可作为区分偏好性强弱的标准[19],PmDXR基因密码子ENc值为48.93,说明PmDXR基因在编码氨基酸时密码子使用偏好性较弱,其表达水平可能相对较低。CAI值为0.184,进一步表明PmDXR基因对密码子的选择偏好性较弱。
-
CUSP和CodonW计算结果(表2)表明:有25个密码子的RSCU值大于1(除终止密码子TGA),其中,GCA、GCT、AGA、AAT、GGA、CCA、CCT、TCA、TCT、ACA、TAT、GTT共12个密码子的RSCU值大于1.5,且编码精氨酸的AGA与编码丝氨酸的TCT密码子RSCU值分别为3.60、2.17,说明PmDXR基因中共有25个密码子具有偏好性,其中,有12个密码子偏好性较强,2个偏好性极强。TGC、TGT、TCC、TGG共4个密码子(除起始密码子ATG)的RSCU值等于1,说明PmDXR基因中有4个密码子没有偏好性。29个密码子的RSCU值小于1,说明PmDXR基因对这些密码子的使用偏好性较弱。编码Arg的密码子CGC和CGT的RSCU值为0,说明它们可能不参与马尾松PmDXR基因的翻译过程。此外,马尾松中NCG型密码子的RSCU值(GCG: 0.15,CCG: 0.24,TCG: 0.33,ACG: 0.15)较低,推断马尾松可能是一种甲基化程度较高的植物。
密码子
Codon氨基酸
Amino acid比例
Fraction频率
Frequency数目
Number相对密码子使用度
RSCU密码子
Codon氨基酸
Amino acid比例
Fraction频率
Frequency数目
Number相对密码子使用度
RSCUGCA Ala 0.377 37.879 20 1.51 CTT Leu 0.235 22.727 12 1.41 GCC 0.189 18.939 10 0.75 TTA 0.176 17.045 9 1.06 GCG 0.038 3.788 2 0.15 TTG 0.196 18.939 10 1.18 GCT 0.396 39.773 21 1.58 AAA Lys 0.543 35.985 19 1.09 AGA Arg 0.600 22.727 12 3.60 AAG 0.457 30.303 16 0.91 AGG 0.150 5.682 3 0.90 ATG Met 1.000 18.939 10 1.00 CGA 0.150 5.682 3 0.90 TTC Phe 0.316 11.364 6 0.63 CGC 0.000 0.000 0 0.00 TTT 0.684 24.621 13 1.37 CGG 0.100 3.788 2 0.60 CCA Pro 0.441 28.409 15 1.76 CGT 0.000 0.000 0 0.00 CCC 0.088 5.682 3 0.35 AAC Asn 0.222 3.788 2 0.44 CCG 0.059 3.788 2 0.24 AAT 0.778 13.258 7 1.56 CCT 0.412 26.515 14 1.65 GAC Asp 0.385 18.939 10 0.77 AGC Ser 0.056 3.788 2 0.33 GAT 0.615 30.303 16 1.23 AGT 0.083 5.682 3 0.50 TGC Cys 0.500 7.576 4 1.00 TCA 0.278 18.939 10 1.67 TGT 0.500 7.576 4 1.00 TCC 0.167 11.364 6 1.00 CAA Gln 0.600 11.364 6 1.20 TCG 0.056 3.788 2 0.33 CAG 0.400 7.576 4 0.80 TCT 0.361 24.621 13 2.17 GAA Glu 0.471 30.303 16 0.94 ACA Thr 0.500 24.621 13 2.00 GAG 0.529 34.091 18 1.06 ACC 0.269 13.258 7 1.08 GGA Gly 0.400 26.515 14 1.60 ACG 0.038 1.894 1 0.15 GGC 0.143 9.470 5 0.57 ACT 0.192 9.470 5 0.77 GGG 0.200 13.258 7 0.80 TGG Trp 1.000 15.152 8 1.00 GGT 0.257 17.045 9 1.03 TAC Tyr 0.091 1.894 1 0.18 CAC His 0.563 17.045 9 1.13 TAT 0.909 18.939 10 1.82 CAT 0.438 13.258 7 0.88 GTA Val 0.244 20.833 11 0.98 ATA Ile 0.390 30.303 16 1.17 GTC 0.089 7.576 4 0.36 ATC 0.146 11.364 6 0.44 GTG 0.222 18.939 10 0.89 ATT 0.463 35.985 19 1.39 GTT 0.444 37.879 20 1.78 CTA Leu 0.157 15.152 8 0.94 TAA TER 0.000 0.000 0 0.00 CTC 0.098 9.470 5 0.59 TAG 0.000 0.000 0 0.00 CTG 0.137 13.258 7 0.82 TGA 1.000 1.894 1 3.00 注:下划线表示该密码子RSCU > 1。
Note: The underline means that the value of RSCU > 1.Table 2. RSCU of PmDXR gene in P. massoniana
-
38个物种DXR基因密码子的A3s、C3s、U3s、G3s、ENc值、CAI值、GC和GC3s含量见表1。分析比较A3s、C3s、U3s和G3s发现,38种植物DXR的A3s和U3s普遍高于C3s和G3s,表明DXR编码区优先使用A或U结尾的同义密码子。38个物种DXR基因的ENc值为47.89~59.49,平均值为52.59,其中,裸子植物DXR的ENc平均值为49.04,单子叶植物DXR的ENc平均值为54.51,双子叶植物DXR的ENc平均值为52.94,表明裸子植物DXR基因的密码子偏好性相对单子叶和双子叶植物较高。由此可见,DXR基因在各物种中表现出较弱的密码子偏好性,表达水平普遍不高,但物种间还是存在一定差异。38个物种DXR基因的CAI值介于0.184~0.244,进一步说明DXR基因的密码子偏好性较弱。裸子植物DXR的GC含量介于43.75%~45.26%,GC3s介于33.75%~36.82%。单子叶植物DXR的GC含量介于47.49%~48.03%,GC3s介于41.60%~43.64%。双子叶植物DXR的GC含量介于43.65%~49.23%,GC3s介于34.11%~51.68%。整体看,裸子植物DXR的GC含量和GC3s低于双子叶植物和单子叶植物,且38种植物DXR基因均偏好以A/T结尾的密码子。
-
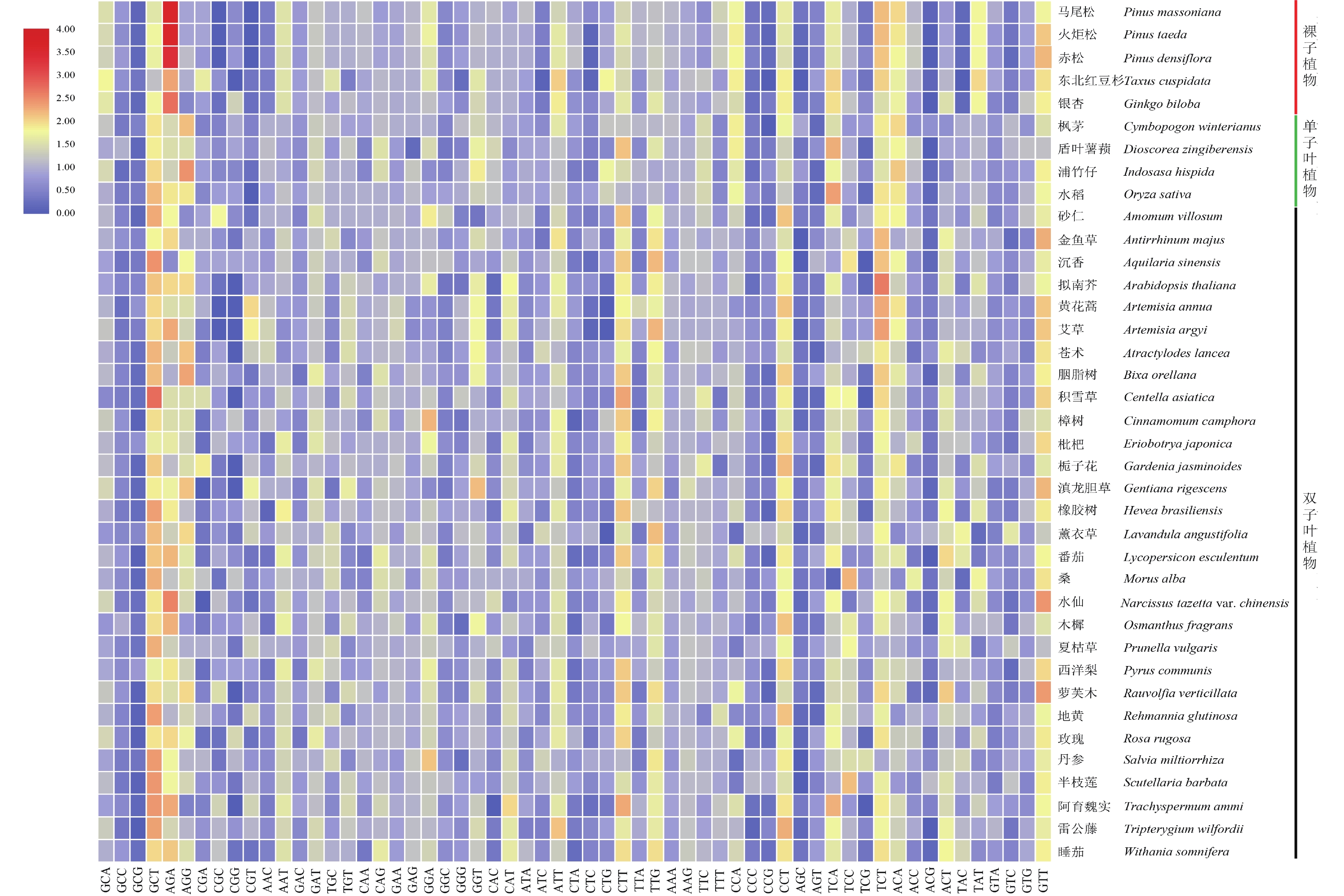
RSCU是衡量密码子偏好性的重要指标,用38个物种DXR基因密码子的RSCU值制作热图(图1),结果显示:裸子植物与单子叶植物和双子叶植物DXR的RSCU值存在一定差异。整体看,裸子植物DXR中RSCU值大于1.5的密码子较单子叶植物和双子叶植物多,东北红豆杉RSCU大于1.5的密码子有16个,丹参和夏枯草只有6个。说明裸子植物密码子使用偏好性比其他植物强,这与基于ENC分析的结果相同。在所分析的绝大部分物种的DXR基因密码子中,GCT、AGA、GAT、ATT、CTT、TTG、CCT、TCT、GTT的RSCU值都大于1,说明这几个密码子出现频率较高,属于共同偏好使用的密码子,且这些密码子偏向以A/T结尾。除此之外,通过对马尾松、火炬松、赤松DXR的密码子分析发现,AGA、TCT的RSCU值均大于2,由此推测松科植物对这2个密码子的偏好性极强。
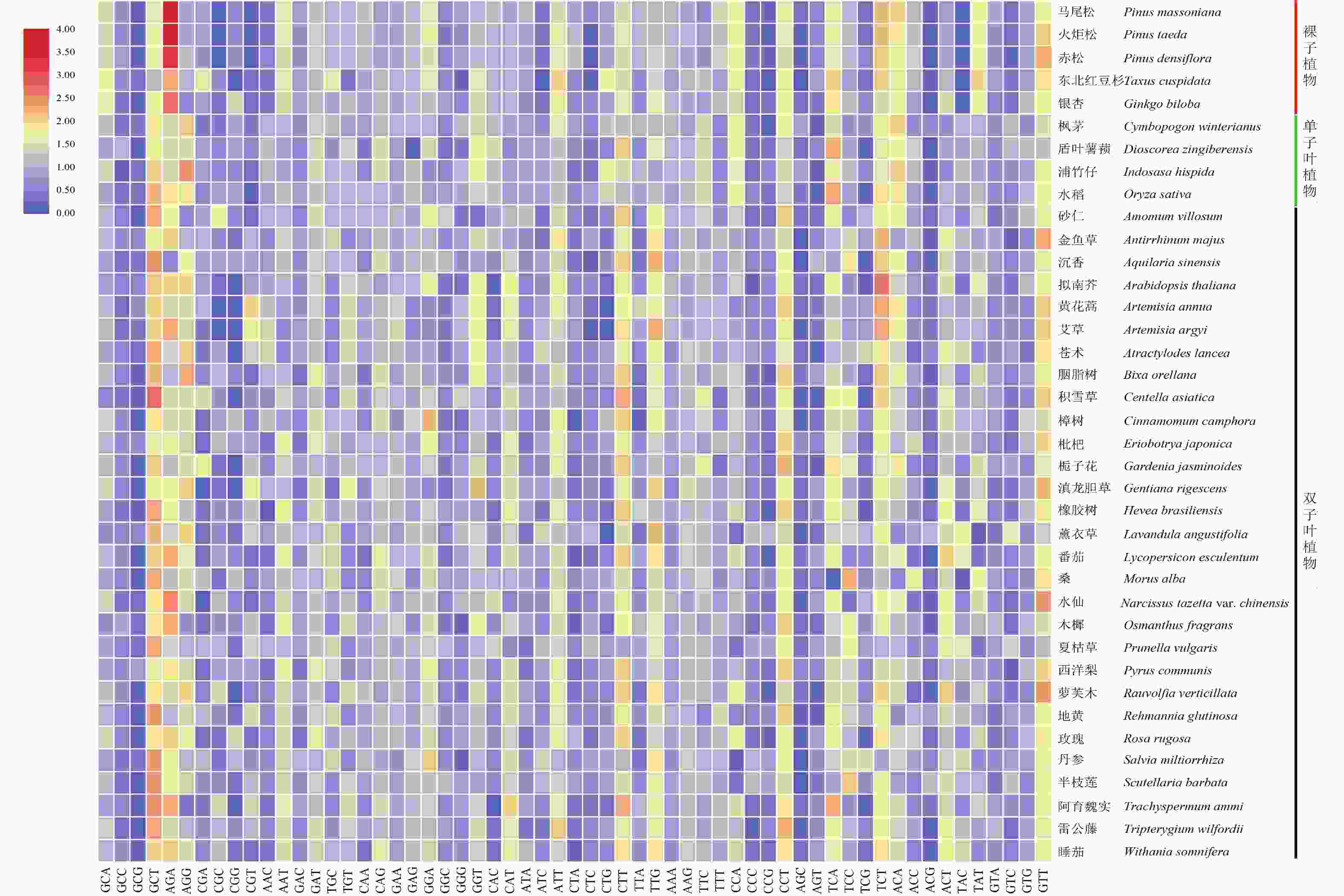
Figure 1. Heat map of RSCU values of DXR gene from 38 species
-
用SPSS20.0对DXR密码子成分 GC、GC3s、ENc、CAI、密码子偏爱指数(CBI)、最优密码子使用频率(Fop)、A3s、U3s、C3s、G3s进行两两Pearson关联分析(表3)。结果显示:ENc 和GC、GC3s、CAI、CBI、Fop、C3s、G3s参数呈显著或极显著正相关,与U3s、A3s 参数呈极显著负相关,说明DXR密码子偏好性越强,其碱基中A、T含量越高。
参数
parameterGC GC3s ENc CAI CBI Fop A3s U3s C3s G3s GC 1.000 GC3s 0.945** 1.000 ENc 0.796** 0.763** 1.000 CAI 0.191 0.313 0.331* 1.000 CBI 0.587** 0.671** 0.590** 0.730** 1.000 Fop 0.572** 0.663** 0.580** 0.756** 0.997** 1.000 A3s −0.583** −0.719** −0.595** −0.544** −0.659** −0.658** 1.000 · U3s −0.733** −0.651** −0.443** 0.162 −0.248 −0.236 −0.053 1.000 C3s 0.877** 0.940** 0.776** 0.302 0.662** 0.662** −0.625** −0.661** 1.000 G3s 0.791** 0.851** 0.556** 0.299 0.538** 0.524** −0.741** −0.419** 0.628** 1.000 注:**表示在0.01水平上显著相关,*表示在0.05水平上显著相关。
Notes: ** means significant correlations at 0.01 level, * means significant correlations at 0.01 leve.Table 3. Correlation analysis of DXR related parameters
-
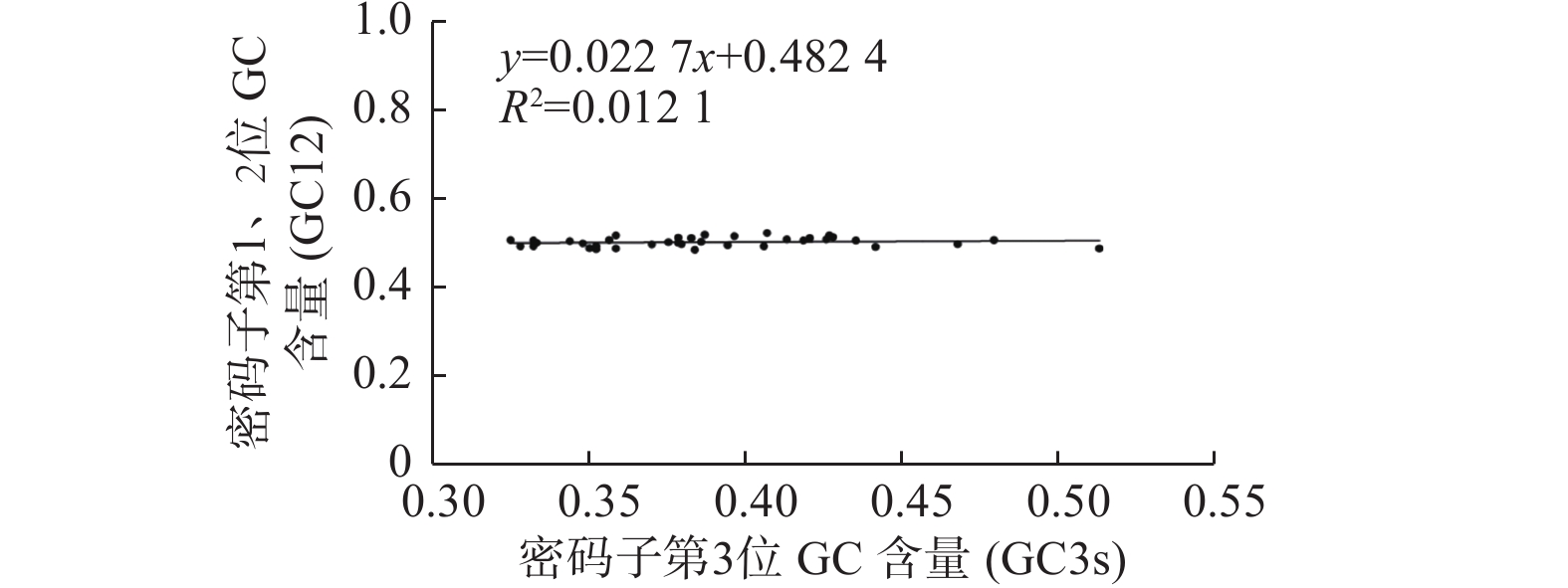
密码子第1、2位GC含量平均值(GC12)与GC3s相关性分析表明相关不显著,对不同物种DXR基因进行中性分析(图2)发现:GC3s的范围为33.75%~51.68%,GC12的范围为47.79%~50.74%。GC12的范围相对较小,绝大部分基因靠近回归线分布。GC12与GC3s的线性回归系数为0.0227,远小于1,说明DXR基因密码子第1、2位碱基与第3位碱基组成差异较大。由此推断,在进化过程中DXR基因密码子的偏好性主要受自然选择影响[20]。
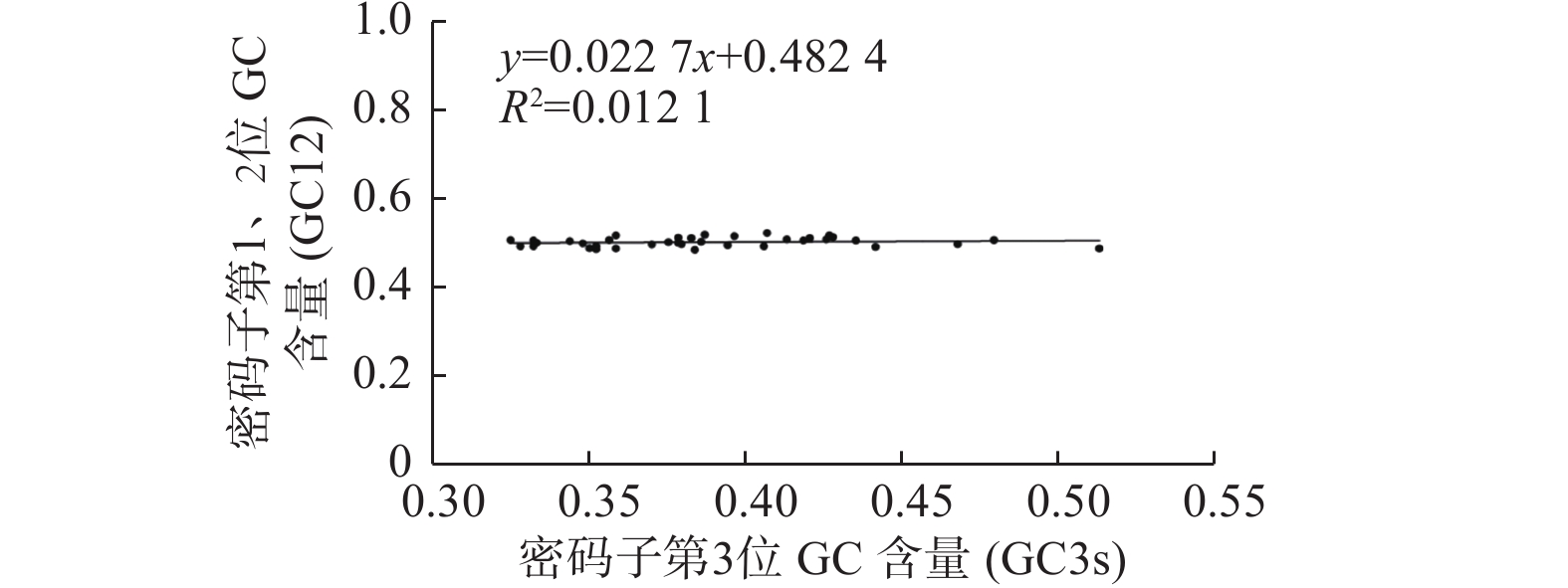
Figure 2. Neutrality plot of DXR genes in different species
-
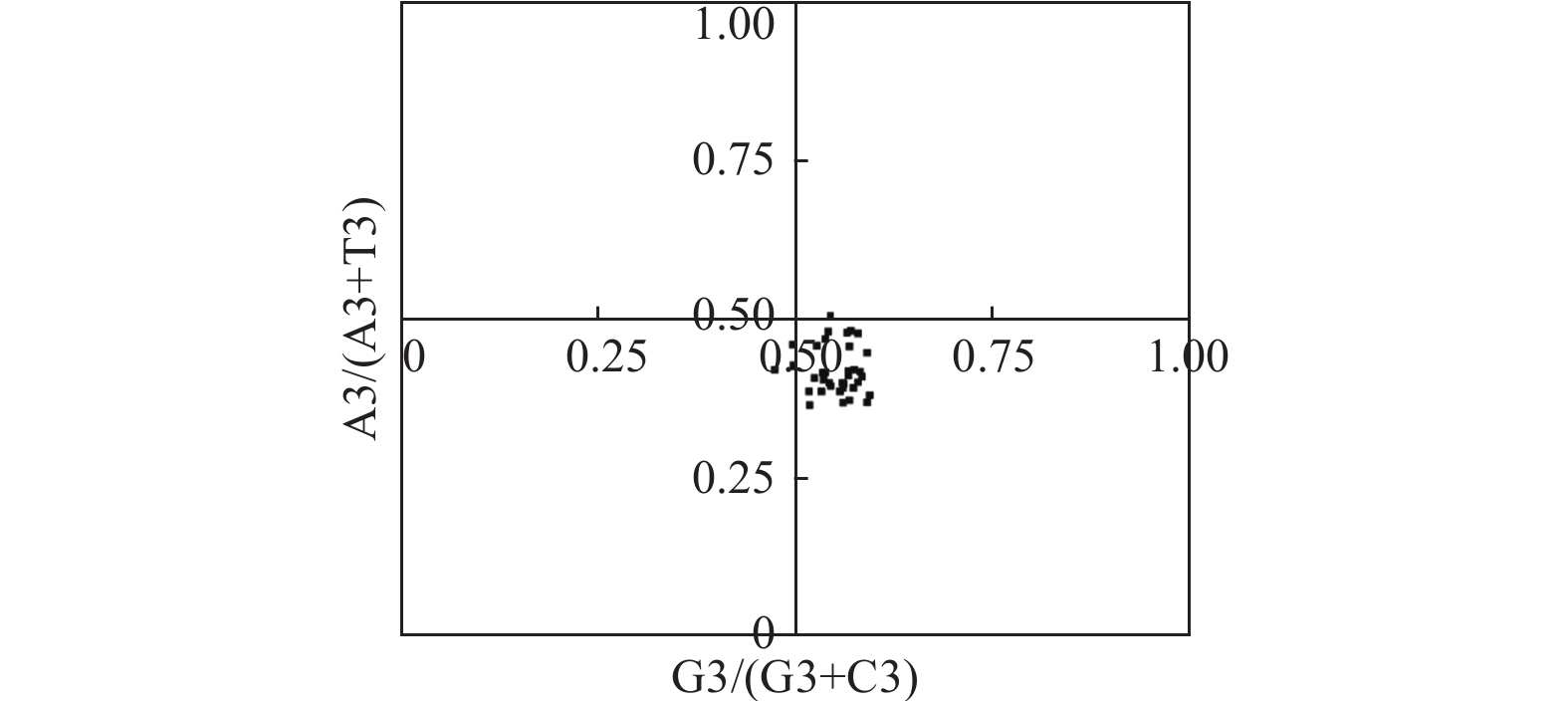
图3显示:在PR2-plot分析中,A3/(A3 + T3)或G3/(G3 + C3)值都偏离了0.5,大都分布在平面图的右下方,表明密码子第3位碱基T的频率高于A,G的频率高于C,表明多数物种DXR密码子偏好模式很大程度上受自然选择的影响[21],与中性分析的结果一致。
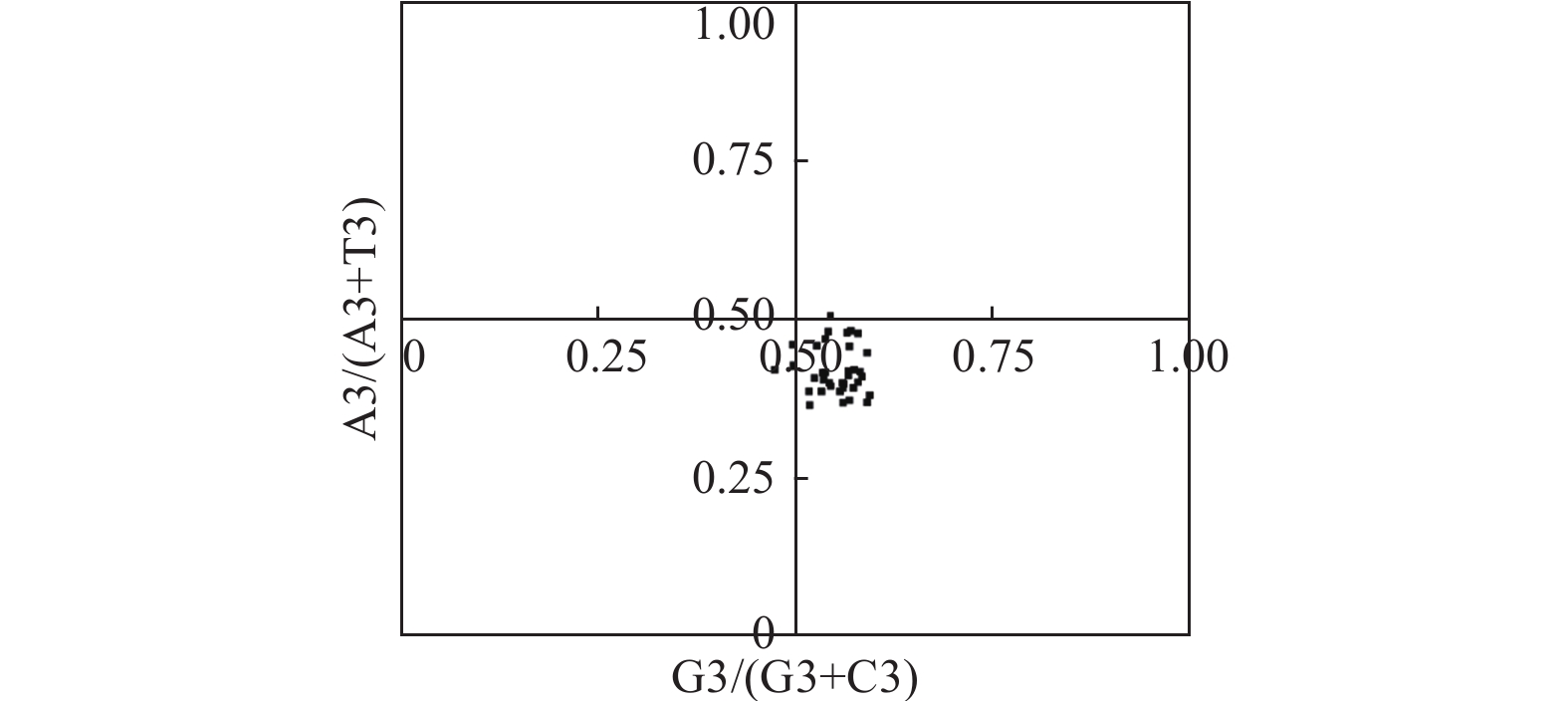
Figure 3. PR2-plot analysis of of DXR genes in different species
-
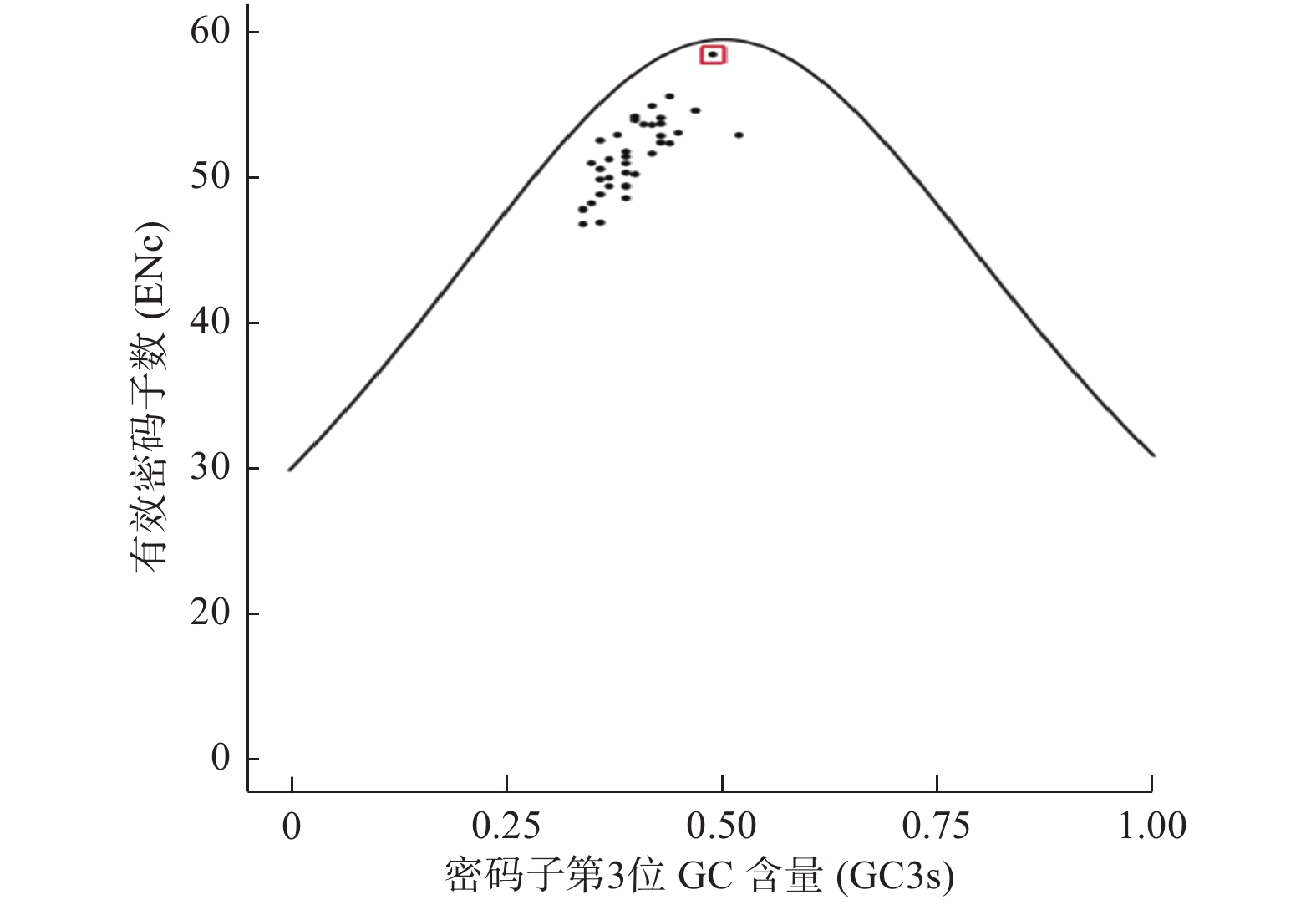
ENc-plot分析结果(图4)发现:38个物种DXR基因位点均分布在标准曲线下方,实际ENc值与理论ENc值存在差异,且大部分离标准曲线较远,说明大部分DXR基因密码子偏好性形成受自然选择的作用较大;但其中夏枯草DXR基因位点距离标准曲线较近,说明夏枯草DXR基因偏好性主要受突变影响。
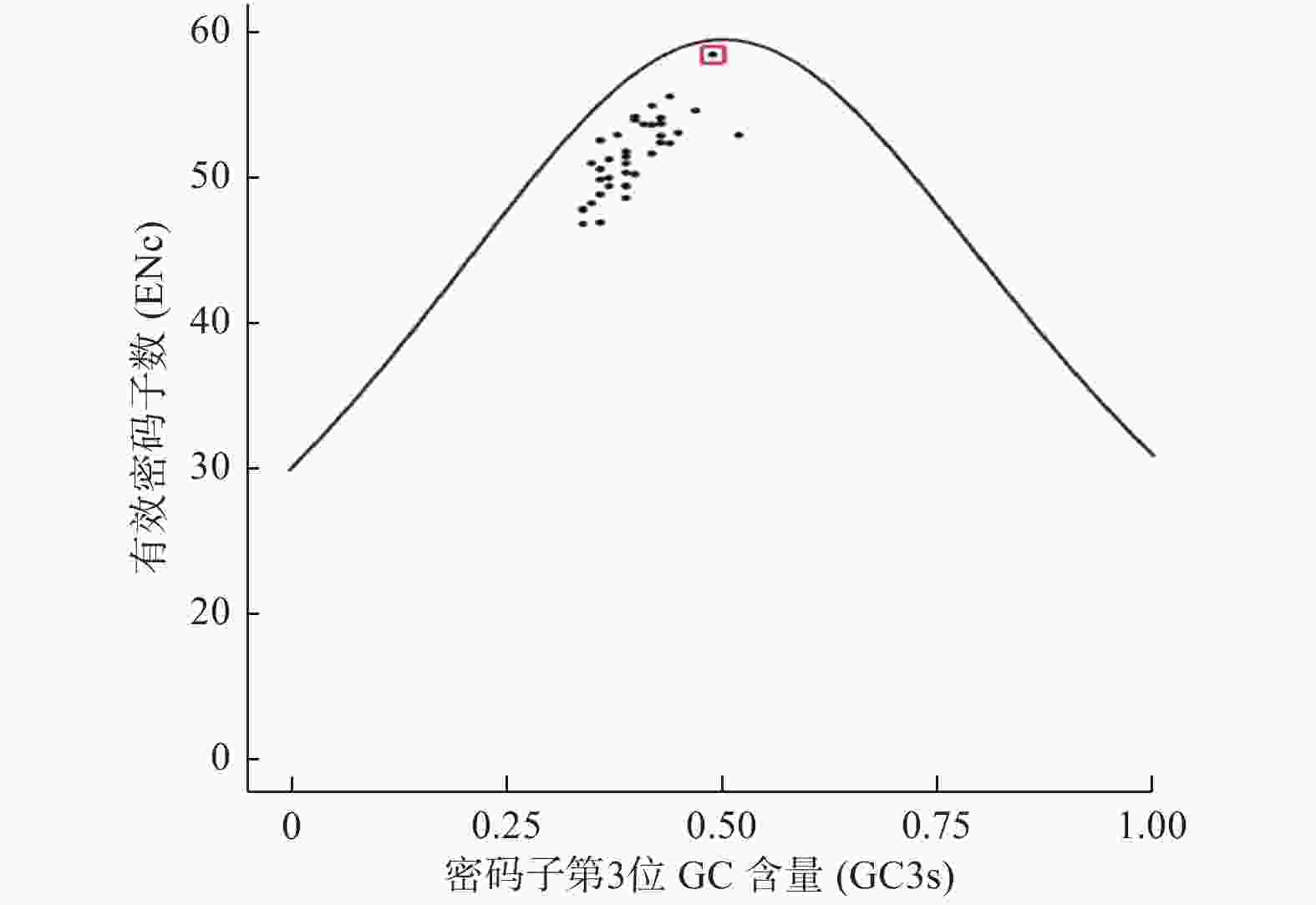
Figure 4. ENc-plot of DXR genes in different species
-
采用邻接法构建38个物种DXR基因的CDS序列系统进化树(图5)。38个物种聚为2大类,第1类由裸子植物组成,第2类由单子叶植物和双子叶植物组成。在单子叶植物中,3种禾本科植物的CDS序列同源性较高,聚在一个小的分支上。使用SPSS20.0软件对38个物种DXR基因的RSCU值进行聚类分析(图6),同样将38个物种聚为2大类,第1大类为裸子植物,其中,同属于松科的马尾松、赤松和火炬松的DXR基因聚在一个小支上,与银杏、东北红豆杉的2个小支共聚在一个大支上,表明具有相近亲缘关系的物种在密码子使用模式上较为相似。第2大类中,禾本科植物仍聚为一个小类别,同属蔷薇科的枇杷、西洋梨和玫瑰聚为一小类,菊科植物黄花蒿、艾草与属于龙胆科的滇龙胆草聚为一小类,但是也发现基于RSCU值的聚类分析结果与CDS序列构建的进化树存在差异。由此可见,并非所有亲缘关系较近的物种其DXR基因的密码子使用偏好性都具有相似性,部分物种在长期进化过程中可能会形成独特的密码子使用偏好性。
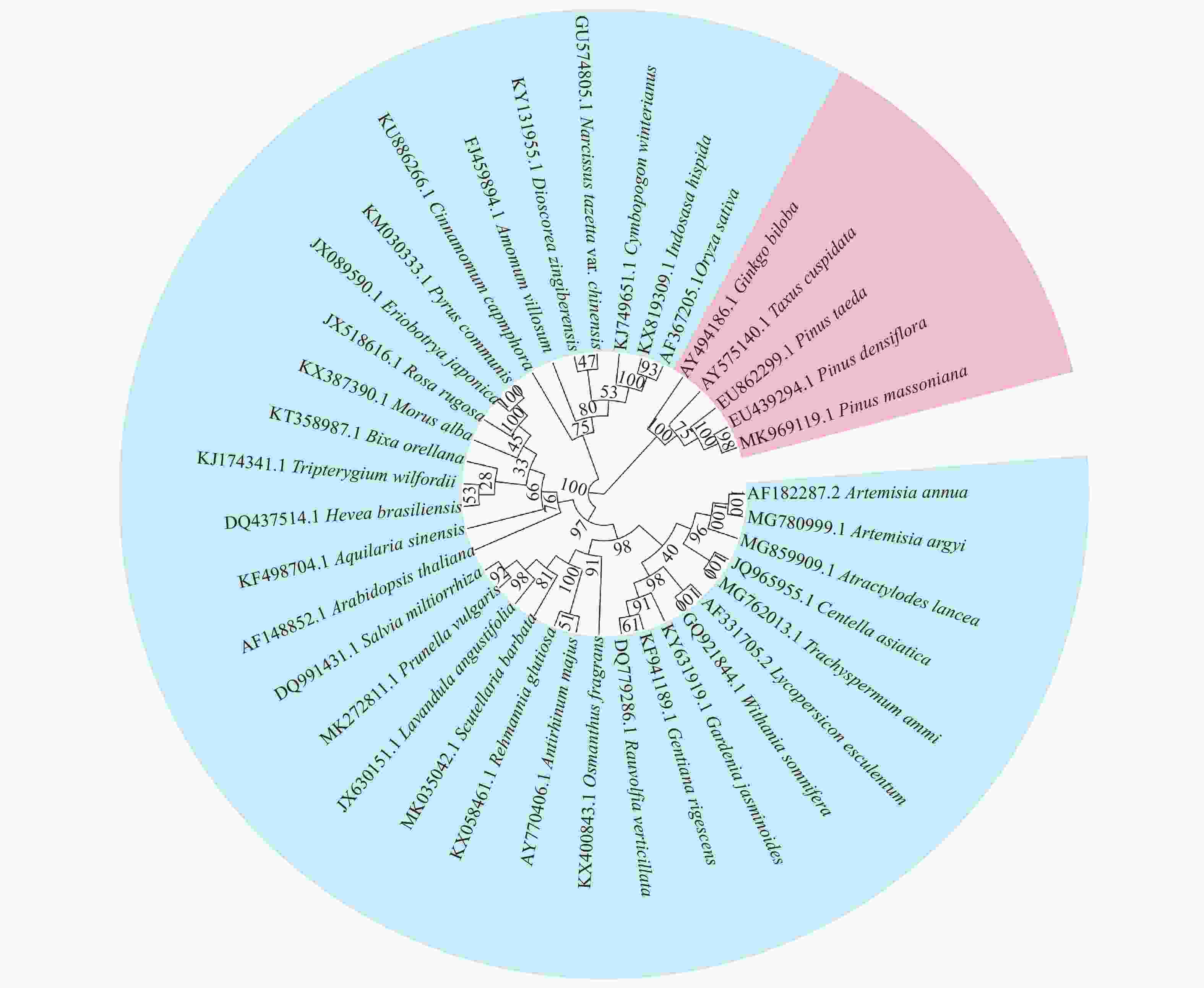
Figure 5. evolutionary tree of DXR

Figure 6. RSCU cluster analysis of DXR
-
将马尾松PmDXR密码子使用频率分别与拟南芥、烟草、酿酒酵母、大肠杆菌基因组密码子的使用频率进行比较(表4)。密码子使用频率比值是评价物种间密码子使用偏好性差异的指标之一,若比值为0.5~2.0,表示偏好性差异较小,若比值≤0.5、≥2.0,则表示偏好性差异较大[22]。在异源表达过程中,应尽量选择密码子使用偏好性差异较小的作为受体,以提高转录、翻译效率,进而提高表达量。分析结果表明:PmDXR与酿酒酵母基因组有15个密码子的偏好性差异较大,与大肠杆菌基因组密码子偏好性差异较大有25个,可见,酵母更适合做PmDXR基因的真核表达系统。PmDXR基因与拟南芥、烟草基因组分别有12、9个密码子的使用频率差异较大,说明在PmDXR基因遗传转化试验中,烟草相较拟南芥更适合作为异源表达受体。
氨基酸
Amino acid密码子
Codon马尾松DXR基因
PmDXR拟南芥基因组
At烟草基因组
Nt酵母基因组
Sc大肠杆菌基因组
EcPmDXR/At PmDXR/Nt PmDXR/Sc PmDXR/Ec Ala GCA 37.879 17.5 23.1 16.2 20.6 2.16 1.64 2.34 1.84 GCC 18.939 10.3 12.5 12.6 25.1 1.84 1.52 1.50 0.75 GCG 3.788 9.0 5.8 6.2 31.7 0.42 0.65 0.61 0.12 GCT 39.773 28.3 31.2 21.2 15.6 1.41 1.27 1.88 2.55 Arg AGA 22.727 19.0 16.0 21.3 2.9 1.20 1.42 1.07 7.84 AGG 5.682 11.0 12.2 9.2 1.9 0.52 0.47 0.62 2.99 CGA 5.682 6.3 5.3 3.0 3.9 0.90 1.07 1.89 1.46 CGC 0.000 3.8 3.9 2.6 21.0 0.00 0.00 0.00 0.00 CGG 3.788 4.9 3.7 1.7 6.3 0.77 1.02 2.23 0.60 CGT 0.000 9.0 7.5 6.4 20.3 0.00 0.00 0.00 0.00 Asn AAC 3.788 20.9 17.9 24.8 21.4 0.18 0.21 0.15 0.18 AAT 13.258 22.3 28.0 35.7 18.6 0.59 0.47 0.37 0.71 Asp GAC 18.939 17.2 16.9 20.2 18.6 1.10 1.12 0.94 1.02 GAT 30.303 36.6 36.9 37.6 32.1 0.83 0.82 0.81 0.94 Cys TGC 7.576 7.2 7.2 4.8 6.9 1.05 1.05 1.58 1.10 TGT 7.576 10.5 9.8 8.1 5.5 0.72 0.77 0.94 1.38 Gln CAA 11.364 19.4 20.7 27.3 15.0 0.59 0.55 0.42 0.76 CAG 7.576 15.2 15.0 12.1 29.5 0.50 0.51 0.63 0.26 Glu GAA 30.303 34.3 36.0 45.6 38.2 0.88 0.84 0.66 0.79 GAG 34.091 32.2 29.4 19.2 17.7 1.06 1.16 1.78 1.93 Gly GGA 26.515 24.2 23.2 10.9 9.0 1.10 1.14 2.43 2.95 GGC 9.470 9.2 11.2 9.8 27.9 1.03 0.85 0.97 0.34 GGG 13.258 10.2 10.5 6.0 11.3 1.30 1.26 2.21 1.17 GGT 17.045 22.2 22.3 23.9 24.4 0.77 0.76 0.71 0.70 His CAC 17.045 8.7 8.7 7.8 9.8 1.96 1.96 2.19 1.74 CAT 13.258 13.8 13.4 13.6 13.6 0.96 0.99 0.97 0.97 Ile ATA 30.303 12.6 14.0 17.8 5.4 2.41 2.16 1.70 5.61 ATC 11.364 18.5 13.9 17.2 24.2 0.61 0.82 0.66 0.47 ATT 35.985 21.5 27.8 30.1 29.8 1.67 1.29 1.20 1.21 Leu CTA 15.152 9.9 9.4 13.4 4.0 1.53 1.61 1.13 3.79 CTC 9.470 16.1 12.3 5.4 11.0 0.59 0.77 1.75 0.86 CTG 13.258 9.8 10.2 10.5 50.9 1.35 1.30 1.26 0.26 CTT 22.727 24.1 24.0 12.3 11.7 0.94 0.95 1.85 1.94 TTA 17.045 12.7 13.4 26.2 13.9 1.34 1.27 0.65 1.23 TTG 18.939 20.9 22.3 27.2 14.0 0.91 0.85 0.70 1.35 Lys AAA 35.985 30.8 32.6 41.9 33.2 1.17 1.10 0.86 1.08 AAG 30.303 32.7 33.5 30.8 10.7 0.93 0.90 0.98 2.83 Met ATG 18.939 24.5 25.0 20.9 27.0 0.77 0.76 0.91 0.70 Phe TTC 11.364 20.7 18.0 18.4 16.9 0.55 0.63 0.62 0.67 TTT 24.621 21.8 25.1 26.1 23.2 1.13 0.98 0.94 1.06 Pro CCA 28.409 16.1 19.8 18.3 8.5 1.76 1.43 1.55 3.34 CCC 5.682 5.3 6.6 6.8 5.8 1.07 0.86 0.84 0.98 CCG 3.788 8.6 5.0 5.3 21.8 0.44 0.76 0.71 0.17 CCT 26.515 18.7 18.7 13.5 7.3 1.42 1.42 1.96 3.63 Ser AGC 3.788 11.3 10.0 9.8 16.0 0.34 0.38 0.39 0.24 AGT 5.682 14.0 13.3 14.2 9.5 0.41 0.43 0.40 0.60 TCA 18.939 18.3 17.6 18.7 7.8 1.03 1.08 1.01 2.43 TCC 11.364 11.2 10.2 14.2 8.9 1.01 1.11 0.80 1.28 TCG 3.788 9.3 5.3 8.6 8.7 0.41 0.71 0.44 0.44 TCT 24.621 25.2 20.0 23.5 8.7 0.98 1.23 1.05 2.83 Thr ACA 24.621 15.7 17.4 17.8 8.2 1.57 1.42 1.38 3.00 ACC 13.258 10.3 9.7 12.7 22.8 1.29 1.37 1.04 0.58 ACG 1.894 7.7 4.5 8.0 14.8 0.25 0.42 0.24 0.13 ACT 9.470 17.5 20.3 20.3 9.1 0.54 0.47 0.47 1.04 Trp TGG 15.152 12.5 12.2 10.4 15.2 1.21 1.24 1.46 1.00 Tyr TAC 1.894 13.7 13.5 14.8 12.1 0.14 0.14 0.13 0.16 TAT 18.939 14.6 17.8 18.8 16.5 1.30 1.06 1.01 1.15 Val GTA 20.833 9.9 11.4 11.8 11.1 2.10 1.83 1.77 1.88 GTC 7.576 12.8 11.1 11.8 15.1 0.59 0.68 0.64 0.50 GTG 18.939 17.4 16.7 10.8 25.5 1.09 1.13 1.75 0.74 GTT 37.879 27.2 26.8 22.1 18.5 1.39 1.41 1.71 2.05 * TAA 0.000 0.9 1.1 1.1 2.0 0.00 0.00 0.00 0.00 * TAG 0.000 0.5 0.5 0.5 0.3 0.00 0.00 0.00 0.00 * TGA 1.894 1.2 1.0 0.7 1.1 1.58 1.89 2.71 1.72 注:下划线表示密码子使用频率差异较大,*表示终止密码子。
Notes: The underline indicates that the codon usage frequency is quite different, * indicates the stop codon.Table 4. Comparison of codon usage preference between PmDXR gene in P. massoniana and other model organisms
2.1. 马尾松DXR基因密码子偏性分析
2.1.1. GC、GC3s、ENc和CAI分析
2.1.2. 同义密码子相对使用度分析
2.2. 38个物种DXR基因密码子使用偏好性比较
2.2.1. 相关参数比较分析
2.2.2. DXR同义密码子相对使用度分析
2.2.3. DXR密码子成分相关分析
2.2.4. 不同物种DXR基因中性绘图分析
2.2.5. 不同物种DXR基因PR2-plot分析
2.2.6. 不同物种DXR基因ENc-plot分析
2.3. DXR基因CDS序列和密码子使用偏好性的系统聚类分析


2.4. 马尾松DXR基因的外源宿主选择
-
在漫长的自然选择和进化进程中,各物种形成了特定的密码子使用模式,其使用偏好性对研究基因进化和功能鉴定具有重要的生物学意义。本研究用生物信息学方法对马尾松PmDXR基因密码子偏好模式进行系统分析,确定了该基因有12个高频率密码子,其中,AGA、TCT具有极强的偏好性;同时发现编码Arg的CGC、CGT可能不参与PmDXR基因的编码过程。PmDXR基因中密码子的GC和GC3s含量分别为43.75%和34.47%,说明其密码子比较偏好以A/T结尾,这一结果符合马尾松总体上偏好使用第3位为A/T碱基的密码子的特征[23]。38个物种DXR基因的CAI值介于0.184~0.244,ENc值为47.89~59.49,平均值为52.59,其中,裸子植物DXR基因的ENc平均值为49.04,较单子叶植物和双子叶植物低,表明裸子植物DXR基因的密码子偏好性相对双子叶和单子叶植物较高。由此可见,DXR基因密码子偏好性较弱,在各物种的表达水平普遍不高,但物种间还是存在一定差异。
密码子偏好性的形成受突变压力和自然选择等因素的综合影响,对不同物种DXR基因的中性分析、ENc-plot分析、PR2分析结果均表明,DXR基因密码子偏好性形成的主要影响因素为选择压力。基于RSCU值的聚类分析与基于CDS的进化分析结果表明,具有相近亲缘关系的物种其密码子使用偏好性较为相似,但部分物种基于RSCU值的聚类分析与基于CDS序列的进化分析结果存在差异,推测部分物种在长期的进化过程中可能形成了独特的密码子使用模式,这在前人的研究结果中也曾出现[24-28]。本研究中,松科植物在2种聚类结果中均聚在一个大支上,3种禾本科植物基于CDS序列聚类分析与基于RSCU值的聚类结果也相似,双子叶植物中同属一个科植物的种聚类分析结果具有一定相似性,这表明物种间亲缘关系越近,其DXR基因的密码子使用差异越小。
在异源表达过程中,为实现外源基因的成功表达并提高其表达量,应尽量选择密码子使用偏好性差异较小的作为受体。DXR作为萜类合成途径中的关键酶基因,了解其密码子使用特性并选择合适的异源表达受体对研究其功能具有重要指导作用。本研究通过比较PmDXR基因与拟南芥、烟草、酿酒酵母、大肠杆菌的基因组密码子使用频率的差异,发现相比于大肠杆菌原核表达系统,酵母真核表达系统更适合作为PmDXR基因的表达系统。在PmDXR遗传转化功能验证中,模式植物拟南芥、烟草均可作为其遗传转化受体,其中,烟草更为理想。本研究结果初步揭示了PmDXR基因密码子的使用规律,可为后续开展基因功能验证等相关研究提供理论基础。目前,马尾松还没有成熟的遗传转化体系,对PmDXR基因的研究通常需要借助遗传转化体系成熟的受体,今后可依据此研究结果为PmDXR基因选择合适的遗传转化受体,进一步了解其在MEP途径中的调控机制。
-
本研究表明:马尾松PmDXR基因的密码子偏好性较弱,偏好使用A/T结尾的密码子。不同物种DXR基因的密码子偏好性存在一定差异,整体看,裸子植物DXR基因密码子偏好性比被子植物强。基于RSCU值和基于CDS的聚类分析结果存在一定差异,且基于CDS序列的进化分析结果更能反映物种亲缘关系的远近。中性分析、PR2-plot分析、ENc-plot分析结果均表明DXR基因密码子偏好性形成主要受选择压力的影响。酵母真核表达系统较大肠杆菌原核表达系统更适合马尾松PmDXR异源表达,模式植物烟草较拟南芥更适合做马尾松PmDXR遗传转化受体。
 DownLoad:
DownLoad: