


-
沙棘(Hippophae rhamnoides L.)是胡颓子科沙棘属的一种雌雄异株的灌木或小乔木,耐旱性和耐寒性良好。沙棘果实具有独特的药用和营养价值,在中国和俄罗斯有上百年的药用历史[1]。沙棘果实富含维生素、有机酸、氨基酸、脂肪酸、抗氧化剂和类黄酮[2-4],在炎症性疾病、肝脏性疾病、动脉粥样硬化等疾病的治疗以及保护心脏等方面具有显著功效[5]。在沙棘种子含有的众多脂肪酸中,α-亚麻酸(α-Linolenic acid,C18:3N3)的含量十分丰富,且α-亚麻酸对大脑发育、心血管健康、炎症治疗等都具有重要作用[6-9],但α-亚麻酸是人体必需脂肪酸,不能在人体内合成,只能通过食物摄取。豆油、花生油等常见植物食用油中的主要脂肪酸为油酸、亚油酸、棕榈酸和硬脂酸[10-11],α-亚麻酸含量极低,而沙棘中高积累20.3%~36.3%的α-亚麻酸[12],并且沙棘作为耐旱木本植物可以在沙地、荒山、盐碱地等边际土地生长,可避免与大田油料作物争夺土地,是理想的α-亚麻酸供给对象。
目前,油料作物脂肪酸合成途径及调控基因的研究较多,在沙棘的相关研究中多围绕脂肪酸组分及脂肪酸测定方法展开,少有针对沙棘中α-亚麻酸合成机制的研究。本研究选取α-亚麻酸含量存在差异的2个沙棘品种种子,对其含量差异及相关基因进行分析,在基因表达水平上研究沙棘中α-亚麻酸合成及代谢规律,为培育高α-亚麻酸含量的沙棘良种奠定理论基础。
-
沙棘种子采自中国林业科学研究院沙漠林业实验中心(内蒙古磴口县),2个沙棘品种——蒙古大果沙棘向阳(Hippophae rhamnoides ‘Mongolia’)(XY)和中国沙棘丰宁(H. rhamnoides ‘Sinensis’)(FN)的种子分别采自3个不同的发育时期,依次为未成熟期(T1)、半成熟期(T2)和成熟期(T3),在这3个时期沙棘种子的种皮颜色依次呈现为白色、黑白相间斑点色、黑色。因沙棘种皮与果皮的转色期基本一致,为保证时期界定更加精准,同时利用色差计测量沙棘果皮的红绿色指标 a 值和黄蓝指标 b 值,计算色泽比(h=a/b),种皮呈白色且果皮h值介于−0.18~−0.10的界定为T1时期,种皮呈黑白相间斑点色且果皮h值介于0.20~0.28的界定为T2时期,种皮呈黑色且果皮h值介于0.52~0.60的界定为T3时期。沙棘果实采集后快速将种子从果实中剥离,放入液氮中进行速冻,然后转移至−80℃冰箱长久保存,用于下一步分析。
-
将样品冰上解冻,取50 mg样品于2 mL离心管中。加入1 mL的氯仿甲醇(2:1 v/v)溶液,超声30 min。取上清液,加入1%硫酸甲醇溶液2 mL,80℃水浴,甲酯化30 min,随后加入1 mL的正己烷萃取,5 mL的纯水洗涤。吸取上清液500 μL,加入25 μL的水杨酸甲酯(500 mg·L−1)作为内标,混匀加入进样瓶,进样量1 μL,分流比 10∶1,分流进样,进行GC-MS 检测。
-
样品采用 AgilentDB-WAX 毛细管柱(30 m×0.25 mmID×0.25 μm)气相色谱系统进行分离。程序升温:初始温度为 50℃,保持 3 min,随后以10℃·min−1的速度升温至220℃并维持 20 min。载气为氦气,载气流速1.0 mL·min−1。样本队列中每间隔6个实验样本设置1个QC样本,用于检测和评价系统的稳定性及重复性。采用 Agilent7890A/5975C 气-质联用仪进行质谱分析。进样口温度 280℃,离子源温度230℃,传输线温度250℃。电子轰击电离(EI)源,SIM 扫描方式,电子能量70 eV。采用 MSDChemStation 软件提取色谱峰面积及保留时间。绘制标准曲线,计算样品中中长链脂肪酸的含量。
-
按照Qingen(Qingen,DE)试剂盒[13]操作说明对样品中的总RNA进行提取,并去除总RNA中DNA片段的残留。分别使用NanoPhotometer 分光光度仪、Qbuit 2.0 荧光计和Qubit RNA 检测试剂盒以及Agilent 2100和RNA Nano 6000试剂盒检测RNA纯度、RNA浓度以及RNA完整性。RNA检测合格后进行文库制备。使用Illumina HiSeq 2000 对RNA文库进行测序,获得原始测序数据。为了保证测序数据质量,对原始数据进行过滤,从而获得高质量数据(Clean data),用于后续生物信息分析。首先使用DESeq R(1.18.0)[14]进行基因表达量分析,筛选差异表达基因,然后通过GOseq R数据包[15]和KOBAS(2.0)[16]分别对差异表达基因进行GO功能注释和KEGG通路分析。
-
利用气相色谱分析,在2个品种沙棘的种子中共检测到27种脂肪酸(表1),其中,主要脂肪酸为C18:3N3(α-亚麻酸)、C18:2(亚油酸)、C16:0(棕榈酸)、C18:1(油酸)、C18:0(硬脂酸)。随着果实不断成熟,2个品种沙棘种子中,5种主要脂肪酸的含量均逐渐增加,其中,α-亚麻酸和亚油酸含量上升幅度较大。
表 1 XY与FN 2个沙棘品种种子在不同发育时期脂肪酸含量的动态变化
Table 1. Dynamic changes of fatty acid composition in the seeds of XY and FN at different developmental stages
μg·g−1 组分
CompositionsXY FN T1 T2 T3 T1 T2 T3 C10:0 1.04±0.09 b 1.75±0.13 ab 2.43±0.69 a 1.04±0.07 b 1.72±0.20 ab 2.24±0.20 a C12:0 1.11±0.27 a 2.06±0.78 a 5.62±0.32 a 0.34±0.40 a 1.34±0.17 a 2.09±1.25 a C13:0 0.08±0.05 b 0.30±0.08 ab 0.35±0.188 a 0.08±0.02 b 0.31±0.10 ab 0.48±0.10 a C14:0 20.70±2.32 c 40.23±2.638 a 53.98±18.17 a 17.66±1.34 c 40.85±7.66 a 58.91±7.54 a C14:1 136.34±14.20 a 145.86±15.94 a 122.48±14.74 a 121.50±9.22 a 121.08±9.35 a 117.06±7.36 a C15:0 6.20±1.43 c 19.42±1.16 b 28.45±10.91 ab 6.84±0.91 c 24.40±5.20 b 37.00±5.97 a C15:1 16.90±2.09 a 14.37±2.21 ab 8.59±0.85 c 14.65±0.78 ab 12.06±4.19 abc 10.52±1.83 bc C16:0 1 219.70±135.84 b 2 337.41±132.23 a 2 873.02±915.18 a 1 141.08±69.79 b 2 482.63±282.95 a 3 165.25±346.54 a C16:1 38.04±6.73 c 70.55±12.28 bc 112.18±36.82 ab 28.40±4.84 c 86.75±22.13 ab 126.20±25.12 a C17:0 7.89±1.39 b 16.93±1.34 a 19.21±6.58 a 9.02±0.85 b 15.92±1.62 a 21.51±2.86 a C17:1 66.01±7.57 a 51.99±6.67 ab 47.64±2.13 ab 56.48±12.46 ab 44.60±3.13 b 51.85±7.10 ab C18:0 695.59±62.14 c 1 330.90±97.82 ab 1 656.18±553.21 a 634.31±23.71 c 1 035.51±73.38 bc 1 280.61±137.68 ab C18:1 78.99±20.21 d 1 824.10±207.78 c 3 447.79±1 190.81 a 101.66±20.47 d 2 197.36±133.31 bc 3 029.64±464.58 ab C18:2 311.23±62.81 d 4 231.52±246.51 c 6 879.55±2 327.19 ab 265.09±48.17 d 5 028.36±863.33 bc 7 443.04±795.32 a C18:3N3 298.54±76.45 c 4 980.34±261.85 ab 7 463.53±2 636.16 a 223.36±46.56 c 4 307.42±690.29 b 6 526.07±969.49 ab C20:0 19.50±3.83 c 102.61±13.30 b 200.82±74.68 a 16.14±2.06 c 94.96±7.90 b 139.33±19.39 b C20:1 7.42±1.60 c 50.74±4.18 b 90.17±30.25 a 5.87±0.58 c 53.50±4.71 b 83.41±16.89 a C20:2 0.93±0.22 c 8.69±0.52 b 14.62±5.51 ab 0.81±0.13 c 9.72±3.29 b 16.89±3.08 a C21:0 3.85±0.72 b 8.44±0.77 ab 11.93±3.97 a 3.76±0.51 b 8.15±1.48 ab 11.94±2.08 a C20:3N3 1.90±0.21 bc 3.02±0.23 ab 3.86±0.93 a 1.53±0.28 c 2.51±0.55 bc 3.94±0.72 a C20:5N3 0 a 0.15±0.08 a 1.04±0.79 a 0 a 0.33±0.03 a 0.87±0.61 a C22:0 12.20±1.62 c 36.59±4.23 b 57.26±20.45 ab 14.29±2.13 c 44.80±9.54 b 68.61±10.83 a C22:1N9 24.60±3.00 ab 29.29±6.16 a 26.30±5.35 ab 16.34±0.58 b 21.45±3.73 ab 20.85±4.84 ab C23:0 2.00±0.44 c 10.73±1.28 b 18.25±7.01 a 2.41±0.44 c 9.76±2.18 b 15.84±3.27 ab C24:0 4.74±0.60 c 17.42±2.00 b 29.66±10.75 a 5.35±0.46 c 18.09±3.08 b 26.69±4.54 ab C24:1 4.05±3.29 a 11.48±0.91 a 6.68±3.95 a 4.38±3.87 a 3.94±3.01 a 4.47±1.55 a C22:6N3 2.62±0.11 c 3.43±0.05 a 3.25±0.25 ab 2.55±0.08 c 3.07±0.09 b 3.26±0.18.00 ab 注:同行不同字母代表显著差异(P< 0.05)。
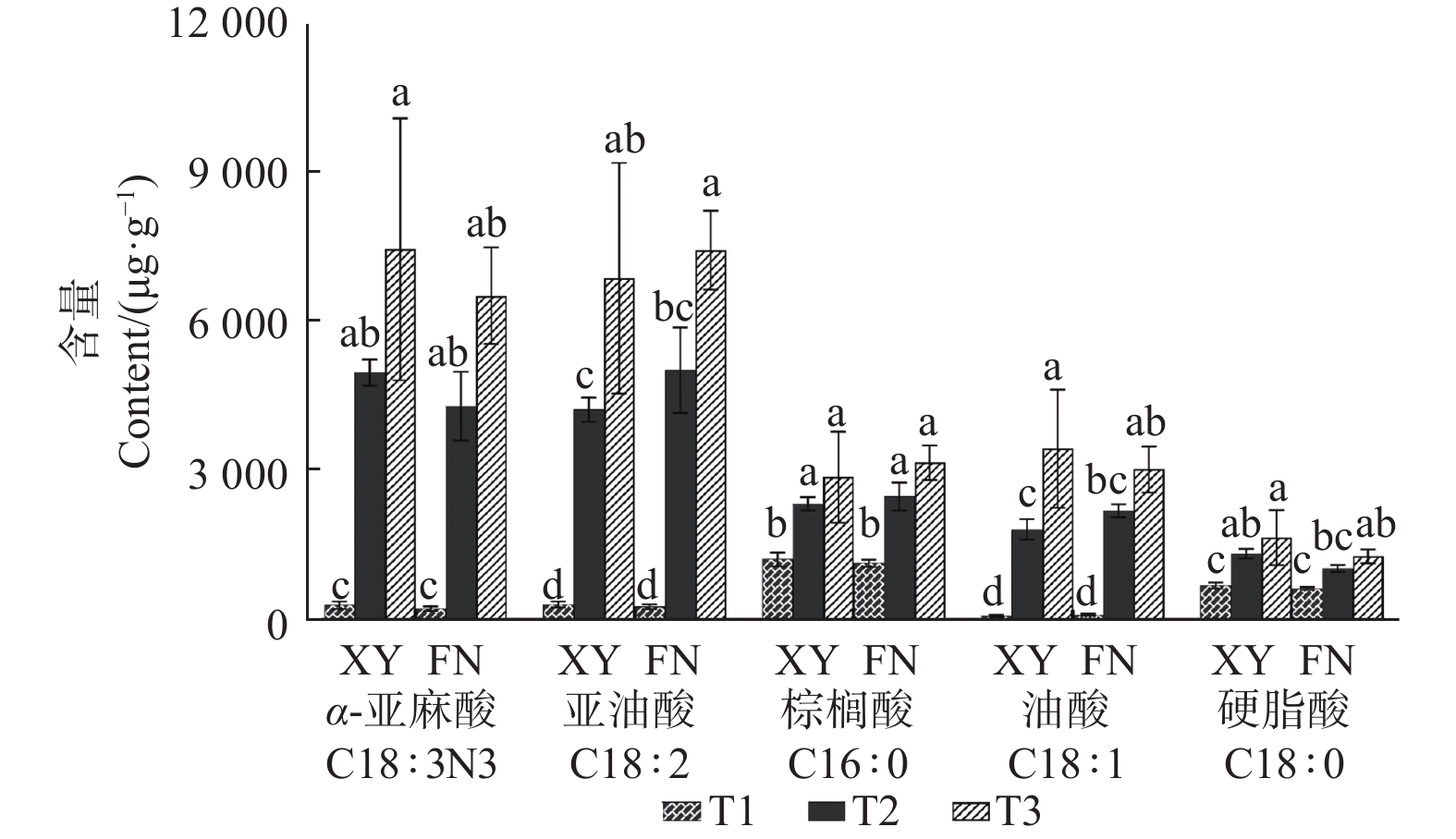
Note: Different letters in the same line represent significant differences(P< 0.05)从图1可以看出:XY与FN的5种主要脂肪酸的含量均在T2时期表现出显著差异。按照脂肪酸含量从大到小排序,在XY中,依次为α-亚麻酸、亚油酸、油酸、棕榈酸、硬脂酸,含量最高的脂肪酸为α-亚麻酸,亚油酸次之;而在FN中,脂肪酸从大到小排序依次为亚油酸、α-亚麻酸、棕榈酸、油酸、硬脂酸,亚油酸大于α-亚麻酸含量(图1)。
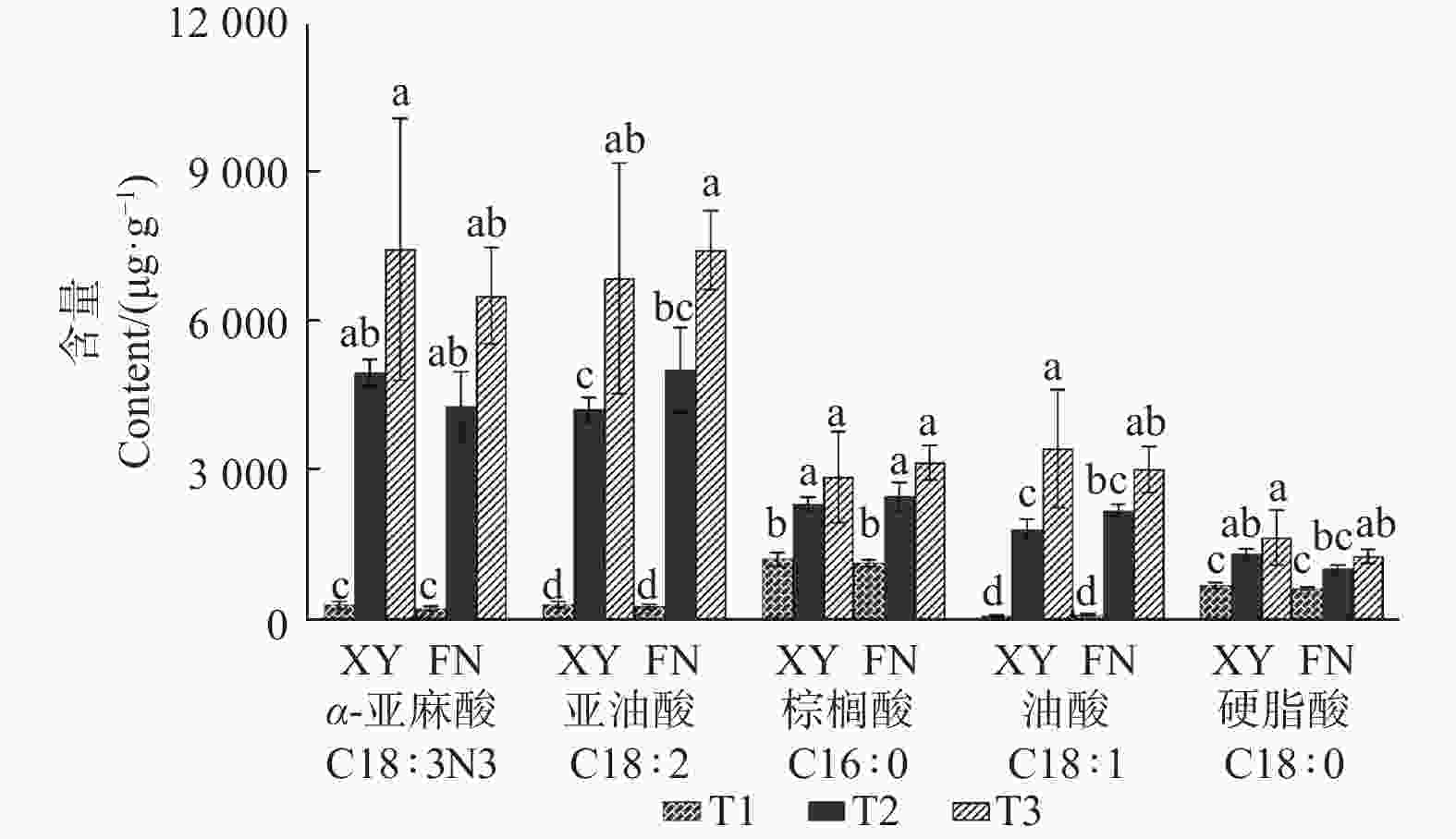
图 1 沙棘XY与FN种子中主要脂肪酸组分的含量变化
Figure 1. Distribution of main fatty acid components in XY and FN seeds of sea buckthorn
在T2时期,2个沙棘品种种子中α-亚麻酸与亚油酸的比值(α-亚麻酸/亚油酸)存在显著差异(P<0.05)。XY中,T2时期3个样本的α-亚麻酸/亚油酸比值分别为1.21、1.17、1.16,而FN分别为0.92、0.75、0.92(表2)。
表 2 T2时期XY与FN种子中的C18:3N3/C18:2
Table 2. C18:3n3 / C18:2 in XY and FN seeds of sea buckthorn at T2
品种
CultivarC18:3N3/C18:2 T2 T2-1 T2-2 T2-3 XY 1.182a 1.21 1.17 1.16 FN 0.862b 0.92 0.75 0.92 注:不同字母代表显著差异(P<0.05),T2-1、T2-2、T2-3代表T2时期的3个生物学重复样本。
Notes: Different letters in the same column represent significant differences(P<0.05).T2-1、T2-2 and T2-3represent three biological replicates of the T2 period. -
以XY与FN种子的3个发育时期、每个发育时期3个生物学重复的18个样品为材料,通过RNA-seq测序分析共产生140 G clean data,933 483 428条高质量序列(Clean reads)(Q20>97%;Q30>93%),其中,81.5%的序列唯一比对到参考基因组上。对2个品种3个不同发育时期及相同时期不同品种间的表达基因进行差异分析,共鉴定出24 917个差异表达基因(DEGs)。从图2A样本DEGs的主成分分析(PCA)可以看出:2个沙棘品种各个时期生物学重复聚类效果良好,不同时期之间与不同品种之间数据离散效果良好。通过DEGs的分布图(图2B)可以看出:在T1-T3的发育过程中,在T2时期XY与FN之间的DEGs最多。

图 2 主成分分析与差异表达基因上调和下调分布情况
Figure 2. PCA and up-regulated and down-regulated distribution of DEGs
依据序列同源性将24 917个差异表达基因进行GO功能注释,2 605个基因注释为生物过程(Biological process),554个基因注释为细胞组成(Cellular component),1 354个基因注释为分子功能(Molecular function)。为了进一步了解基因的生物学功能和相互作用,使用KEGG数据库对基因进行通路富集分析。24 917个DEGs注释到122个KEGG通路,其中,有94个DEGs的功能富集于脂肪酸合成(Fatty acid biosynthesis)通路、脂肪酸代谢(Fatty acid metabolism)通路、α-亚麻酸代谢(α-Linolenicacid metabolism)通路和亚油酸代谢(Linoleicacid metabolism)通路。
-
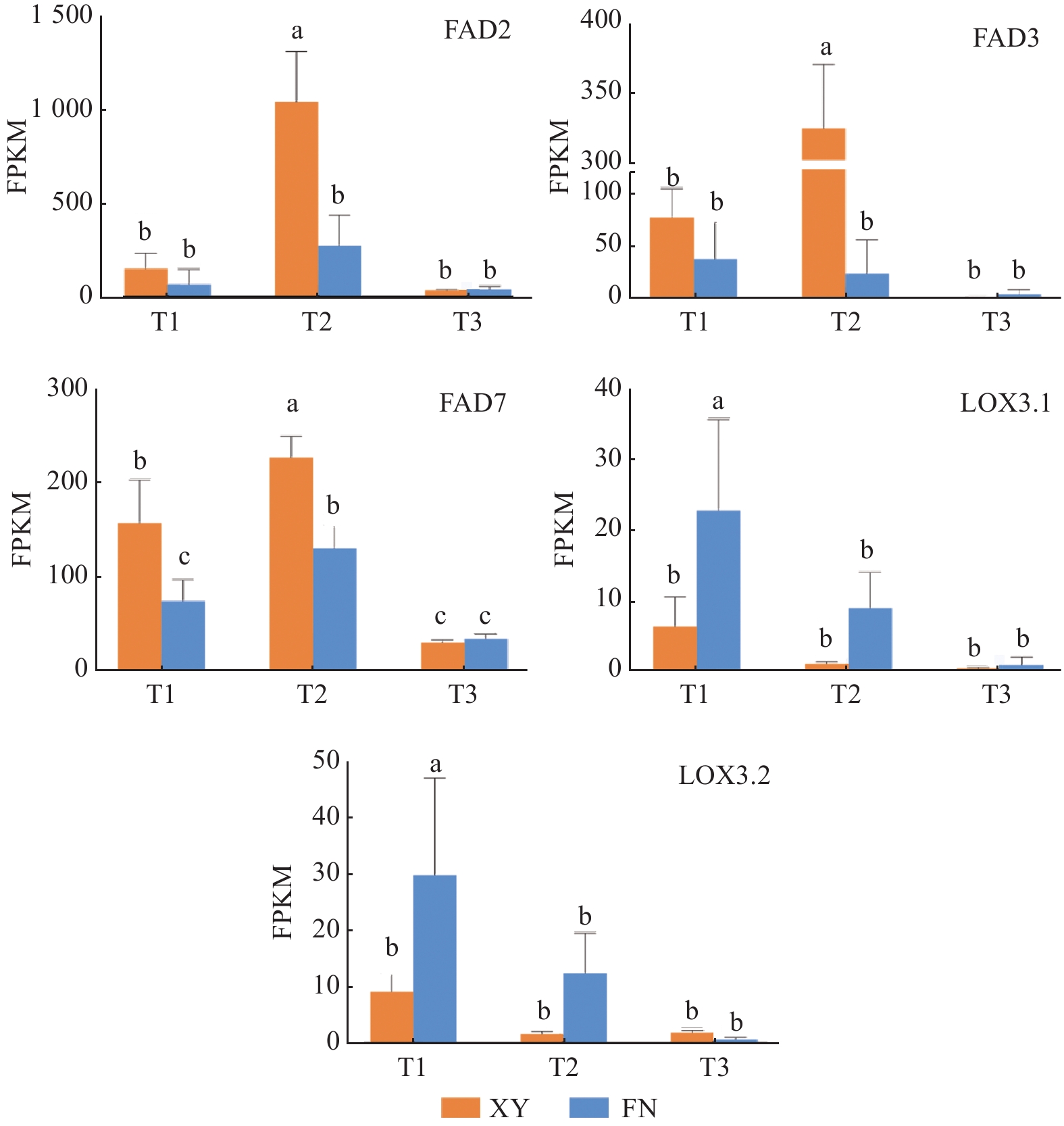
T2时期,基因FAD2在2个沙棘品种中的表达量差异显著(图3),在XY中的表达量显著高于FN,是FN的3.83倍,为α-亚麻酸(C18:3N3)的合成提供充足底物。FAD2基因在XY和FN中表达量均为先上升后下降,在第2时期表达量急剧上升达到峰值,与XY和FN中的亚油酸(C18:2)含量在第二时期显著增加(图1)相一致。

图 3 沙棘种子发育过程中与α-亚麻酸合成相关的基因表达模式
Figure 3. Gene expression profiles of Alpha linolenic acid biosynthetic genes during seed development of sea buckthorn
基因FAD3和基因FAD7调控亚麻酸生成α-亚麻酸(图4),在FN中的整体表达水平远低于XY,与XY中α-亚麻酸含量高于FN(图1)的结果相一致。基因FAD3在FN种子中的表达呈下调趋势,在XY种子中的表达水平先急剧上升,在T2时期达到峰值,后在T3时期表达量下调;T1时期,基因FAD3在XY中的表达量是FN 中的2.02倍,T2时期基因FAD3在XY和FN中的表达差异显著,在XY中的表达量是FN中的13.63倍(图3)。基因FAD7在XY和FN中的表达趋势均为先上升后下降,T1和T2时期在XY中的表达量显著高于在FN中的表达量,T1时期在XY中的表达量是FN 中的2.09倍,T2时期在XY中的表达量是FN 中的1.72倍(图3)。
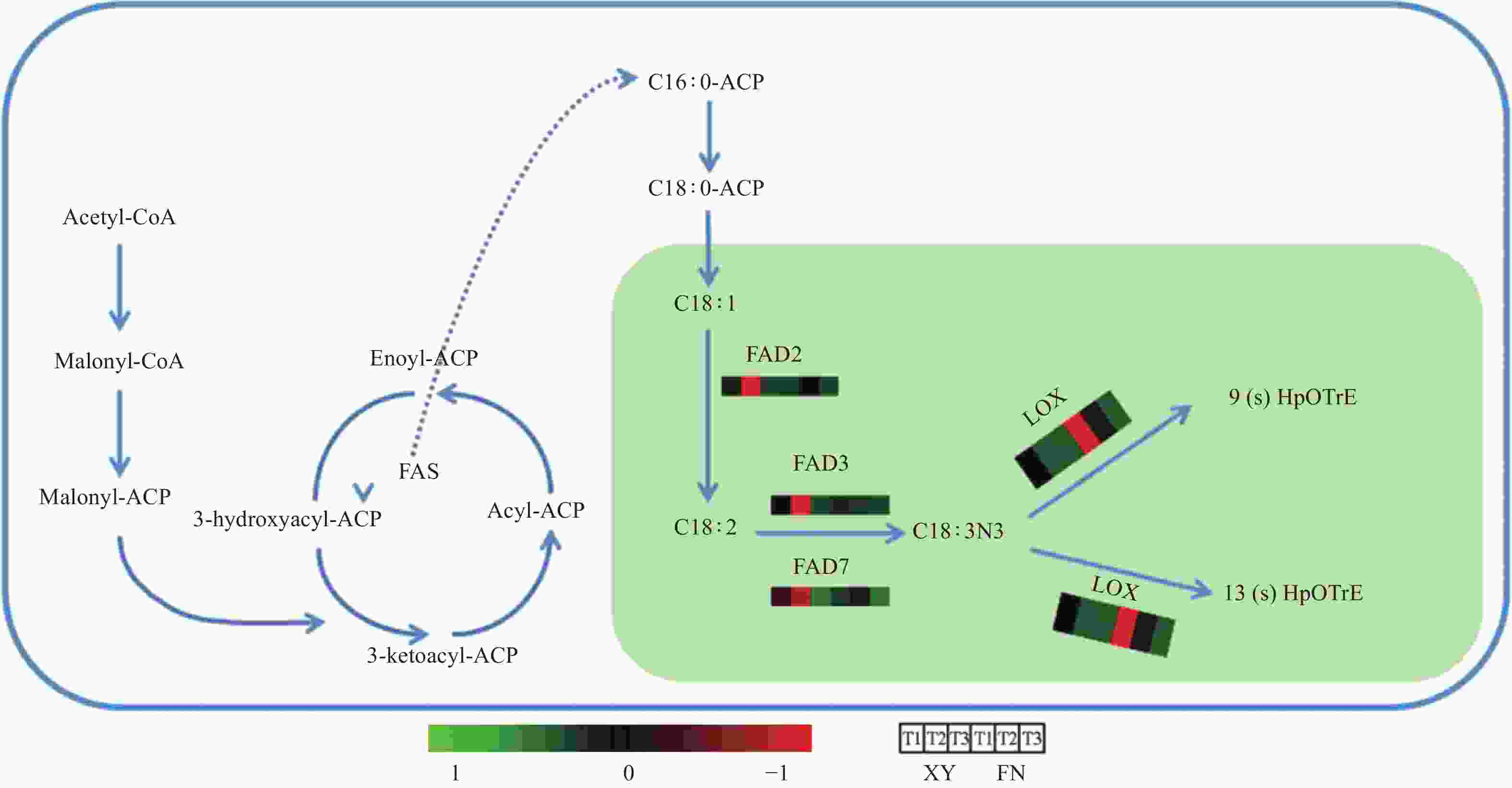
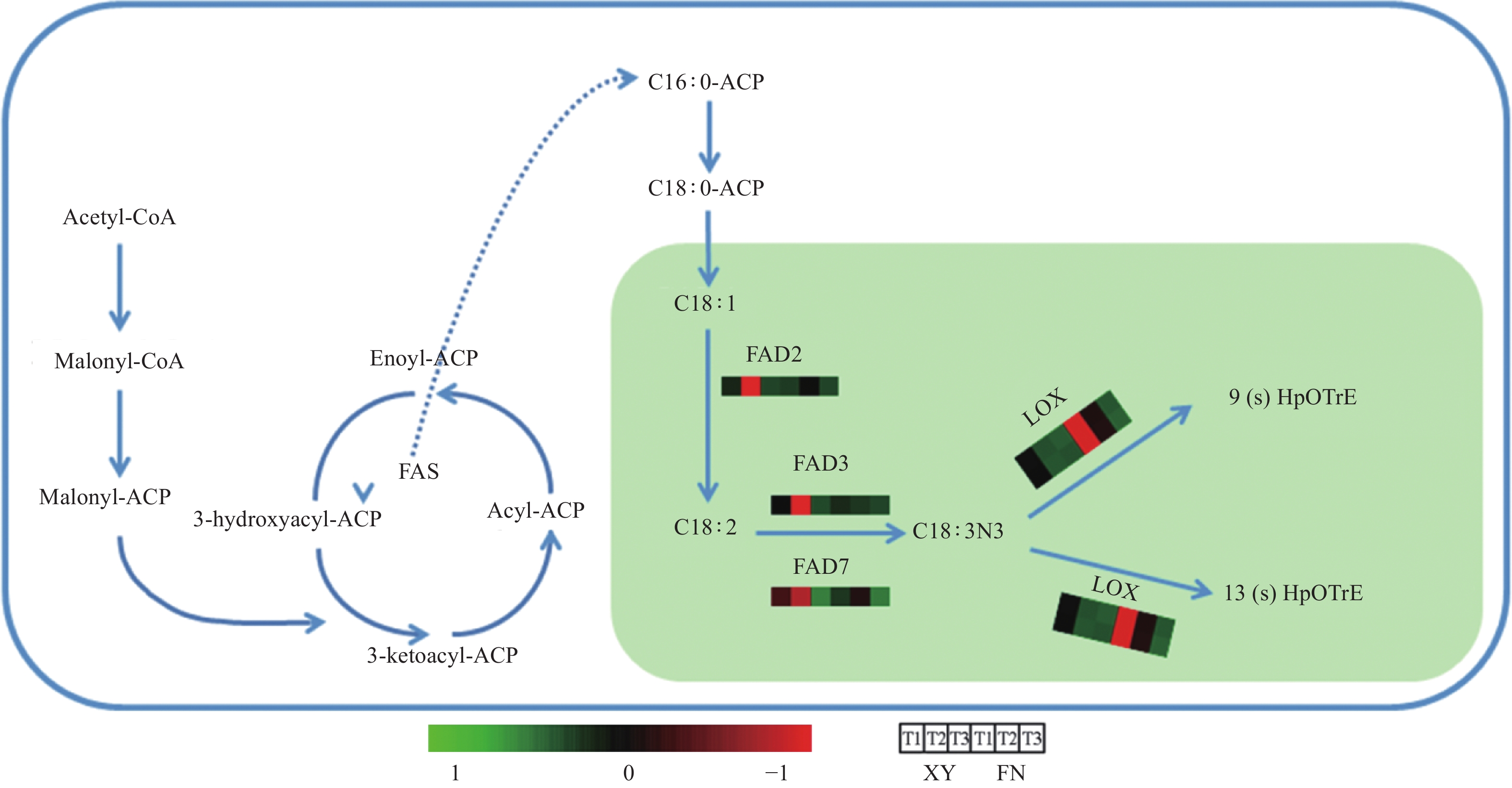
图 4 与α-亚麻酸生物合成与代谢相关的途径
Figure 4. Pathway related to the biosynthesis and metabolism of α-linolenic acid
基因LOX调控α-亚麻酸氧化(图4),生成脂肪酸氢过氧化物,是调控α-亚麻酸转化为其他物质的第一步,基因LOX3.1在XY和FN种子的生长发育过程中表达趋势均为逐渐下降,在FN中的表达量高于XY,T1时期基因LOX3.1在FN中的表达量是XY中的3.58倍,T2时期基因LOX3.1在FN中的表达量是XY中的8.02倍(图3)。基因LOX3.2在FN中的表达量同样高于XY,T1时期在FN中的表达量是XY中的3.24倍,T2时期在FN中的表达量是XY中的7.12倍(图3)。基因LOX的高表达不利于α-亚麻酸的积累,与图1中所显示的XY中α-亚麻酸含量高于FN的结果相一致。
-
沙棘种子中富含不饱和脂肪酸,其中,α-亚麻酸含量一般较高。α-亚麻酸作为人体必需脂肪酸,具有多种有益的生理功能,因此,为了进一步提高沙棘种子中α-亚麻酸含量,培育沙棘新品种和良种,在分子水平上研究沙棘中α-亚麻酸的合成代谢规律至关重要。亚油酸是α-亚麻酸合成的底物,脱饱和生成亚麻酸[17-18]。本研究选取的2个沙棘品种中,α-亚麻酸/亚麻酸的比值存在显著差异,XY种子中α-亚麻酸是含量最高的脂肪酸,亚油酸含量次之,而在FN中正相反,含量最高的脂肪酸是亚油酸,α-亚麻酸含量次之。因此,基于这2个品种存在的差异挖掘沙棘种子中调控亚油酸向α-亚麻酸转化的关键基因以及抑制α-亚麻酸分解的关键基因,探究α-亚麻酸合成代谢规律。
FAD2催化油酸转化为亚油酸[19-21],FAD2基因表达量下降使亚油酸合成受阻,进而影响α-亚麻酸的生成,而FAD2基因的高表达可以为α-亚麻酸的合成提供充足的底物。2个沙棘品种种子中的亚油酸含量在T2时期快速积累,与T2时期基因FAD2的表达水平急剧上升相一致。在XY和FN种子中,T1和T2时期之间的亚油酸含量差异显著。T3时期基因FAD2的表达量降低,亚油酸含量积累速度明显下降。
α-亚麻酸在FAD3和FAD7的催化下由亚油酸脱饱和产生[17-18]。FAD3与FAD7同属于ω-6 型脂肪酸去饱和酶,已有的研究表明,基因FAD3在植物种子α-亚麻酸的合成过程中起到关键作用[22-24]。实验表明,拟南芥的FAD3基因在大豆中异位表达可使大豆中α-亚麻酸含量增加,大豆的FAD3C基因在芝麻中特异性表达同样也可使芝麻中α-亚麻酸的含量显著提高[25]。此外,Wu等将麻风树的JcFAD3基因在拟南芥的种子中特异性表达使拟南芥种子中α-亚麻酸的含量提高了20.50%~24.94%;同时亚油酸的含量相比野生型降低了11.4%~23.5%[26]。在烟草中过表达拟南芥FAD7基因,同样使烟草中α-亚麻酸的含量升高且亚油酸含量下降[23]。
在T1和T2时期,XY中基因FAD3和基因FAD7的表达量远高于FN,尤其在T2时期,基因FAD3在XY和FN中的表达差异显著,在XY中的表达量是FN中的13.63倍,调控大量亚油酸脱饱和生成α-亚麻酸,因此,XY种子中α-亚麻酸的含量大于亚油酸的含量,而FN中亚油酸含量比α-亚麻酸高。这解释了为何基因FAD2在XY种子中的表达量高于FN,但亚油酸含量却是FN种子大于XY种子。
基因LOX是α-亚麻酸代谢过程中的关键基因,是调控α-亚麻酸转化为其他物质的第一步,调控α-亚麻酸氧化,生成脂肪酸氢过氧化物[27-29]。基因LOX3.1和基因LOX3.2在XY和FN发育的T1时期表达量处于3个时期的最大值,抑制α-亚麻酸的积累,协同基因FAD2、FAD3、FAD7在T1时期的低表达水平,导致T1时期α-亚麻酸含量处于低水平,T2时期基因LOX3.1和LOX3.2表达量下调,基因FAD2、FAD3、FAD7表达量急剧上升,因而α-亚麻酸含量在T2时期显著增长。同时,基因LOX3.1和LOX3.2在FN种子中的表达量高于XY种子,抑制FN种子中α-亚麻酸的积累,使FN种子中α-亚麻酸积累量低于XY。
沙棘种子中α-亚麻酸的高积累源于多个基因的协同调控作用,基因FAD2、FAD3、FAD7的高表达协同基因LOX3.1和LOX3.2的低表达使α-亚麻酸的高积累得以实现。
本研究在基因和代谢水平上研究沙棘中α-亚麻酸合成代谢规律,对进一步提高沙棘种子中α-亚麻酸含量、培育沙棘良种具有重要意义。
-
通过对2个不同沙棘品种种子中中长链脂肪酸和转录组数据联合分析得出:沙棘种子中α-亚麻酸的高积累源于多个基因的协同调控作用。基因FAD2调控油酸去饱和生成亚油酸,其高表达为α-亚麻酸的合成提供了充足的底物。α-亚麻酸在基因FAD3和基因FAD7的调控下由亚油酸去饱和产生,基因FAD3和基因FAD7的高表达促进亚油酸向α-亚麻酸转化,使α-亚麻酸积累,同时使亚油酸含量下降。沙棘种子中基因LOX3.1和LOX3.2是α-亚麻酸代谢过程中的关键基因,其表达水平下降有利于种子中α-亚麻酸的积累。
不同沙棘品种种子中α-亚麻酸含量差异及相关基因分析
Analysis of α-Linolenic Acid Content and Related Genes in Seeds of Sea Buckthorn Cultivars
-
摘要:
目的 探讨α-亚麻酸含量存在差异的2个沙棘品种种子中的基因调控差异,为进一步提高沙棘种子中α-亚麻酸含量、培育沙棘良种奠定基础。 方法 以2个不同品种的沙棘种子为研究对象,通过气相色谱-质谱分析和转录组测序分析确定2个沙棘品种种子中的α-亚麻酸含量差异以及相关差异表达基因。 结果 蒙古大果沙棘(Hippophae rhamnoides ‘Mongolia’向阳)(XY)和中国沙棘(H. rhamnoides ‘Sinensis’丰宁)(FN)种子中的α-亚麻酸含量占比不同,XY中含量最高的脂肪酸是α-亚麻酸,其次是亚油酸,而FN中α-亚麻酸含量次于亚油酸。半成熟期(T2),基因FAD2和基因FAD3在XY中的表达量均显著高于FN,分别是FN的3.83倍和13.63倍。未成熟期(T1)和T2时期基因FAD7在XY中的表达量均显著高于在FN中的表达量,分别是FN的2.09倍和1.72倍。相反,T2时期基因LOX3.1和LOX3.2在FN中的表达量均高于XY,分别是XY的8.02倍和7.12倍。 结论 沙棘种子中α-亚麻酸的高积累源于多个基因的协同作用,基因FAD2、FAD3、FAD7的高表达和基因LOX3.1和LOX3.2的低表达共同调控α-亚麻酸的高积累。 Abstract:Objective To study the differences of gene regulation in seeds of two sea buckthorn (Hippophae rhamnoides) cultivars with different α-linolenic acid content and to find the method of further increase the content of α-linolenic acid in H. rhamnoides seed. Method The difference in α-linolenic acid content and the expressed genes in the seeds of two H. rhamnoides cultivars were analyzed and determined by gas chromatography-mass spectrometry and transcriptomics. Result The proportion of α-linolenic acid in H. rhamnoides 'Mongolia' (XY) and H. rhamnoides ‘Sinensis’ (FN) seeds was different. The fatty acid with the highest content of XY was α-linolenic acid, followed by linoleic acid. The content of α-linolenic acid in FN was lower than that of linoleic acid. In semi-mature stage (T2), the expression levels of gene FAD2 and gene FAD3 in XY were 3.83 times and 13.63 times that in FN respectively. The expression of gene FAD7 in XY was 2.09 times and 1.72 times that in FN in immature stage (T1) and T2respectively. It was consistent with the trend of linoleic acid and α-linolenic acid content. On the contrary, the expressions of gene LOX3.1 and gene LOX3.2 in FN were 8.02 times and 7.12 times that of XY in T2 respectively. Conclusion The high accumulation of α-linolenic acid in H. rhamnoides seeds is due to the synergistic effect of multiple genes. High expression of gene FAD2, FAD3, FAD7 and low expression of gene LOX3.1, LOX3.2 contribute to the accumulation of α-linolenic acid. -
图 1 沙棘XY与FN种子中主要脂肪酸组分的含量变化
Figure 1. Distribution of main fatty acid components in XY and FN seeds of sea buckthorn
图 2 主成分分析与差异表达基因上调和下调分布情况
Figure 2. PCA and up-regulated and down-regulated distribution of DEGs
图 3 沙棘种子发育过程中与α-亚麻酸合成相关的基因表达模式
Figure 3. Gene expression profiles of Alpha linolenic acid biosynthetic genes during seed development of sea buckthorn
图 4 与α-亚麻酸生物合成与代谢相关的途径
Figure 4. Pathway related to the biosynthesis and metabolism of α-linolenic acid
表 1 XY与FN 2个沙棘品种种子在不同发育时期脂肪酸含量的动态变化
Table 1. Dynamic changes of fatty acid composition in the seeds of XY and FN at different developmental stages
μg·g−1 组分
CompositionsXY FN T1 T2 T3 T1 T2 T3 C10:0 1.04±0.09 b 1.75±0.13 ab 2.43±0.69 a 1.04±0.07 b 1.72±0.20 ab 2.24±0.20 a C12:0 1.11±0.27 a 2.06±0.78 a 5.62±0.32 a 0.34±0.40 a 1.34±0.17 a 2.09±1.25 a C13:0 0.08±0.05 b 0.30±0.08 ab 0.35±0.188 a 0.08±0.02 b 0.31±0.10 ab 0.48±0.10 a C14:0 20.70±2.32 c 40.23±2.638 a 53.98±18.17 a 17.66±1.34 c 40.85±7.66 a 58.91±7.54 a C14:1 136.34±14.20 a 145.86±15.94 a 122.48±14.74 a 121.50±9.22 a 121.08±9.35 a 117.06±7.36 a C15:0 6.20±1.43 c 19.42±1.16 b 28.45±10.91 ab 6.84±0.91 c 24.40±5.20 b 37.00±5.97 a C15:1 16.90±2.09 a 14.37±2.21 ab 8.59±0.85 c 14.65±0.78 ab 12.06±4.19 abc 10.52±1.83 bc C16:0 1 219.70±135.84 b 2 337.41±132.23 a 2 873.02±915.18 a 1 141.08±69.79 b 2 482.63±282.95 a 3 165.25±346.54 a C16:1 38.04±6.73 c 70.55±12.28 bc 112.18±36.82 ab 28.40±4.84 c 86.75±22.13 ab 126.20±25.12 a C17:0 7.89±1.39 b 16.93±1.34 a 19.21±6.58 a 9.02±0.85 b 15.92±1.62 a 21.51±2.86 a C17:1 66.01±7.57 a 51.99±6.67 ab 47.64±2.13 ab 56.48±12.46 ab 44.60±3.13 b 51.85±7.10 ab C18:0 695.59±62.14 c 1 330.90±97.82 ab 1 656.18±553.21 a 634.31±23.71 c 1 035.51±73.38 bc 1 280.61±137.68 ab C18:1 78.99±20.21 d 1 824.10±207.78 c 3 447.79±1 190.81 a 101.66±20.47 d 2 197.36±133.31 bc 3 029.64±464.58 ab C18:2 311.23±62.81 d 4 231.52±246.51 c 6 879.55±2 327.19 ab 265.09±48.17 d 5 028.36±863.33 bc 7 443.04±795.32 a C18:3N3 298.54±76.45 c 4 980.34±261.85 ab 7 463.53±2 636.16 a 223.36±46.56 c 4 307.42±690.29 b 6 526.07±969.49 ab C20:0 19.50±3.83 c 102.61±13.30 b 200.82±74.68 a 16.14±2.06 c 94.96±7.90 b 139.33±19.39 b C20:1 7.42±1.60 c 50.74±4.18 b 90.17±30.25 a 5.87±0.58 c 53.50±4.71 b 83.41±16.89 a C20:2 0.93±0.22 c 8.69±0.52 b 14.62±5.51 ab 0.81±0.13 c 9.72±3.29 b 16.89±3.08 a C21:0 3.85±0.72 b 8.44±0.77 ab 11.93±3.97 a 3.76±0.51 b 8.15±1.48 ab 11.94±2.08 a C20:3N3 1.90±0.21 bc 3.02±0.23 ab 3.86±0.93 a 1.53±0.28 c 2.51±0.55 bc 3.94±0.72 a C20:5N3 0 a 0.15±0.08 a 1.04±0.79 a 0 a 0.33±0.03 a 0.87±0.61 a C22:0 12.20±1.62 c 36.59±4.23 b 57.26±20.45 ab 14.29±2.13 c 44.80±9.54 b 68.61±10.83 a C22:1N9 24.60±3.00 ab 29.29±6.16 a 26.30±5.35 ab 16.34±0.58 b 21.45±3.73 ab 20.85±4.84 ab C23:0 2.00±0.44 c 10.73±1.28 b 18.25±7.01 a 2.41±0.44 c 9.76±2.18 b 15.84±3.27 ab C24:0 4.74±0.60 c 17.42±2.00 b 29.66±10.75 a 5.35±0.46 c 18.09±3.08 b 26.69±4.54 ab C24:1 4.05±3.29 a 11.48±0.91 a 6.68±3.95 a 4.38±3.87 a 3.94±3.01 a 4.47±1.55 a C22:6N3 2.62±0.11 c 3.43±0.05 a 3.25±0.25 ab 2.55±0.08 c 3.07±0.09 b 3.26±0.18.00 ab 注:同行不同字母代表显著差异(P< 0.05)。
Note: Different letters in the same line represent significant differences(P< 0.05) 下载: 导出CSV
下载: 导出CSV
表 2 T2时期XY与FN种子中的C18:3N3/C18:2
Table 2. C18:3n3 / C18:2 in XY and FN seeds of sea buckthorn at T2
品种
CultivarC18:3N3/C18:2 T2 T2-1 T2-2 T2-3 XY 1.182a 1.21 1.17 1.16 FN 0.862b 0.92 0.75 0.92 注:不同字母代表显著差异(P<0.05),T2-1、T2-2、T2-3代表T2时期的3个生物学重复样本。
Notes: Different letters in the same column represent significant differences(P<0.05).T2-1、T2-2 and T2-3represent three biological replicates of the T2 period.
下载: 导出CSV
-
[1] 臧茜茜, 邓乾春, 从仁怀, 等. 沙棘油功效成分及药理功能研究进展[J]. 中国油脂, 2015, 40(5):76-81. doi: 10.3969/j.issn.1003-7969.2015.05.017 [2] Tiitinen K M, Hakala M A, Kallio H P. Quality components of Sea buckthorn (<italic>Hippophae rhamnoides</italic>) varieties[J]. J Agric Food Chem, 2005, 53(5): 1692-1699. doi: 10.1021/jf0484125 [3] Tiitinen K M, Yang B, Haraldsson G G,<italic> et al</italic>. Fast analysis of sugars, fruit acids, and vitamin C in Sea buckthorn (<italic>Hippophae rhamnoides</italic> L.) varieties[J]. J Agric Food Chem, 2006, 54(7): 2508-2513. doi: 10.1021/jf053177r [4] Andersson S C, Olsson M E, Johansson E,<italic> et al</italic>. Carotenoids in Sea buckthorn (<italic>Hippophae rhamnoides</italic> L.) berries during ripening and use of pheophytin a as a maturity marker[J]. Journal of Agricultural & Food Chemistry, 2009, 57(1): 250-258. [5] Kanayama Y, Kato K, Stobdan T,<italic> et al</italic>. Research progress on the medicinal and nutritional properties of Sea buckthorn (<italic>Hippophae rhamnoides</italic>)-a review[J]. Journal of Horticultural Science and Biotechnology, 2012, 87(3): 203-210. doi: 10.1080/14620316.2012.11512853 [6] Marta S M, Cuenca A P. The impact of Sea buckthorn oil fatty acids on human health[J]. Lipids in Health and Disease, 2019, 18(1): 145-157. doi: 10.1186/s12944-019-1065-9 [7] Das U. Essential fatty acids - a review[J]. Current Pharmaceutical Biotechnology, 2006, 7: 467-482. doi: 10.2174/138920106779116856 [8] Leaf A. Omega-3 fatty acids and prevention of arrhythmias[J]. Current Opinion in Lipidology, 2007, 18(1): 31-34. doi: 10.1097/MOL.0b013e328012d61b [9] Wang C, Harris W S, Mei C,<italic> et al</italic>. n 3 Fatty acids from fish or fish-oil supplements, but not -linolenic acid, benefit cardiovascular disease outcomes in primary- and secondary-prevention[J]. Am J Clin Nutr, 2006, 84(1): 5-17. doi: 10.1093/ajcn/84.1.5 [10] 姚振纯, 刘继德. 大豆脂肪酸组分与改良[J]. 大豆通报, 1997(1):14-14. [11] 迟晓元, 郝翠翠, 潘丽娟, 等. 不同花生品种脂肪酸组成及其积累规律的研究[J]. 花生学报, 2016, 45(3):32-36. [12] Yang B, Kallio H P. Fatty acid composition of lipids in Sea buckthorn (<italic>Hippophae rhamnoides</italic> L.) berries of different origins[J]. Journal of Agricultural & Food Chemistry, 2001, 49: 1939-1947. [13] Ingrid E J T, Chanderbali A S, Gitzendanner M A,<italic> et al</italic>. Modified CTAB and TRIzol protocols improve RNA extraction from chemically complex embryophyta[J]. Applications in Plant Sciences, 2015, 3(5): 1400105. doi: 10.3732/apps.1400105 [14] Wang L, Feng Z, Wang X,<italic> et al</italic>. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data[J]. Bioinformatics, 2009, 26: 136-138. [15] Young M D, Wakefield M J, Smyth G K,<italic> et al</italic>. Gene ontology analysis for RNA-seq: accounting for selection bias[J]. Genome biology, 2010, 11(2): 1-12. [16] Mao X, Cai T, Olyarchuk JG,<italic> et al</italic>. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary[J]. Bioinformatics, 2005, 21(19): 3787-3793. doi: 10.1093/bioinformatics/bti430 [17] Jadhav A, Katavic V, Marillia E F,<italic> et al</italic>. Increased levels of erucic acid in <italic>Brassica carinata</italic> by co-suppression and antisense repression of the endogenous FAD2 gene[J]. Metabolic Engineering, 2005, 7(3): 215-220. doi: 10.1016/j.ymben.2005.02.003 [18] Niu B, Guo L, Zhao M,<italic> et al</italic>. Molecular cloning, characterization, and expression of an ω-3 fatty acid desaturase gene from <italic>Sapium sebiferum</italic>[J]. Journal of Bioscience & Bioengineering, 2008, 106(4): 375-380. [19] DellaPenna, D. Plant metabolic engineering[J]. Plant Physiology, 2001, 125(1): 160-163. doi: 10.1104/pp.125.1.160 [20] Huang J, Zhang T, Zhang Q,<italic> et al</italic>. The mechanism of high contents of oil and oleic acid revealed by transcriptomic and lipidomic analysis during embryogenesis in <italic>Carya cathayensis </italic>Sarg<italic></italic>[J]. BMC Genomics, 2016, 17(1): 113-131. doi: 10.1186/s12864-016-2434-7 [21] 李 丽, 崔顺立, 穆国俊, 等. 高油酸花生遗传改良研究进展[J]. 中国油料作物学报, 2019, 41(6):986-997. [22] Liang L H, Jie Y Z, Li X,<italic> et al</italic>. Identification and evaluation of ω-3 fatty acid desaturase genes for hyperfortifying α-linolenic acid in transgenic rice seed[J]. Journal of Experimental Botany, 2012, 63(8): 3279-3287. doi: 10.1093/jxb/ers051 [23] Hiroaki K, Tatsurou H, Gorou H,<italic> et al</italic>. Genetic enhancement of cold tolerance by expression of a gene for chloroplast ω-3 fatty acid desaturase in transgenic tobacco[J]. Plant Physiology, 1994, 105: 601-605. doi: 10.1104/pp.105.2.601 [24] 韩 平, 阮成江, 丁 健, 等. 紫斑牡丹种子高积累碳十八不饱和脂肪酸的多基因调控[J]. 分子植物育种, 2019, 17(7):2101-2108. [25] Bhunia R K, Chakraborty A, Kaur R,<italic> et al</italic>. Seed-specific increased expression of 2S albuminpromoter of sesame qualifies it as a useful genetic tool for fatty acid metabolic engineering and related transgenic intervention in sesame and other oil seed crops[J]. Plant Molecular Biology, 2014, 86: 351-365. doi: 10.1007/s11103-014-0233-6 [26] Wu P, Zhang S, Zhang L,<italic> et al</italic>. Functional characterization of two microsomal fatty acid desaturases from <italic>Jatropha curcas</italic> L.[J]. Journal of Plant Physiology, 2013, 170(15): 1360-1366. doi: 10.1016/j.jplph.2013.04.019 [27] Kühn H, Borchert A. Regulation of enzymatic lipid peroxidation: the interplay of peroxidizing and peroxide reducing enzymes[J]. Free Radical Biology and Medicine, 2002, 33(2): 154-172. doi: 10.1016/S0891-5849(02)00855-9 [28] 陈 昊, 谭晓风. 基于油脂合成期油桐种仁转录组数据的α-亚麻酸代谢途径解析[J]. 林业科学, 2015, 51(3):41-48. [29] Xie D, Dai Z, Yang Z,<italic> et al</italic>. Combined genome-wide association analysis and transcriptome sequencing to identify candidate genes for flax seed fatty acid metabolism[J]. Plant science: an international journal of experimental plant biology, 2019, 286: 98-107. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 4705
- HTML全文浏览量: 2028
- PDF下载量: 66
- 被引次数: 0
