


-
竹蝗属(Ceracris)隶属直翅目(Orthoptera)蝗总科(Acridoidea)网翅蝗科(Arcypteridae),是东洋区的特有种昆虫。在国外主要分布于东南亚地区,包括越南、老挝、缅甸和泰国等国家,在我国主要分布在秦岭一淮河以南的地区,包括云南、广西、四川、江苏、安徽、福建等地区[1]。
目前有记载的竹蝗属昆虫有17种[2],其中以黄脊竹蝗(Ceracris kiangsu Tsai)和青脊竹蝗(Ceracris nigricornis Walker)危害最严重,其若虫和成虫以取食竹叶为主,幼竹被危害后会死亡,成竹受灾后2~3 a不萌发新笋,被啃食过的竹林会大面积枯死,竹叶如同火烧状枯黄[3]。近年来,青脊竹蝗在我国的分布范围最广,种群数量最多[4];而黄脊竹蝗从境外入侵造成的危害更大。2020年6月,黄脊竹蝗经中老边境大规模迁飞入侵我国云南西双版纳、普洱、红河等多个地区,发生面积高达3 118.47 hm2[5],发生严重区域密度高达200~800头·m−2[6]。云南省与老挝有着相似的气候条件和植被类型[7],境外黄脊竹蝗的入侵对我国边境地区的生态环境安全和粮食安全造成了巨大的威胁。
竹蝗属昆虫是东洋区特有种,所属的种类在我国基本都有分布,因此世界范围内关于竹蝗的研究大都集中在国内,研究主要集中在单种竹蝗属昆虫生物学特性[8-9],食性分析[10-12]以及防治策略[13-16]上,关于相似种间形态学及生物学的比较鲜有研究。近年来竹蝗灾害依旧在发生,尤其是2020年境外黄脊竹蝗大规模侵入,对我国林业尤其是竹类造成了严重的经济损失,对我国生态安全造成了严重威胁。准确识别不同林业害虫并明确其生物学特性,是制定害虫针对性预报监测和科学防治的基础[17],蝗虫的分类鉴定主要是依靠成虫的外部形态特征,包括蝗虫的体色外观、纹路、体长大小及其他部位的大小比例来进行。汪清清[2]等通过几何测量技术和形态数据聚类分析对黄脊竹蝗和青脊竹蝗的体长、前后翅大小以及后足股节进行了比对,同时还利用线粒体COⅠ和Cyt b基因建树分析,基于形态学及分子遗传学区分了两种竹蝗。
欧晓红[4]等调查发现,竹蝗属害虫通常同域混生,而对同域内不同的竹蝗种类进行分类鉴定是进行防治工作的必要前提。故本文作者采集了同一地理区域(四川长宁)混生的黄脊竹蝗和青脊竹蝗成虫并在室内饲养,从外观特征、重要部位的大小比例、外生殖器等进行了比较分析;同时比较了两种竹蝗的活动规律、交尾时间以及取食习性等,以期明晰竹蝗属两种混合发生种群之间形态学、生物学的异同,为第一线林农和基层技术人员制定两种竹蝗针对性、精准性的防控策略提供基础依据,研究成果在森林保护工作中具有重要现实意义。
-
本实验于2022年6月开展,将采集四川省宜宾市长宁县的黄脊竹蝗与青脊竹蝗成虫饲养于云南省昆明市的云南省林业和草原科学院的实验大棚内。作者团队在前期研究中发现,黄脊竹蝗若虫喜食无芒雀麦(Bromus inermis Leyss.)[18],查阅文献资料得知成虫喜食毛竹(Phyllostachys edulis (Carrière) J. Houz.)、芭蕉(Musa basjoo Siebold),因此,本研究选择饲养所用的植物为无芒雀麦、毛竹、芭蕉。其中无芒雀麦草种购置于北京正道种业有限公司,并分批种植在实验大棚内,以保证饲草的供应。毛竹和芭蕉均采集于云南省林业和草原科学院后山。
-
由于长宁的地理位置和昆明存在差异,为了确定黄脊竹蝗是否适合在昆明进行饲养观察,提前半个月对长宁和昆明实验大棚内的相关指标进行测定。于每日的8点、10点、12点、14点、17点和20点,对饲养环境的温度和湿度进行记录,并根据长宁与昆明的差异性来作出相应的调整。
将采集的黄脊竹蝗成虫和青脊竹蝗成虫分别饲养在长1 m宽1 m高1.5 m且底部镂空侧面和顶部有拉链开关的养虫笼中,每笼各饲养150只。养虫笼底部接地,在笼子底部铺上一层薄膜,在薄膜上铺厚度8 cm粒度40目的砂壤土,并在四周用土掩盖笼子和地面之间的缝隙。饲养挑取新鲜且完整无病害的无芒雀麦、毛竹和芭蕉,并在饲养前用清水洗净,同时将3种植物插入有水的锥形瓶中并放入养虫笼内。每日更换。
-
对黄脊竹蝗和青脊竹蝗成虫的基本外部特征进行观察记录,包括体色、纹路等;体长、触角、后足腿节以及头幅进行测量记录;外生殖器拍照记录。
-
准备6个长35 cm宽35 cm高35 cm的养虫笼,随机抓取雌雄成虫各5只,共10只放入1个养虫笼中。黄脊竹蝗和青脊竹蝗各3笼,饲养方法同1.2。使用电子数显游标卡尺(0~200 mm)对两种竹蝗成虫的触角、后足股节和头幅进行测量并做好相应的记录。再分别于交尾前、交尾怀卵后、产卵后3个时间段使用电子数显游标卡尺(0~200 mm)对黄脊竹蝗和青脊竹蝗的体长进行测量记录,成虫死亡后实验结束。
-
从养虫笼中抓取黄脊竹蝗和青脊竹蝗雌雄成虫,立即放入事先准备好的毒瓶(瓶中装有乙酸乙酯浸润的棉花)中,蝗虫毒死后,放入含有75%乙醇的瓶中分装,带回实验室。用2号昆虫针将制作好的蝗虫标本固定在泡沫板上后,再拿到体式显微镜(SZN71宁波舜宇)下观察拍照。
-
分别观察饲养在长1 m宽1 m高1.5 m规格的养虫笼和长35 cm宽35 cm高35 cm规格的养虫笼中的黄脊竹蝗和青脊竹蝗的活跃程度,并做好相应的记录。
-
为观察黄脊竹蝗和青脊竹蝗交尾日节律,在进入交尾盛季后,准备6个长50 cm宽50 cm高50 cm的养虫笼,黄脊竹蝗和青脊竹蝗各3笼,每个养虫笼饲养20只成虫,且保持雌雄比例和体长大小一致。从早上8点至晚上20点,每隔1 h对黄脊竹蝗和青脊竹蝗的交尾情况进行观察,并做好相应的记录。重复观察3 d并记录好当天的天气状况。
-
对黄脊竹蝗成虫和青脊竹蝗成虫的食量进行测定,准备6个长50 cm宽50 cm高50 cm的养虫笼,黄脊竹蝗和青脊竹蝗各3笼,每个养虫笼饲养20只成虫,且保持雌雄比例和体长大小一致。每次试验进行前,对供试虫进行12 h的饥饿处理。将无芒雀麦、毛竹、芭蕉各25 g随机等距摆放于养虫笼内,每次试验的时间控制在24 h。试验于养虫室内进行,控制温度25 ± 3 ℃,湿度60%~70%。每次试验结束后用网格纸测量统计并记录被取食叶片的缺刻面积。
-
本试验数据整理采用Excel,数据处理、方差分析以及制图采用SPSS软件。
-
观察黄脊竹蝗成虫图1(a)的形态特征,黄脊竹蝗成虫体色以绿色和黄色为主色调,额顶凸出呈锐角三角形状,有一逐渐加宽的黄色纵纹由额顶延伸至前胸背板;触角呈黑色丝状且顶端为黄色,复眼为深黑色的卵圆形;后足腿节整体呈黄绿色,且中部有整齐排列的“人”字形状的褐色沟纹;胫节呈蓝黑色且有成排的刺。

图 1 黄脊竹蝗与青脊竹蝗成虫
Figure 1. C. kiangsu and C. nigricornis adults
青脊竹蝗成虫图1(b)的形态特征,青脊竹蝗成虫体色以深绿色或翠绿色为主色调,头部、胸部两侧板以及前胸背板均为绿色,头顶和三角形的面部呈一锐角;为黑色丝状触角,后足腿节与胫节连接处有一淡黄斑圈,胫节黑色,有褐色斑圈。
对比黄脊竹蝗成虫与青脊竹蝗成虫形态特征后发现:黄脊竹蝗身体以黄、绿色为主,青脊竹蝗身体呈翠绿或暗绿色;黄脊竹蝗前胸两侧板至两前翅的前缘中域内外缘边为黑褐色,青脊竹蝗则是从额顶触角窝至两前翅末端均为黑褐色;黄脊竹蝗额顶至前胸背板有一逐渐加宽的黄色纵纹,而青脊竹蝗这个部位均为翠绿色,并无黄色纵纹,这也是黄脊竹蝗和青脊竹蝗的最大区别。
-
黄脊竹蝗与青脊竹蝗成虫在不同阶段的体长测量结果如表1所示。由表1可知,黄脊竹蝗雌性成虫平均体长为35~38 mm,雄性成虫为30~33 mm;青脊竹蝗雌性成虫平均体长为34~36 mm,雄性成虫24~29 mm。同种竹蝗成虫之间比较时,雌性成虫与雄性成虫的平均体长差异显著(p<0.05);两种竹蝗之间比较时,雄性成虫的平均体长差异显著(p<0.05),雌性成虫交尾前与产卵后平均体长差异显著(p<0.05),交尾怀卵后平均体长差异不显著(p>0.05)。
表 1 黄脊竹蝗成虫与青脊竹蝗成虫体长比较
Table 1. Comparison of body length of adult C. kiangsu and C. nigricornis mm
种类
Species黄脊竹蝗 青脊竹蝗 交尾前
Precopulation♀ 36.55 ± 1.42 Ac 34.63 ± 0.44 Bc ♂ 31.35 ± 0.75 Aa 26.82 ± 2.77 Ba 怀卵后
Pregnant with eggs♀ 42.87 ± 2.88 Aa 41.27 ± 0.92 Aa ♂ 30.40 ± 1.20 Ab 24.89 ± 1.38 Bb 产卵后
Postspawning♀ 38.44 ± 1.78 Ab 36.23 ± 0.95 Bb ♂ 31.12 ± 0.98 Aa 23.68 ± 1.45 Bc 注:表中数值都为平均值 ± 标准误差。同行大写字母表示组间差异性,同列小写字母表示组内差异性
Notes: Data are means ± SD.The same uppercase letter indicates the difference between groups, and the lowercase letter in the same column indicates the difference within a group黄脊竹蝗与青脊竹蝗成虫的触角、后足股节及头幅的长度如表2所示。黄脊竹蝗雌性成虫触角平均长度22~25 mm,后足股节平均长度21~22 mm,头幅平均长度5~6 mm;雄性成虫触角平均长度23~24 mm,后足股节平均长度17~18 mm,头幅平均长度3~4 mm。青脊竹蝗雌虫成虫触角平均长度17~19 mm,后足股节平均长度17~20 mm,头幅平均长度4~5 mm;雄性成虫触角平均长度16~19 mm,后足股节平均长度15~16 mm,头幅平均长度3~4 mm。同种竹蝗雌雄成虫之间比较,黄脊竹蝗雌性成虫与雄性成虫触角差异不显著(p>0.05),后足股节差异显著(p<0.05),头幅差异显著(p<0.05);青脊竹蝗雌性成虫与雄性成虫触角差异不显著(p>0.05),后足股节差异显著(p<0.05),头幅差异显著(p<0.05)。两种竹蝗之间比较,雌雄成虫触角、后股足节、头幅差异均显著(p<0.05)。
表 2 黄脊竹蝗成虫与青脊竹蝗成虫触角、后足股节和头幅比较
Table 2. Comparison of antenna, hind femur and head width of adult C. kiangsu and C. nigricornis mm
种类
Species触角
Antenna后足股节
Hind femur头幅
Head width黄脊竹蝗 ♀ 23.46 ± 1.29 a 21.68 ± 0.28 a 5.33 ± 0.2 a ♂ 23.22 ± 0.9 a 17.86 ± 0.23 c 4.08 ± 0.19 b 青脊竹蝗 ♀ 18.34 ± 0.56 b 18.67 ± 1.62 b 4.61 ± 0.18 b ♂ 17.75 ± 1.58 b 16.09 ± 0.47 d 3.5 ± 0.29 c 注:表中数值都为平均值 ± 标准误差。同列小写字母表示组间差异性
Notes: Data are means ± SD.The same column of lowercase letters indicates differences between groups -
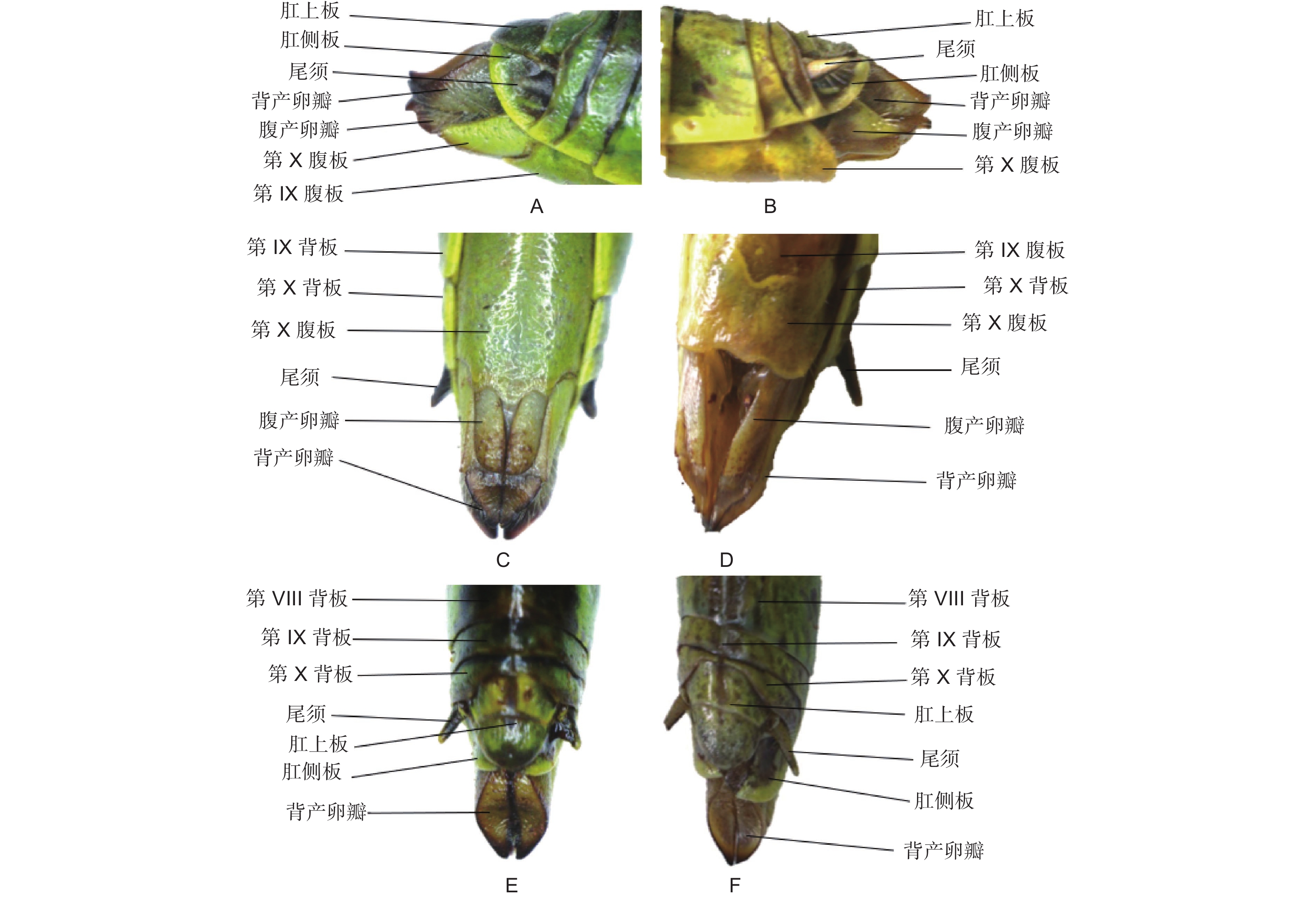
黄脊竹蝗雌成虫的外生殖器如图2所示,又称产卵器,整体呈管状构造,具有一定的伸缩性,主要由3对产卵瓣组成,分别是着生于第8腹节的腹产卵瓣和着生于第9腹节的内产卵瓣和背产卵瓣。其中内产卵瓣退化较严重,承担产卵功能的主要是腹产卵瓣和背产卵瓣。黄脊竹蝗雄成虫的外生殖器如图3所示,又称交配器,整体看起来是由下生殖板向上弯曲包裹形成的球面,在交配时承担挟持雌虫作用的抱握器和将精子进送入雌虫体内的阳具都被下生殖板包裹其中。黄脊竹蝗雌雄成虫外生殖器均含有尾须结构。
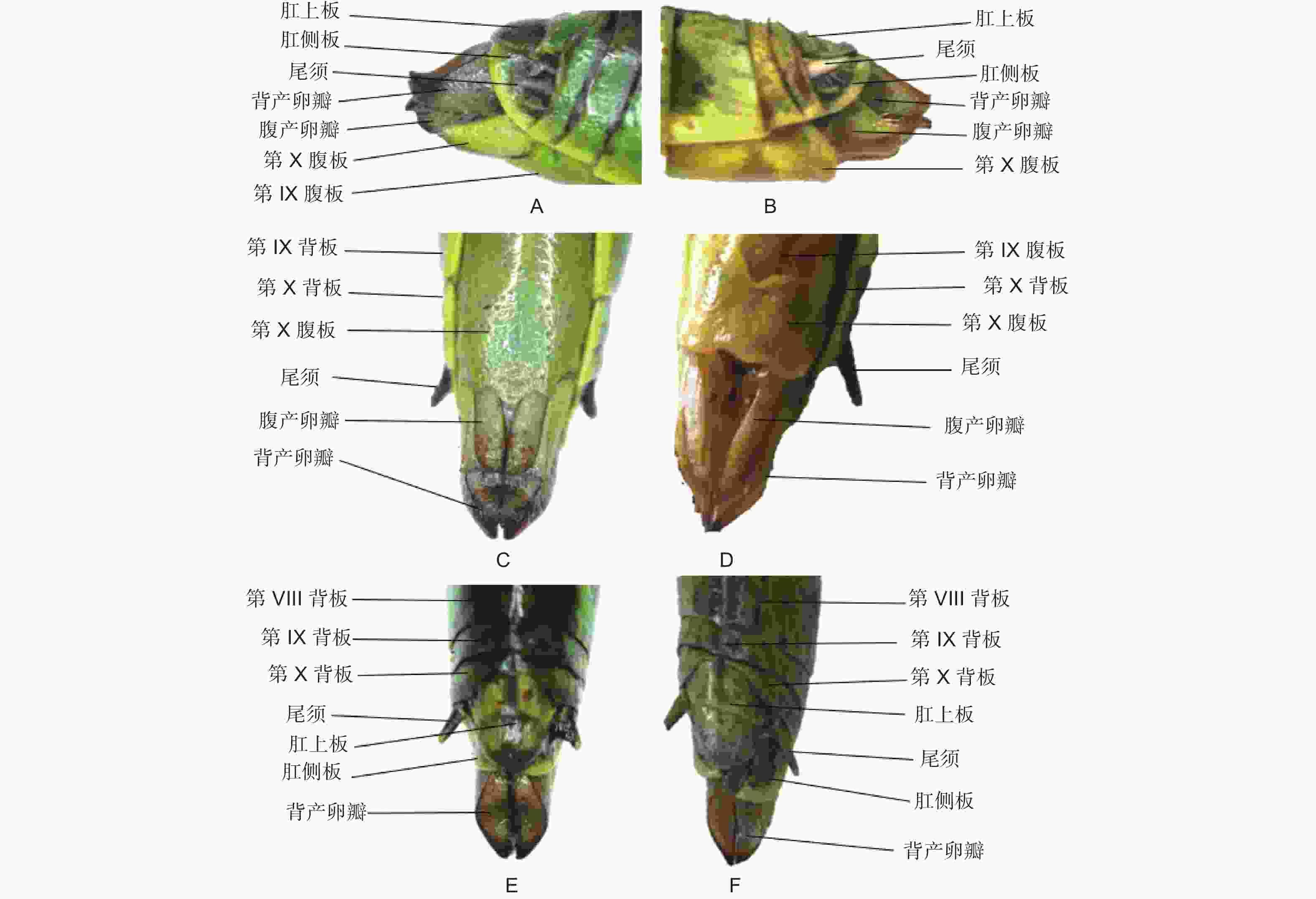
图 2 黄脊竹蝗与青脊竹蝗雌性成虫外生殖器比较
Figure 2. Comparison of external genitalia of female adult C. kiangsu and C. nigricornis
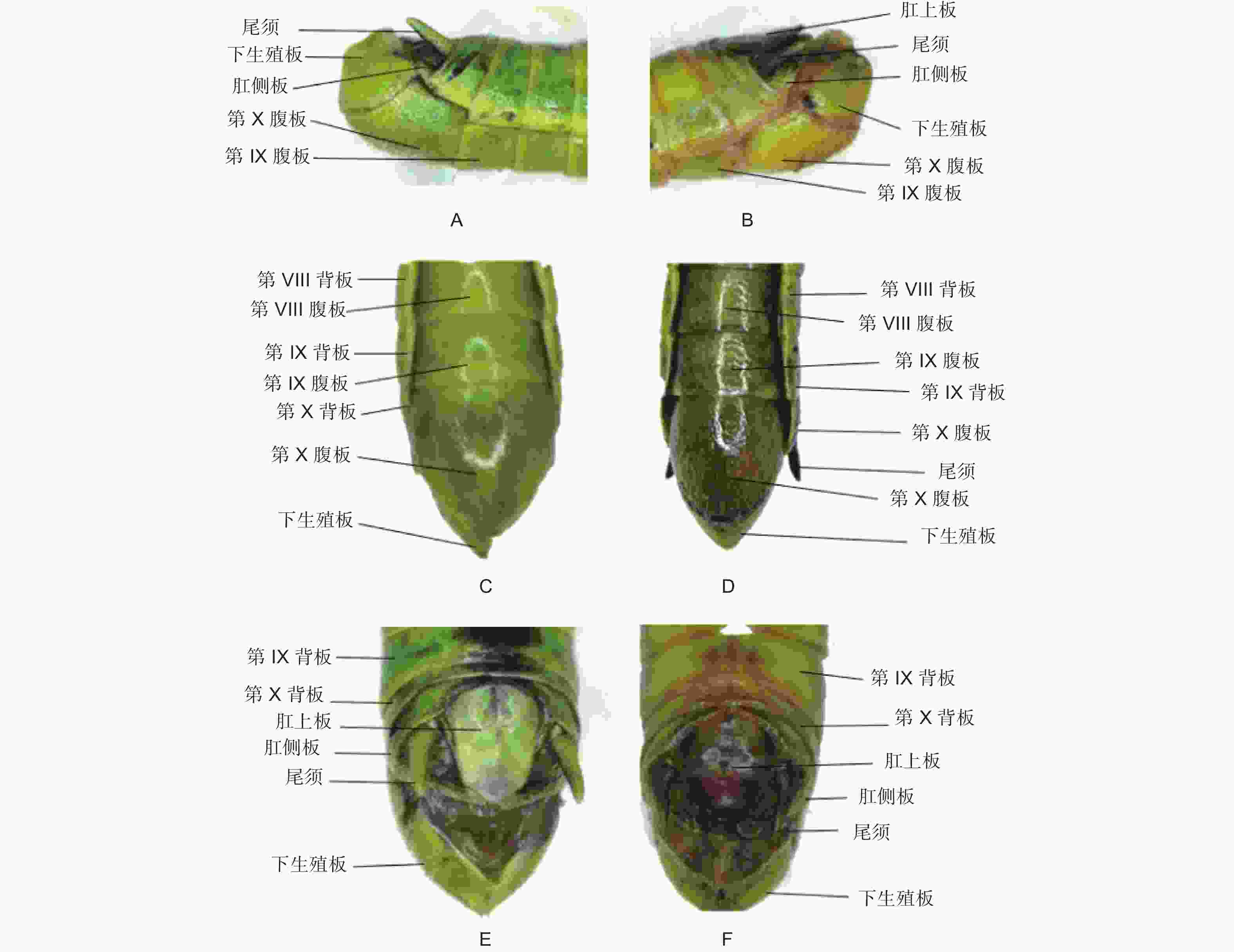
图 3 黄脊竹蝗与青脊竹蝗雄性成虫外生殖器比较
Figure 3. Comparison of external genitalia of male adult C. kiangsu and C. nigricornis
经过观察后发现,青脊竹蝗雌雄成虫的外生殖器与黄脊竹蝗雌雄成虫的外生殖器结构基本一致,因此这里不再做描述。
-
经过观察后发现,黄脊竹蝗和青脊竹蝗饲养于长1 m宽1 m高1.5 m规格的养虫笼中的活跃程度更高;饲养于长35 cm宽35 cm高35 cm规格养虫笼中的活跃度较低。此外,饲养过程中还观察到黄脊竹蝗和青脊竹蝗的活跃时间段有所区别,总结后发现,在上午的时间段,更能观察到较多的黄脊竹蝗进行跳跃、爬动等活动;而青脊竹蝗进行活动的时间段更多是集中在下午的时间段。
-
黄脊竹蝗成虫与青脊竹蝗成虫在交尾方式上并无太大差别,均是雄虫趴在雌虫背上,再用前、中两足抱住雌虫的胸部,雄虫生殖器向下弯曲插入雌虫生殖器进行交配,交尾时间一般持续60~70 min,且都存在一雌多雄的交尾情况。交尾成功后,雄虫会一直趴在雌虫背上,雌虫四处活动包括休息、取食时都不分开,此时需避免外界的刺激,一旦外界刺激过大,就会终止交尾。两种竹蝗均存在交配回避的现象,即雌虫受精怀卵后,雄虫再趴在雌虫背上时,雌虫会用后足踢背上的雄虫,并将生殖器官向下弯曲,拒绝雄虫的配对。
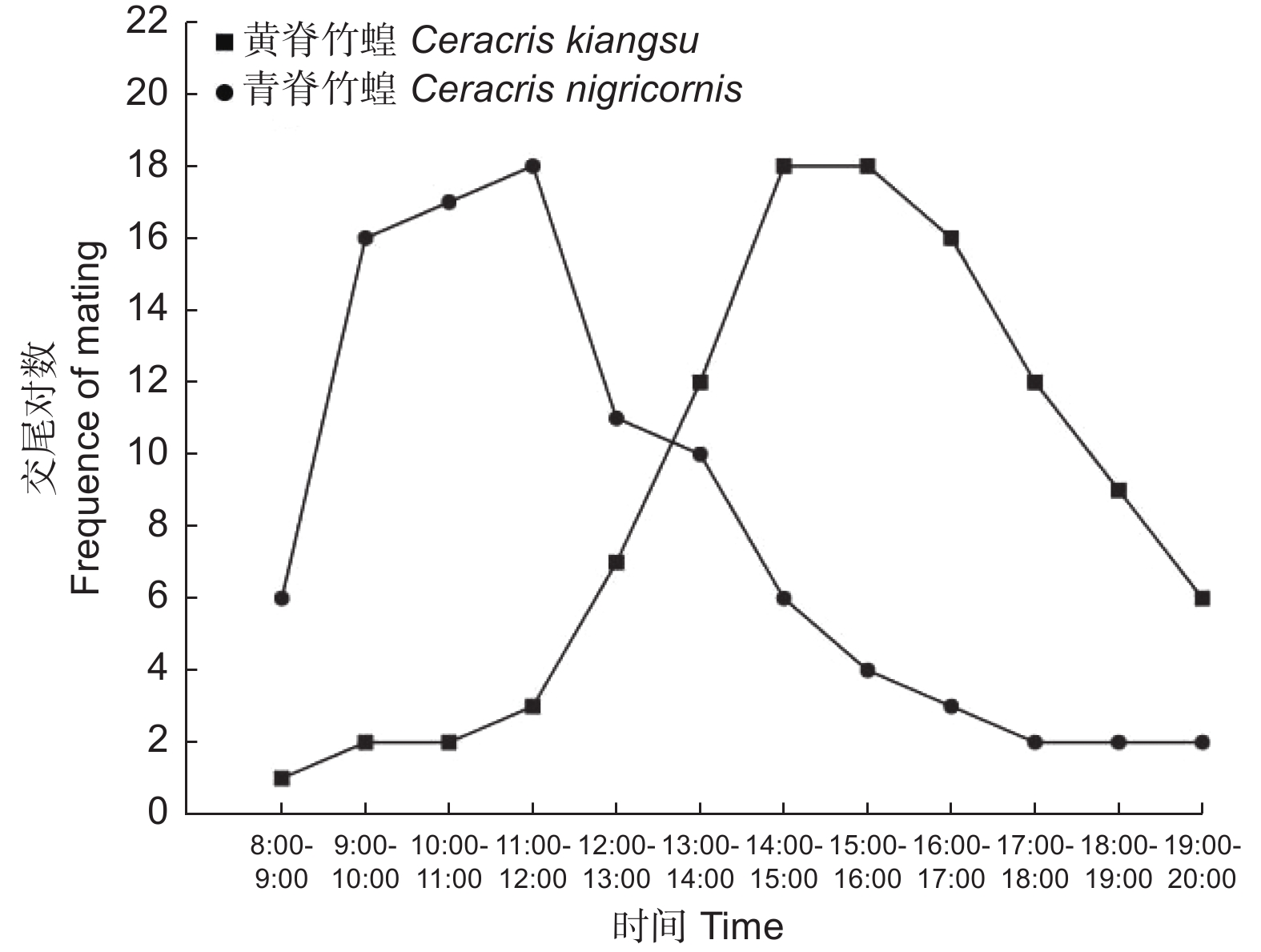
交尾节律的观察于2022年8月17日,18和19日连续3 d进行,将3 d累计交尾的竹蝗对数为指标,当天的环境温度分别为23~28 ℃,19~35 ℃,20~26 ℃。由图4可以看出,两种竹蝗成虫交尾的偏好时间不一样。青脊竹蝗成虫交尾的高峰期多在早晨9时至12时;而和黄脊竹蝗成虫交尾的高峰期多在下午14时至17时。除了交尾高峰期的时间段,在其他时间段也有零星的竹蝗进行交尾。
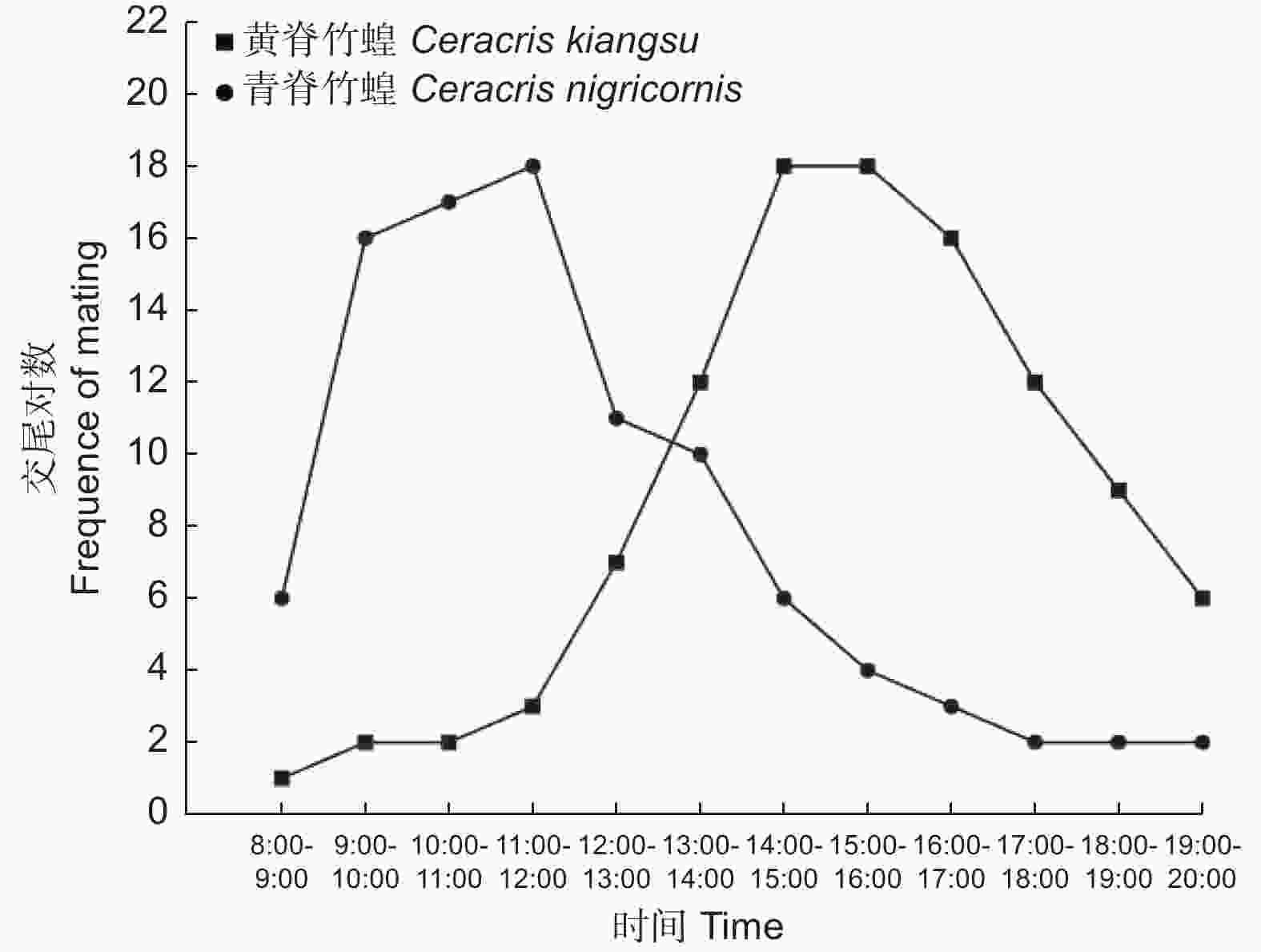
图 4 黄脊竹蝗和青脊竹蝗成虫在3 d内不同时段内的交尾情况
Figure 4. Mating situation of adult C. kiangsu and C. nigricornis in different time periods within 3 d
-
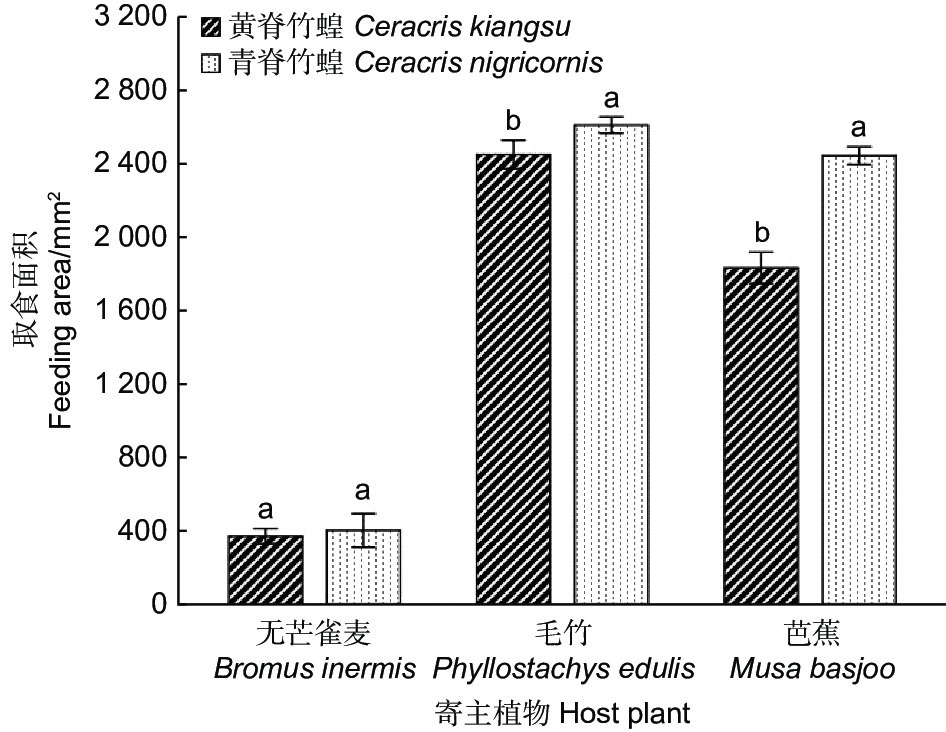
为了研究黄脊竹蝗与青脊竹蝗对不同寄主植物的取食偏好性,本实验还对黄脊竹蝗与青脊竹蝗对3种不同寄主植物的取食面积大小进行了研究,实验结果如图5所示。结果表明黄脊竹蝗与青脊竹蝗对芭蕉的取食面积差异显著(p<0.05),其中黄脊竹蝗对芭蕉的平均取食面积为1 834.72 mm2,青脊竹蝗的平均取食面积为2 446.09 mm2;对毛竹的取食面积差异也显著(p<0.05),黄脊竹蝗对毛竹的平均取食面积为2 452.14 mm2,青脊竹蝗对毛竹的平均取食面积为2 612.53 mm2;而对无芒雀麦的取食差异不显著(p>0.05),平均取食面积均在380 mm2左右。
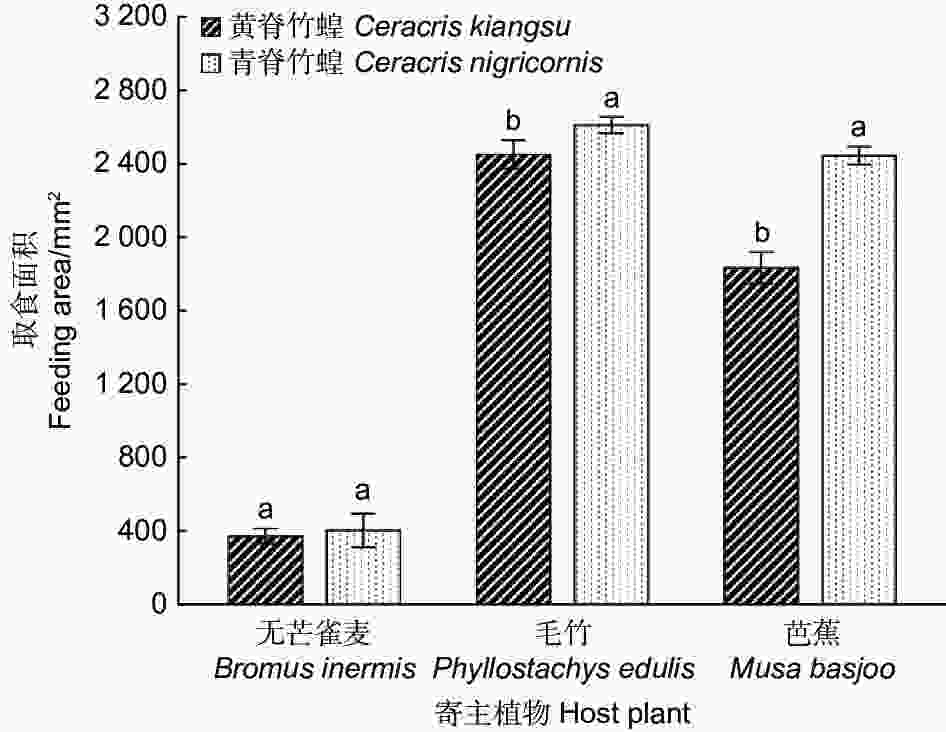
图 5 黄脊竹蝗和青脊竹蝗成虫对3种不同寄主植物的取食偏好
Figure 5. Feeding preferences of adult C. kiangsu and C. nigricornis on three different host plants
对比黄脊竹蝗与青脊竹蝗对不同寄主植物的取食面积可以发现,黄脊竹蝗与青脊竹蝗均喜食芭蕉与毛竹,对无芒雀麦的取食量较小。
-
蝗虫的鉴定主要依据于其成虫的形态特征,动物界雌雄形态及体型大小普遍存在差异,蝗虫也不例外,体型大的雌性对后代繁殖或许更加有利[19-20]。本研究结果表明,黄脊竹蝗和青脊竹蝗两种竹蝗雌性成虫的体型均大于雄性成虫,且雌性成虫在交尾怀卵后的阶段体长还会增加5~8 mm左右,这是因为雌虫怀卵后,腹部会变得膨大,长度甚至会超过翅长。而在雌虫产完卵后,体长会在怀卵阶段的基础上缩短5~6 mm,但是仍比交尾前阶段增加了2~3 mm,推测可能是雌虫产卵时产卵管伸长后发生了一定的塑性形变。对于触角、后足股节以及头幅这3个形态特征指标,雌雄成虫在不同发育阶段差异不显著。分析认为,本研究是围绕成虫展开实验观察,而成虫的发育已经较为稳定,故在成虫的不同发育阶段中并无显著变化。同一物种的昆虫由于生存环境的不同,在长期进化过程中会产生不同的地理种群,体型的差异是地理种群间反映环境因子作用效应最直观的特征[2]。本实验除饲养来自四川长宁的黄脊竹蝗,还一并饲养了采自西双版纳勐腊县的黄脊竹蝗。比较来自不同地域的黄脊竹蝗后发现,勐腊的黄脊竹蝗体型小于长宁的黄脊竹蝗,单从形态方面来说,推测可能因为地理隔离产生了不同的地理种群,出现了地理亚种,作者将在下一步工作基于分子层面进一步进行验证该推测。
昆虫种群的延续主要依靠雌雄个体的交配行为及交配的成功率[21],个体间的交配成功与两性的交配活动以及交配的昼夜节律密切相关[22],同域发生的昆虫,为了避免交配时被干扰,同时提高其交配成功率,种间会达成一种默契,从而在交配行为节律出现相互错峰的现象[23]。本实验发现两种竹蝗的交尾喜好时间有所不同,青脊竹蝗成虫交尾高峰期主要是在早晨9:00—12:00时,该时间段平均温度为20 ℃,平均湿度为70%;黄脊竹蝗成虫交尾高峰期主要是在下午14:00—17:00时,该时间段的平均温度为30 ℃,平均湿度为50%。两种竹蝗均是采自同一地理区域,而交尾高峰出现时间段不同,应该是同域两种竹蝗为了避免交配时被干扰,同时提高其交配成功率的一种交配行为策略,此外,两种竹蝗间,为了提高同域发生时种间的生存率,其本身的日节律在长期进化过程中也出现了相互有所不同,表现出相互间对环境温湿度的适应性有所不同。交配是生物界最重要的行为过程,关乎生物自身种群繁衍壮大,本研究确认了,同域发生的两种竹蝗交配高峰期相互错开,非常有利于其交配的成功率,非常有利其下一代种群的延续,这是两种竹蝗长期进化形成的利于种群大发展的一种生存策略。实验还发现,不同地域的黄脊竹蝗出现交尾高峰期的月份不一致,如长宁的黄脊竹蝗是在8月中下旬,而勐腊的黄脊竹蝗出现在9月中下旬,推测可能是勐腊的黄脊竹蝗发育历期比长宁的黄脊竹蝗晚,故相应的交尾高峰也迟一些。由于实验条件的限制,该推测还需进一步验证。
植食性昆虫的取食行为和寄主植物本身的特性密切相关,寄主植物的物理形态,营养成分等都会影响植食性昆虫的取食选择[10],不仅如此,当同时存在几种不同的寄主植物时,昆虫的取食行为具有一定的选择性[24]。实验表明,两种竹蝗的取食习性基本一致,均喜食毛竹和芭蕉,而对无芒雀麦的取食量较小。作者团队研究表明,蝗蝻期黄脊竹蝗喜食草本植物,如:无芒雀麦,不喜食竹类植物(孔维维等,待发表),蝗蝻期青脊竹蝗可能也表现出与黄脊竹蝗同样的趋势。竹蝗对氮素取食有偏好性,而毛竹中N含量较高[10],这也使得竹蝗喜食毛竹。然而在提供了毛竹的前提下,竹蝗对芭蕉的取食量也依旧很大,这也说明竹蝗对芭蕉有着潜在的威胁,故在野外除加强对竹林的监控以外,也需加强对芭蕉林的监测。实验还发现青脊竹蝗的取食量远大于黄脊竹蝗,分析原因可能是青脊竹蝗的发育历期较黄脊竹蝗更长,更需要营养物质的补充,故取食量也更大一些。
-
综上所述,黄脊竹蝗与青脊竹蝗在形态特征和生活习性上既存在着一定的相似性,如生殖器外部形态特征、交尾方式和取食偏好等;但又有一定的差异性,如交尾喜好时间和取食量大小等。因此在防治手段上,既可以利用两种竹蝗之间的相似性设计通用的防治方法,又可以利用差异性来进行针对性的防治,或是两者相结合以达到更好的防治效果。此外,本实验仅是在室内大棚对两种竹蝗进行饲养与观察,可能会与野外观察到的情况有些差异,故在进行防治工作时,应将室内大棚观测的结果与野外实际情况相结合,更加高效精准的对竹蝗进行防控,更好的保护我国林业资源。
同域两种竹蝗成虫形态学及生物学特性比较
Comparison of Morphological and Biological Characteristics of Two Species of Ceracris Insects Adults in the Same Area
-
摘要:
目的 为比较同域混合发生的两种竹蝗属昆虫,即黄脊竹蝗与青脊竹蝗形态学与生物学之间的异同点。 方法 采自同一地理区域的两种竹蝗的成虫,在大棚内饲养,对其形态特征:体长、触角、后足股节、头幅、外生殖器等,及生活习性:活动规律、交尾习性、寄主植物取食习性等进行观察记录和统计分析。 结果 结果表明,黄脊竹蝗雌成虫体长36.55 ± 1.42 mm,触角23.46 ± 1.29 mm,后足股节21.68 ± 0.28 mm,头幅5.33 ± 0.2 mm;雄成虫体长31.35 ± 0.75 mm,触角23.22 ± 0.9 mm,后足股节17.86 ± 0.23 mm,头幅4.08 ± 0.19 mm。青脊竹蝗雌成虫体长34.63 ± 0.44 mm,触角18.34 ± 0.56 mm,后足股节18.67 ± 1.62 mm,头幅4.61 ± 0.18 mm;雄成虫体长26.82 ± 2.77 mm,触角17.75 ± 1.58 mm,后足股节16.09 ± 0.47 mm,头幅3.5 ± 0.29 mm。同种竹蝗间雌雄成虫体长差异显著,两种竹蝗比较时雌雄体长、触角、后足股节以及头幅差异显著,雌性成虫的体长变化趋势相似,雌雄外生殖器结构相似。黄脊竹蝗交尾高峰期为14:00—17:00时,青脊竹蝗交尾高峰期为9:00—12:00时。两种竹蝗成虫在面对3种不同的寄主植物时表现出相同的取食偏好,均是最喜食毛竹,其次是芭蕉,最后是无芒雀麦,但青脊竹蝗的取食量大于黄脊竹蝗。 结论 两种竹蝗形态和生物学习性表现出的相似性和差异性是两种竹蝗能够同域发生的基础,研究结果为监测和防控两种竹蝗提供了一定的基础信息支持。 Abstract:Objective To compare the similarities and differences between the morphology and biology of two insects of the Ceracris, Ceracris kiangsu Tsai and Ceracris nigricornis Walker. Method The adults of two kinds of Ceracris collected from the same geographical area were reared in a greenhouse, and their morphological characteristics: body length, antennae, hind femur, head width, external genitalia, etc., and living habits: activity rules, mating habits. The feeding habits of host plants were observed, recorded and statistically analyzed. Result The results showed that the body length of the female adult C. kiangsu was 36.55 ± 1.42 mm, the antennae was 23.46 ± 1.29 mm, the hind femur was 21.68 ± 0.28 mm, and the head width was 5.33 ± 0.2 mm; the body length of the male adults was 31.35 ± 0.75 mm, the antennae was 23.22 ± 0.9 mm, the hind femur was 17.86 ± 0.23 mm,and the head width was 4.08 ± 0.19 mm. The body length of the female adult of C. nigricornis was34.63 ± 0.44 mm, the antennae was 18.34 ± 0.56 mm, the hind femur was 18.67 ± 1.62 mm, and the head width was 4.61 ± 0.18 mm; the body length of the male adult was 26.82 ± 2.77 mm, the antennae was 17.75 ± 1.58 mm, the hind femur was 16.09 ± 0.47 mm, the head width was 3.5 ± 0.29 mm. There were significant differences in body length between male and female adults of the same species of Ceracris insects. There were significant differences in body length, antennae, hind femur, and head width between the two types of Ceracris insects. The peak mating period of the C. kiangsu was 14-17 o'clock, and the peak mating period of the C. nigricornis was 9-12 o'clock. Two kinds of Ceracris adults showed the same feeding preference when faced with three different host plants, and they all preferred to eat bamboo (Phyllostachys edulis), followed by plantain (Musa basjoo), and finally brome (Bromus inermis), but the C. nigricornis consumed more food than the C. kiangsu. Conclusion The similarity and difference in morphology and biological learning of the two kinds of Ceracris insects are the basis for the sympatric occurrence of the two kinds of Ceracris insects, which provides some basic information support for the monitoring and control of the two kinds of Ceracris insects. -
Key words:
- Ceracris kiangsu
- / Ceracris nigricornis
- / Mating style
- / Mating rhythm
- / Feeding preference
-
图 2 黄脊竹蝗与青脊竹蝗雌性成虫外生殖器比较
Figure 2. Comparison of external genitalia of female adult C. kiangsu and C. nigricornis
图 3 黄脊竹蝗与青脊竹蝗雄性成虫外生殖器比较
Figure 3. Comparison of external genitalia of male adult C. kiangsu and C. nigricornis
图 4 黄脊竹蝗和青脊竹蝗成虫在3 d内不同时段内的交尾情况
Figure 4. Mating situation of adult C. kiangsu and C. nigricornis in different time periods within 3 d
图 5 黄脊竹蝗和青脊竹蝗成虫对3种不同寄主植物的取食偏好
Figure 5. Feeding preferences of adult C. kiangsu and C. nigricornis on three different host plants
表 1 黄脊竹蝗成虫与青脊竹蝗成虫体长比较
Table 1. Comparison of body length of adult C. kiangsu and C. nigricornis mm
种类
Species黄脊竹蝗 青脊竹蝗 交尾前
Precopulation♀ 36.55 ± 1.42 Ac 34.63 ± 0.44 Bc ♂ 31.35 ± 0.75 Aa 26.82 ± 2.77 Ba 怀卵后
Pregnant with eggs♀ 42.87 ± 2.88 Aa 41.27 ± 0.92 Aa ♂ 30.40 ± 1.20 Ab 24.89 ± 1.38 Bb 产卵后
Postspawning♀ 38.44 ± 1.78 Ab 36.23 ± 0.95 Bb ♂ 31.12 ± 0.98 Aa 23.68 ± 1.45 Bc 注:表中数值都为平均值 ± 标准误差。同行大写字母表示组间差异性,同列小写字母表示组内差异性
Notes: Data are means ± SD.The same uppercase letter indicates the difference between groups, and the lowercase letter in the same column indicates the difference within a group 下载: 导出CSV
下载: 导出CSV
表 2 黄脊竹蝗成虫与青脊竹蝗成虫触角、后足股节和头幅比较
Table 2. Comparison of antenna, hind femur and head width of adult C. kiangsu and C. nigricornis mm
种类
Species触角
Antenna后足股节
Hind femur头幅
Head width黄脊竹蝗 ♀ 23.46 ± 1.29 a 21.68 ± 0.28 a 5.33 ± 0.2 a ♂ 23.22 ± 0.9 a 17.86 ± 0.23 c 4.08 ± 0.19 b 青脊竹蝗 ♀ 18.34 ± 0.56 b 18.67 ± 1.62 b 4.61 ± 0.18 b ♂ 17.75 ± 1.58 b 16.09 ± 0.47 d 3.5 ± 0.29 c 注:表中数值都为平均值 ± 标准误差。同列小写字母表示组间差异性
Notes: Data are means ± SD.The same column of lowercase letters indicates differences between groups
下载: 导出CSV
-
[1] 程 佳, 潘涌智, 舒金平. 黄脊竹蝗研究现状及进展[J]. 山东林业科技, 2010, 40(2):116-119. [2] 汪青青. 竹蝗属几何形态测量学研究及青脊竹蝗种下分化[D]. 西安: 陕西师范大学, 2019. [3] 汤兆勇. 黄脊竹蝗的发生及综合防治[J]. 现代农业科技, 2019(1):118-119. [4] 欧晓红, 陈 方, 和秋菊. 竹蝗属物种特性比较研究[J]. 林业科学研究, 2000, 13(4):385-390. [5] 卓富彦, 李红梅, 吕建平, 等. 2020年云南农区黄脊竹蝗应急防控策略及展望[J]. 中国植保导刊, 2021, 41(5):99-101. [6] 卓富彦, 朱景全, 任彬元, 等. 2020 年云南省黄脊竹蝗发生防控初报[J]. 中国植保导刊, 2020, 40(8):60-62. [7] 杨学礼, 李红卫, 卓富彦, 等. 黄脊竹蝗在云南江城的发生与防控[J]. 中国植保导刊, 2022, 42(6):53-55. [8] 孙立峰, 奚月明, 居 峰, 等. 黄脊竹蝗生物学特性及综合防控实践[J]. 安徽农业科学, 2016, 44(29):139-141. [9] 张威. 黄脊竹蝗趋泥行为驱动力及泥源搜索机制研究[D]. 北京: 中国林业科学研究院, 2016. [10] 张守科, 张 威, 舒金平, 等. 黄脊竹蝗取食偏好与竹叶品质的关系[J]. 生态学杂志, 2017, 36(8):2266-2273. [11] 金 京, 谢 榕, 李 霞, 等. 3种不同寄主植物对黄脊竹蝗取食偏好性和生长发育的影响[J]. 浙江农林大学学报, 2020, 37(6):1143-1148. [12] 王永芬, 徐胜涛, 番华彩, 等. 黄脊竹蝗在香蕉和芭蕉上的取食特性分析[J]. 热带农业科学, 2021, 41(7):68-74. [13] LIU D Z, ZHAO S Y, YANG X M, et al. Radar monitoring unveils migration dynamics of the yellow-spined bamboo locust (Orthoptera: Arcypteridae)[J]. Computers and Electronics in Agriculture, 2021, 187: 1-8. [14] 许春枝, 洪宜聪, 郭宝宝, 等. 2种植物源农药烟雾对黄脊竹蝗的防治效果[J]. 世界竹藤通讯, 2021, 19(2):34-42. [15] 韩伟君, 胡 彦, 马莉莉, 等. 2020年云南省黄脊竹蝗发生规律初探[J]. 中国植保导刊, 2022, 42(1):98-100. [16] 泽桑梓, 蔡茂伟, 林 晓, 等. 2020年云南省黄脊竹蝗发生与防控[J]. 中国森林病虫, 2021, 40(4):41-43. [17] 陆宴辉, 吴孔明. 我国棉花盲蝽生物学特性的研究进展[J]. 应用昆虫学报, 2012, 49(3):578-584. [18] 孔唯唯. 云南热性灌草丛黄脊竹蝗食性研究[D]. 昆明: 云南农业大学, 2021. [19] WILKINS M R, SEDDON N, SAFRAN R J. Evolutionary divergence in acoustic signals: causes and consequences[J]. Trends in Ecology and Evolution, 2013, 28: 156-166. doi: 10.1016/j.tree.2012.10.002 [20] MHATRE N, MONTEALEGRE Z F, BALAKRISHNAN R, et al. Changing resonator geometry to boost sound power decouples size and song frequency in a small insect[J]. Biophysics and Computational biology, 2012, 109(22): 1444-1452. [21] XU T, HANSEN L, TEALE S A. Mating and adult feeding behaviour influence pheromone production in female Asian longhorn beetle Anoplophora glabripennis (Coleoptera: Ceramby-cidae)[J]. Agricultural and Forest Entomology, 2020, 23(3): 276-286. [22] 刘 伟, 彭 媛, 黄丽莉, 等. 草地贪夜蛾交配行为及繁殖的观察[J]. 江西农业大学学报, 2023, 45(1):53-60. [23] 戴长庚, 李鸿波, 张昌容, 等. 贵州水稻二化螟成虫羽化和交配节律研究[J]. 杂交水稻, 2020, 35(3):79-82. [24] 李 超, 程登发, 郭文超, 等. 不同寄主植物对马铃薯甲虫的引诱作用[J]. 生态学报, 2013, 33(8):2410-2415. -

 点击查看大图
点击查看大图

计量
- 文章访问数: 958
- HTML全文浏览量: 451
- PDF下载量: 18
- 被引次数: 0
