

-
紫楠(Phoebe sheareri (Hemsl.) Gamble)系樟科(Lauraceae)楠属(Phoebe)中经济价值极高的中国特有珍贵树种,也是商品木材“金丝楠木”的原植物种之一。其木材坚硬致密,纹理清晰,有光泽,耐腐且富楠木芳香,是高级家具、雕刻的理想用材;同时其枝叶繁茂且常绿,可作为园林绿化树种。紫楠喜温暖、湿润气候,常见于海拔1000 m以下的山地沟谷阔叶林中[1-2],其生境范围较广,位于106.38°~121.45°E、24.50°~32.05°N,在江苏、浙江、安徽、江西、福建、湖南、贵州和广西等省区均有自然散生状分布[3-4]。然而,因气候变迁,人为采伐、开垦,自然更新能力较弱等原因,现存紫楠天然种质资源已接近枯竭。据对天然分布区实地调查,较大群落仅存于浙江省的杭州市灵隐、建德、淳安、临安的部分区域[5-6]。
生态位是群落生态学研究的重要理论基础,通过量化物种之间以及物种与环境之间的关系,用于探究种内关系、种间关系以及物种在群落中所处地位[7-9]。生态位宽度反映物种对环境资源的利用状况,也可以体现物种的生态适应性和分布幅度[10]。生态位重叠是指一定资源序列上,两个物种由于利用同级资源而相互重叠的情况,生态位重叠较大的两个种往往具有相似的生态学和生物学特性[11-12]。种间联结指不同物种在空间分布上的关联性,这种关联性是各物种在不同生境中相互作用所形成的,表现出特定的种间关系或生态习性上的异同[13]。目前国内外对紫楠的研究主要集中在种子休眠机制及萌发特征[14-16]、幼苗光合生理特性[17]、叶片形态与生长特征[18-19]、群落组成[20-21]等方面,有关紫楠种群生态位与种间联结性的研究还未见报道。本研究基于野外样地调查,对紫楠天然分布相对集中的天目山脉区域内6个典型群落进行生态位与种间联结性分析,探讨紫楠群落种间关系和演替动态,揭示紫楠对群落内资源的利用状况,为紫楠珍贵资源的保护与利用提供科学依据。
-
天目山脉位于浙江、安徽两省的交界处(118°36′~120°06′E、29°52′~30°55′N),山脉走向呈东北-西南方向,地势由南向北逐渐降低,一般海拔为100~1000 m,最高峰清凉峰的海拔1787 m。该区土壤主要为黄红壤,中、高山区还分布有山地黄壤和山地黄棕壤,多为酸性土壤。该区北部为北亚热带气候,年均气温15.4℃,年均降水量1300 mm,年均空气相对湿度80%。南部属中亚热带气候,年均气温16.0℃,年均降水量1500 mm,年均空气相对湿度80%~82%。该区域植物种类丰富,植物群落多样,是我国华东地区最主要的植物资源储存库之一。
-
对研究区内紫楠天然分布点实地踏勘后,选择具有代表性的6个天然群落进行调查。每个群落设置2个20 m × 20 m的样地,详细调查并记录样地内所有乔木(胸径≥5 cm)树种名称、胸径、高度、冠幅;在每个样地中随机设置3个2 m × 2 m的灌木样方,调查灌木层(包括未满足乔木层测量标准的更新幼树、苗)的植物名称、平均高度、冠幅、盖度及多度[22]。
-
参考国内外植物群落的研究方法[23-24],计算乔木层、灌木层各物种重要值并选取排序前10的物种进行生态位和种间联结性分析。选用Levins指数生态位宽度(BL)、Shannon指数生态位宽度(BS)、Pianka生态位重叠指数(Oik)分析其生态位特征[25-27]。
-
采用Excel 2019整理数据,应用R 4.1.2中spaa程序包进行生态位和种间联结性分析,运用Origin 2021进行绘图。
-
在调查的6个紫楠天然群落的12个样地中,紫楠在乔木层的重要值最大,为30.71%,薄叶润楠(Machilus leptophylla Hand.-Mazz.)、柳杉(Cryptomeria fortunei Hooibrenk)次之,分别为18.02%、12.70%(表1)。紫楠在灌木层的重要值亦最大,为14.69%,其次为薄叶润楠的12.75%和茶(Camellia sinensis (L.) O. Ktze.)的11.77%(表2),是灌木层中的优势种。根据各物种重要值大小选取排序前10的物种计算生态位宽度(表1、表2)。紫楠在群落乔木层、灌木层的Levins生态位宽度和Shannon生态位宽度均最高,说明紫楠在调查群落中对资源利用能力较强,竞争力较强。总体看来,乔木层、灌木层主要物种的BL和BS大小排序基本一致,且与重要值排序基本一致,说明所调查的紫楠天然群落已达到了较稳定阶段。
表 1 紫楠天然群落乔木层主要物种重要值与生态位宽度
Table 1. Important value and niche breadth of dominant species in the arbor layer of P. sheareri community
序号
No.物种名
Species name缩写
Abbreviation重要值
IV/%生态位宽度 Niche breadth BL BS 1 紫楠 P. sheareri Ps 30.71 5.02 1.27 2 薄叶润楠 M. leptophylla Ml 18.02 4.08 1.06 3 柳杉 C. fortunei Cf 12.70 3.15 0.99 4 枫香 Liquidambar formosana Hance Lf 8.91 3.02 0.95 5 木荷 Schima superba Gardn. et Champ. Ss 7.16 2.04 0.84 6 杉木 Cunninghamia lanceolata (Lamb.) Hook. Cl 6.62 2.53 0.89 7 黄果朴 Celtis sinensis Pers. Cs 5.22 2.13 0.70 8 青冈栎 Quercus glauca Thunb. Cg 5.10 2.07 0.61 9 油桐 Vernicia fordii (Hemsl.) Airy Shaw Vf 4.22 2.25 0.59 10 棕榈 Trachycarpus fortunei (Hook.) H. Wendl. Tf 2.69 2.04 0.30 表 2 紫楠天然群落灌木层主要物种重要值与生态位宽度
Table 2. Important value and niche breadth of dominant species in the shrub layer of P. sheareri community
序号
No.物种名
Species name缩写
Abbreviation重要值
IV/%生态位宽度
Niche breadthBL BS 1 紫楠 P. sheareri Ps 14.69 4.17 1.24 2 薄叶润楠 M. leptophylla Ml 12.75 3.89 0.98 3 茶 C. sinensis Cs 11.77 3.14 0.96 4 阔叶箬竹 Indocalamus latifolius (Keng) McClure Il 9.29 3.05 0.89 5 毛柄连蕊茶 Camellia fraterna Hance Cf 5.73 2.74 0.86 6 高粱泡 Rubus lambertianus Ser. Rl 7.20 2.69 0.86 7 紫麻 Oreocnide frutescens (Thunb.) Miq. Of 7.86 2.56 0.81 8 朱砂根 Ardisia crenata Sims Ac 3.15 2.48 0.75 9 香花鸡血藤 Callerya dielsiana (Harms) P. K. Loc ex Z. Wei & Pedley Cd 4.76 2.71 0.79 10 杜茎山 Maesa japonica (Thunb.) Moritzi. ex Zoll. Mj 4.36 2.31 0.56 -
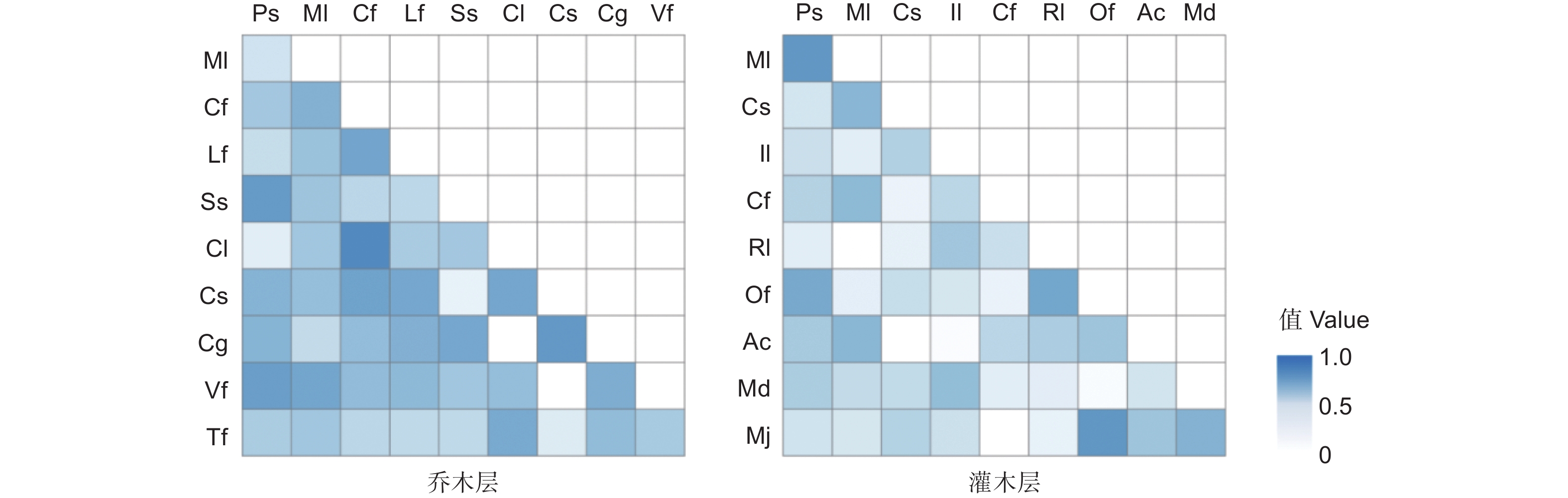
紫楠天然群落乔木层、灌木层优势物种Pianka生态位重叠指数分析表明,在乔木层45组种对中有43组(占95.56%)存在生态位重叠,所有种对Pianka生态位重叠指数的平均值为0.342,重叠指数最大值达0.713,为柳杉和杉木种对,重叠指数大于0.5的共10对(占22.22%),重叠指数小于0.2的共8对(占17.78%)(图1)。此外,乔木层中与紫楠生态位重叠值最大的为木荷(0.591),说明两者间存在一定的竞争关系。在灌木层45组种对中有43组(占95.56%)存在生态位重叠,所有种对Pianka生态位重叠指数的平均值为0.217;重叠指数最大值达0.621,为紫麻和杜茎山种对;重叠指数大于0.5的共3对(占6.67%);重叠指数小于0.2的共25对(占55.56%)。灌木层中植物与紫楠生态位重叠值较大的为薄叶润楠(0.610)和紫麻(0.499)。
图 1 紫楠群落乔木层、灌木层优势物种种间Pianka生态位重叠指数
Figure 1. Pianka niche overlap index among dominant species in the arbor and shrub layer of P. sheareri community
-
乔木层方差比率VR=1.247>1,表明乔木层优势物种间总体呈正联结;乔木层检验统计量W=17.511,介于χ20.95(12)与χ20.05(12)之间(表3),即乔木层优势物种间总体呈不显著正联结。灌木层方差比率VR=0.674<1,表明灌木层优势物种间总体呈负联结;灌木层检验统计量W=41.894,介于χ20.95(36)与χ20.05(36)之间,表明灌木层优势物种间总体呈不显著负联结。
表 3 紫楠群落乔木层、灌木层优势树种总体联结性
Table 3. Overall interspecific associations among dominant species in the arbor layer and shrub layer of P. sheareri community
空间层次
Stand layer方差比率(VR)
Variance ratio检验统计量(W)
Statisticχ2临界值
χ2 threshold检验结果
Results乔木层 1.247 17.511 χ20.95(12)=5.226,χ20.05(12)=21.026 不显著正联结 灌木层 0.674 41.894 χ20.95(36)=23.269,χ20.05(36)=50.998 不显著负联结 -
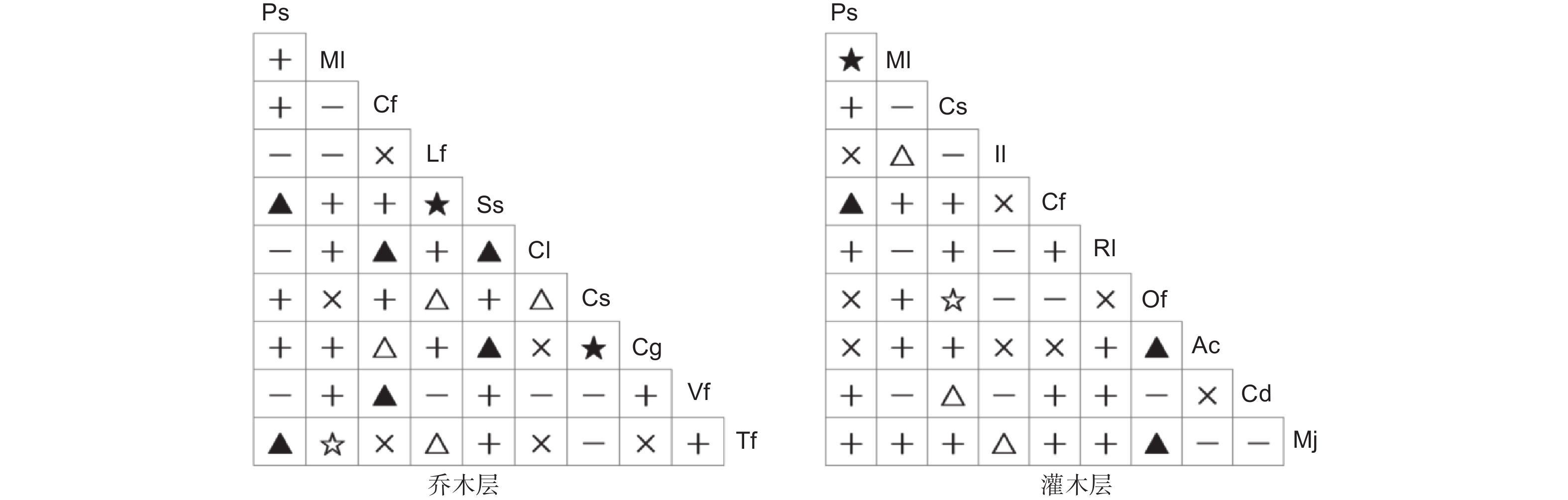
χ2检验结果表明,在紫楠群落乔木层优势物种组成的45个种对中,有正联结为25对、负联结14对、无关联6对,分别占总种对数的55.6%、31.1%和13.3%,正负联结比为1.8∶1(图2)。在25对正联结种对中,极显著正联结2对,分别为枫香-木荷、黄果朴-青冈栎;显著正联结6对,分别为紫楠-木荷、紫楠-棕榈、柳杉-杉木、柳杉-油桐、木荷-杉木、木荷-青冈栎。在14对负联结种对中,极显著负联结1对,为薄叶润楠-棕榈;显著负联结4对,分别为柳杉-青冈栎、枫香-黄果朴、枫香-棕榈、杉木-黄果朴。
图 2 紫楠群落乔木层、灌木层优势物种种间χ2检验半矩阵图
Figure 2. Semi-matrix diagram of χ2 test among dominant species in the arbor and shrub of P. sheareri community
在灌木层优势物种组成的45个种对中,有正联结为22对、负联结15对、无关联8对,分别占总种对数的48.9%、33.3%、17.8%,正负联结比为1.5:1。在22对正联结种对中,极显著正联结1对,为紫楠-薄叶润楠;显著正联结3对,分别为紫楠-毛柄连蕊茶、紫麻-朱砂根、紫麻-杜茎山。在15对负联结种对中,极显著负联结1对,为茶-紫麻;显著负联结3对,分别为薄叶润楠-阔叶箬竹、茶-香花鸡血藤、阔叶箬竹-杜茎山。
-
一般情况下,群落中物种重要值与其所占生态位宽度成正比,生态位宽度反映物种对资源环境的利用与适应情况[30-31]。紫楠的重要值最大,且生态位宽度值高于其他树种,说明其适应于所在群落的生态环境,这与紫楠的生物学特性有关,处于林冠层的紫楠对光照等环境资源的利用能力较强,同时处于中间层的紫楠具有一定耐荫性,能在郁闭度较高的天然阔叶林中生存,也可能与本研究所选群落中紫楠分布相对较集中有关。两物种共同利用或占有同一资源就产生了生态位重叠现象。有研究表明,生态位宽度越大的物种由于其资源利用能力和适应性较强,往往与其他物种的生态位重叠值越高[32-33]。而本研究中物种的生态位宽度与生态位重叠值之间并没有表现出高度相关性,这与舟山群岛次生林[34]、平潭岛台湾相思(Acacia confusa Merr.)群落[35]、君子峰自然保护区观光木(Michelia odora (Chun) Nooteboom & B. L. Chen)群落[36]等的生态位特征相似。例如本研究的乔木层中杉木重要值小于木荷,而生态位宽度却大于木荷,灌木层中毛柄连蕊茶重要值虽小于高粱泡、紫麻,生态位宽度却更大,这与各物种的生物学特性有关,在资源的利用能力上存在较大差异[37]。对崖柏(Thuja sutchuenensis Franch.)[38]、格氏栲(Castanopsis kawakamii Hayata)[39]、浙江楠(Phoebe chekiangensis C. B. Shang)[40]等群落的研究均表明物种间生态位重叠值越大,其竞争关系越大。因此,在保护紫楠种群时,可适当进行人工干预,伐除群落中与其竞争较大的物种,尤其是灌木层中与紫楠生态位重叠较大的薄叶润楠和紫麻,以满足紫楠种群的自然更新。
-
群落随时间不断演替,会逐渐趋向于稳定,群落总体将呈正联结[41-42]。研究表明,群落中优势度较大的物种在一定程度上影响种间关联性程度[43]。本研究中紫楠群落乔木层优势物种间总体呈不显著正联结,而灌木层总体呈不显著负联结,说明乔木层各物种可以和谐共存,对资源的利用也较充分,已处于较稳定状态,而灌木层仍处于不稳定阶段,各物种间竞争较大,具有生态位分离[44]。因此,处于灌木层中的紫楠小苗也容易在群落演替发展过程中被淘汰,较难长成大树;有研究指出,灌木层的种间关系可能受到了乔木层的影响[45]。χ2检验显示乔木层优势物种间正联结种对数多于负联结,正负联结比达1.8∶1,高于灌木层的正负联结比(1.5∶1),且绝大部分种对联结性均不显著,说明紫楠群落乔木层大多数物种间的生态特征较一致,或互相作用有利,但作用强度不高,这符合紫楠群落乔木层已较为稳定的种间关系特征,与秦岭华山松(Pinus armandi Franch.)天然群落特征相似[46]。因此,在无人为干扰和重大自然灾害前提下,紫楠群落能较稳定存在。
-
本研究的天目山脉区域内典型紫楠天然群落中,大部分优势物种的生态特征较相似,其中紫楠具有相对较强的竞争优势,对资源的利用程度较高。群落乔木层优势物种间总体呈不显著正联结,各物种可和谐共存,已处于较稳定阶段,而灌木层仍处于不稳定状态,各物种间存在竞争。因此,在紫楠种群的原地保护时,可适当人工干预,根据种间联结和生态位特征,伐除群落中与其竞争较大的物种,促进自然更新。
紫楠天然群落优势物种生态位与种间联结性
Niche and Interspecific Association of Dominant Species in Phoebe sheareri Natural Community
-
摘要:
目的 为探究紫楠(Phoebe sheareri (Hemsl.) Gamble)天然群落优势物种种间关系和群落稳定性。 方法 基于野外样地调查,从物种组成、重要值、生态位宽度、生态位重叠指数、方差比率法和χ2检验等方面研究紫楠群落乔木层、灌木层各优势物种的生态位特征和种间联结性。 结果 紫楠在调查群落中的重要值最大,且其Levins指数生态位宽度和Shannon指数生态位宽度均最大,说明在群落中资源利用程度较高,竞争优势强;紫楠群落乔木层优势物种间总体呈不显著正联结(VR=1.247,χ20.95(12)>W>χ20.05(12)),而灌木层总体呈不显著负联结(VR=0.674,χ20.95(36)>W>χ20.05(36)),说明乔木层稳定性较高,且高于灌木层;χ2检验结果显示乔木层、灌木层优势物种间正联结种对数均多于负联结,正负联结比分别为1.8∶1和1.5∶1,且绝大部分种对联结性均不显著,表明紫楠群落大多数优势物种间的生态特征较一致。 结论 紫楠群落总体稳定性较好,但仍可进行适当的人工抚育,促进紫楠种群自然更新。 Abstract:Objective In order to explore the interspecific correlations and the stability of Phoebe sheareri natural community. Method Based on field sample survey, the ecological niche characteristics and interspecific associations of dominant species in the arbor layer and shrub layer of P. sheareri community were studied by through species composition, important value, niche breadth, niche overlap index, variance ratio method, and χ2 test. Result The important value, Levins niche breadth, and Shannon-Weaver niche breadth of P. sheareri were the largest in the investigated communities, indicating that both the resource utilization degree and competitive advantage were strong. The dominant species in the arbor layer showed an overall insignificant positive association(VR = 1.247, χ20.95(12) > W > χ20.05(12)), while the shrub layer showed an overall insignificant negative association(VR = 0.674, χ20.95(36) > W > χ20.05(36)), indicating that the stability of the arbor layer was higher than that of the shrub layer. The χ2 test result showed that the positively associated species pairs were more than the negatively onepairs, and the ratio of positive to negative association of arbor layer and shrub layer were 1.8∶1 and 1.5∶1, respectively. The majority of species pairs were not significantly associated, indicating that the ecological characteristics of most dominant species in the P. sheareri community were relatively consistent. Conclusion The P. sheareri community was is relatively stable. However, appropriate artificial tending practices should be carried out to promote the natural regeneration of these P. sheareri populations. -
Key words:
- Phoebe sheareri
- / natural community
- / dominant species
- / niche
- / interspecific association
-
图 1 紫楠群落乔木层、灌木层优势物种种间Pianka生态位重叠指数
Figure 1. Pianka niche overlap index among dominant species in the arbor and shrub layer of P. sheareri community
图 2 紫楠群落乔木层、灌木层优势物种种间χ2检验半矩阵图
Figure 2. Semi-matrix diagram of χ2 test among dominant species in the arbor and shrub of P. sheareri community
表 1 紫楠天然群落乔木层主要物种重要值与生态位宽度
Table 1. Important value and niche breadth of dominant species in the arbor layer of P. sheareri community
序号
No.物种名
Species name缩写
Abbreviation重要值
IV/%生态位宽度 Niche breadth BL BS 1 紫楠 P. sheareri Ps 30.71 5.02 1.27 2 薄叶润楠 M. leptophylla Ml 18.02 4.08 1.06 3 柳杉 C. fortunei Cf 12.70 3.15 0.99 4 枫香 Liquidambar formosana Hance Lf 8.91 3.02 0.95 5 木荷 Schima superba Gardn. et Champ. Ss 7.16 2.04 0.84 6 杉木 Cunninghamia lanceolata (Lamb.) Hook. Cl 6.62 2.53 0.89 7 黄果朴 Celtis sinensis Pers. Cs 5.22 2.13 0.70 8 青冈栎 Quercus glauca Thunb. Cg 5.10 2.07 0.61 9 油桐 Vernicia fordii (Hemsl.) Airy Shaw Vf 4.22 2.25 0.59 10 棕榈 Trachycarpus fortunei (Hook.) H. Wendl. Tf 2.69 2.04 0.30  下载: 导出CSV
下载: 导出CSV
表 2 紫楠天然群落灌木层主要物种重要值与生态位宽度
Table 2. Important value and niche breadth of dominant species in the shrub layer of P. sheareri community
序号
No.物种名
Species name缩写
Abbreviation重要值
IV/%生态位宽度
Niche breadthBL BS 1 紫楠 P. sheareri Ps 14.69 4.17 1.24 2 薄叶润楠 M. leptophylla Ml 12.75 3.89 0.98 3 茶 C. sinensis Cs 11.77 3.14 0.96 4 阔叶箬竹 Indocalamus latifolius (Keng) McClure Il 9.29 3.05 0.89 5 毛柄连蕊茶 Camellia fraterna Hance Cf 5.73 2.74 0.86 6 高粱泡 Rubus lambertianus Ser. Rl 7.20 2.69 0.86 7 紫麻 Oreocnide frutescens (Thunb.) Miq. Of 7.86 2.56 0.81 8 朱砂根 Ardisia crenata Sims Ac 3.15 2.48 0.75 9 香花鸡血藤 Callerya dielsiana (Harms) P. K. Loc ex Z. Wei & Pedley Cd 4.76 2.71 0.79 10 杜茎山 Maesa japonica (Thunb.) Moritzi. ex Zoll. Mj 4.36 2.31 0.56
下载: 导出CSV
表 3 紫楠群落乔木层、灌木层优势树种总体联结性
Table 3. Overall interspecific associations among dominant species in the arbor layer and shrub layer of P. sheareri community
空间层次
Stand layer方差比率(VR)
Variance ratio检验统计量(W)
Statisticχ2临界值
χ2 threshold检验结果
Results乔木层 1.247 17.511 χ20.95(12)=5.226,χ20.05(12)=21.026 不显著正联结 灌木层 0.674 41.894 χ20.95(36)=23.269,χ20.05(36)=50.998 不显著负联结
下载: 导出CSV
-
[1] 陈爱莉, 赵志华, 龚 伟, 等. 气候变化背景下紫楠在中国的适宜分布区模拟[J]. 热带亚热带植物学报,2020,28(5):435-444. doi: 10.11926/jtsb.4201 [2] 李 军, 陆云峰, 杨安娜, 等. 紫楠天然群落物种多样性对不同干扰强度的响应[J]. 浙江农林大学学报,2019,36(2):279-288. doi: 10.11833/j.issn.2095-0756.2019.02.009 [3] 李 娟, 欧汉彪, 林建勇, 等. 我国楠属种质资源分布现状及主要种特征差异[J]. 广西林业科学,2020,49(1):54-59. doi: 10.3969/j.issn.1006-1126.2020.01.011 [4] 陈云霞, 谢春平. 华东地区楠木属植物的地理分布及资源保护对策[J]. 林业科技开发,2014,28(3):13-17. [5] WANG Y, MA X H, LU Y F, et al. Assessing the current genetic structure of 21 remnant populations and predicting the impacts of climate change on the geographic distribution of Phoebe sheareri in southern China[J]. Science of the Total Environment,2022,846: 157391. doi: 10.1016/j.scitotenv.2022.157391 [6] 陆云峰, 杨安娜, 张俊红, 等. 紫楠转录组EST-SSR标记开发及通用性分析[J]. 农业生物技术学报,2018,26(6):1014-1024. [7] VANDERMEER J. The niche construction paradigm in ecological time[J]. Ecological Modelling,2008,214(2): 385-390. [8] 杨宗慧, 郎学东, 李帅锋, 等. 篦子三尖杉群落优势种群生态位和种间关系[J]. 林业科学研究,2015,28(4):473-478. doi: 10.3969/j.issn.1001-1498.2015.04.004 [9] 赵 安, 周晓雷, 史瑞锦, 等. 青藏高原东北边缘云-冷杉林火烧迹地灌木群落种群生态位特征[J]. 林业科学研究,2022,35(6):108-117. [10] 白晓航, 张金屯. 小五台山森林群落优势种的生态位分析[J]. 应用生态学报,2017,28(12):3815-3826. [11] 王 霞, 铁 军, 刘泽宇, 等. 山西陵川南方红豆杉群落优势种生态位特征[J]. 生态科学,2017,36(5):64-72. [12] GUNTON R M, BOYES L J, GRIFFITHS M E, et al. Regeneration niches and functional traits of three common species in subtropical dune forest[J]. Forest Ecology & Management,2010,260(9): 1490-1497. [13] 赵 安, 田 青, 周晓雷, 等. 迭山北坡云冷杉林火烧迹地灌木树种种间关联性[J]. 林业科学研究,2022,35(2):163-170. [14] 姜宗庆, 李成忠, 周 霞, 等. 紫楠种子休眠特性及解除措施研究[J]. 种子,2020,39(12):92-94 + 98. [15] 柳 苗, 高捍东, 高 燕, 等. 休眠解除过程中紫楠种子生理生化特征的变化[J]. 南京林业大学学报(自然科学版),2023,47(2):9-17. [16] 陆云峰, 王 豪, 徐沁怡, 等. 樟科楠属4种植物种实表型性状及其发芽特性研究[J]. 种子,2020,39(12):107-112 + 119. [17] 陈模芳, 韦小丽, 张 怡. 紫楠实生幼苗的光合生理特性[J]. 贵州农业科学,2013,41(10):55-58. doi: 10.3969/j.issn.1001-3601.2013.10.015 [18] 涂 铭, 李玮婷, 费 越, 等. 5种楠属植物叶形态与高光谱特征比较[J]. 西南林业大学学报(自然科学),2017,37(6):113-118. [19] 周存宇, 万小丽, 张 建, 等. 五种楠属植物叶片油细胞和黏液细胞的比较[J]. 湖北农业科学,2015,54(18):4506-4508 + 4518. [20] 邓送求, 闫家锋, 关庆伟. 宝华山紫楠风景林林分空间结构分析[J]. 东北林业大学学报,2010,38(4):29-32. doi: 10.3969/j.issn.1000-5382.2010.04.010 [21] 范 忆, 楼一恺, 库伟鹏, 等. 天目山紫楠种群年龄结构与点格局分析[J]. 浙江农林大学学报,2020,37(6):1027-1035. doi: 10.11833/j.issn.2095-0756.20190631 [22] 方精云, 王襄平, 沈泽昊, 等. 植物群落清查的主要内容、方法和技术规范[J]. 生物多样性,2009,17(6):533-548. [23] 郝建锋, 李 艳, 齐锦秋, 等. 人为干扰对碧峰峡栲树次生林群落物种多样性及其优势种群生态位的影响[J]. 生态学报,2016,36(23):7678-7688. [24] BHUYAN P, KHAN M L, TRIPATHI R S. Tree diversity and population structure in undisturbed and human-impacted stands of tropical wet evergreen forest in Arunachal Pradesh, Eastern Himalayas, India[J]. Biodiversity & Conservation,2003,12(8): 1753-1773. [25] LEVINS R. Evolution in Changing Environments: Some Theoretical Explorations[M]. Princetion: Princetion University Press, 1968. [26] COLWELL R K, FUTUYMA D J. On the measurement of niche breadth and overlap[J]. Ecology,1971,52(4): 567-576. doi: 10.2307/1934144 [27] PIANKA E R. The structure of lizard communities[J]. Annual Review of Ecology and Systematics,1973,4: 53-74. doi: 10.1146/annurev.es.04.110173.000413 [28] SCHLUTER D. A variance test for detecting species associations with some example applications[J]. Ecology,1984,65(3): 998-1005. doi: 10.2307/1938071 [29] GREIG-SMITH P. Quantitative Plant Ecology[M]. London: Butter-worths, 1957: 1-359. [30] PETRAITIS P S. Algebraic and Graphical Relationships Among Niche Breadth Measures[J]. Ecology,1981,62(3): 545-548. doi: 10.2307/1937719 [31] 胡正华, 钱海源, 于明坚. 古田山国家级自然保护区甜槠林优势种群生态位[J]. 生态学报,2009,29(7):3670-3677. doi: 10.3321/j.issn:1000-0933.2009.07.027 [32] 朱德煌, 刘金福, 洪 伟, 等. 戴云山黄山松群落主要树种更新生态位研究[J]. 热带亚热带植物学报,2012,20(6):561-565. doi: 10.3969/j.issn.1005-3395.2012.06.004 [33] 徐远杰, 林敦梅, 石 明, 等. 云南哀牢山常绿阔叶林的空间分异及其影响因素[J]. 生物多样性,2017,25(1):23-33. [34] 高浩杰, 袁 佳, 高平仕. 舟山群岛次生林优势种群的生态位与种间联结[J]. 热带亚热带植物学报,2017,25(1):73-80. doi: 10.11926/jtsb.3620 [35] 马晓迪, 姜德刚, 刘子琳, 等. 平潭岛台湾相思群落优势种群生态位研究[J]. 热带作物学报,2022,42(12):1-13. doi: 10.3969/j.issn.1000-2561.2022.12.024 [36] 刘益鹏, 叶兴状, 叶利奇, 等. 观光木群落优势树种生态位和种间联结[J]. 应用生态学报,2022,33(10):2670-2678. [37] RAJIV P, MONIKA R, VISHAL S, et al. Large scale field-based evaluation of niche breadth, niche overlap and interspecific association of Western Himalayan temperate forest tree species[J]. Ecological Indicators,2023,146: 109876. doi: 10.1016/j.ecolind.2023.109876 [38] 王祥福, 郭泉水, 巴哈尔古丽, 等. 崖柏群落优势乔木种群生态位[J]. 林业科学,2008,44(4):6-13. doi: 10.3321/j.issn:1001-7488.2008.04.004 [39] 刘金福, 洪 伟. 格氏栲群落生态学研究——格氏栲林主要种群生态位的研究[J]. 生态学报,1999,19(3):347-352. doi: 10.3321/j.issn:1000-0933.1999.03.010 [40] 臧 敏, 邱筱兰, 李晓艳. 江西三清山浙江楠群落主要种群生态位研究[J]. 亚热带植物科学,2016,45(1):63-66. [41] 李坚锋, 潘 萍, 欧阳勋志, 等. 庐山常绿阔叶林种间联结性及物种共存机制[J]. 生态学杂志,2022,41(8):1474-1481. [42] 兰航宇, 段文标, 陈立新, 等. 小兴安岭天然针阔混交林主要树种空间格局及其关联性[J]. 生态学报,2019,39(18):6660-6669. [43] 王亚飞, 曲美学, 兰航宇, 等. 云冷杉红松林内优势种群的空间格局及种间关联性[J]. 中南林业科技大学学报,2020,40(1):49-58. [44] 何 江, 徐来仙, 艾训儒. 湖北七姊妹山亚热带常绿落叶阔叶混交林主要木本植物的生态位与种间联结[J]. 林业科学研究,2023,36(3):138-148. [45] 李 娟, 温远光, 王 成, 等. 南宁市城区森林灌木层优势种种间联结分析[J]. 林业科学研究,2009,22(2):230-236. doi: 10.3321/j.issn:1001-1498.2009.02.014 [46] 张明霞, 王得祥, 康 冰, 等. 秦岭华山松天然次生林优势种群的种间联结性[J]. 林业科学,2015,51(1):12-21. doi: 10.11707/j.1001-7488.20150102 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 309
- HTML全文浏览量: 84
- PDF下载量: 1
- 被引次数: 0
