

-
氮(N)的可用性深切影响着生态系统的生产力和地球化学过程。土壤中超过90%的N以有机态形式存在,并不具有直接的生物可利用性,因此氮矿化速率不仅是衡量土壤氮库周转速率的核心指标,同时决定了土壤的供氮能力[1]。团聚体是土壤的基本单元结构,它的形成包含了复杂的生化和物理过程,土壤氮矿化的实质是各个粒径的团聚体氮矿化的整合结果。排除机械外力因素,不同粒径的团聚体被认为是由不同种类的黏合剂胶结而成,因此它们的稳定性并不相同[2]。有报道指出,较小粒径的团聚体具有更大的比表面积,因此可以吸附更多的可矿化有机物质,而较大粒径的团聚体中的有机物质由于空间和氧气扩散有限等原因,与微生物和胞外酶接触的概率会下降,因此较小粒径的团聚体将具有更高的氮矿化速率[3];但也有观点认为相比于小粒径的团聚体,新鲜有机质会首先进入大粒径团聚体,因此大粒径团聚体中的活性有机质含量更高,也将更先被利用和矿化[4]。总体来说,对于团聚体粒径与氮矿化的关系尚有待讨论。
土壤氮矿化同时受到众多因子的调控,其中土壤水分和温度在众多非生物因素中起到主导作用[5]。温度作为影响土壤氮矿化的主控因子,一般认为,温度增加会促进土壤氮矿化,如Katrin等[6]报道土壤净氨化和净硝化与温度成正比。另外,一项 Meta分析显示全球变暖导致净氮硝化和净氮矿化分别增加了52%和32%[7]。但一项在长白山开展的研究中发现,氮矿化与温度没有显著关系[8]。周于波等[9]通过培养研究发现,土壤净氮矿化速率在5 ~26.5 ℃范围内随温度升高而增加,之后随温度升高而显著降低。土壤水分解释了在全球尺度上17%的氮矿化变异,且对森林生态系统的贡献高于农业生态系统和草原生态系统[10]。传统观点认为水分的增加会提高氮矿化速率,如Stanford等[11]、Knoepp等[12]的研究发现,氮矿化速率在土壤水分接近田间持水量时达到最高。但是在该领域的研究结果并不统一,如Guntiñas等[13]报道,氮矿化在土壤含水量为60%~70%田间持水量时达到最高。众多学者证实土壤水分和温度对土壤氮矿化速率的影响存在显著交互作用,如Jennife[14]在针阔混交林的研究中发现,土壤净硝化和净氮矿化速率均显示出随土壤含水量和温度升高而增加的趋势,温度和水分共同解释了83%的土壤氮矿化变异。因此,厘清温度和水分对森林土壤氮矿化的影响及其交互效应,对提高生态系统生产力具有指导意义。
CO2等温室气体排放增加导致的气温上升已经成为全球生态系统正在面临的挑战。林地碳汇是中国碳汇绝对主体,在总量中占比约为80.7%,发展和保护森林生产力已经成为减缓气候变化的重要举措之一[15]。《中国气候变化蓝皮书(2022)》指出[16],我国近几十年来的气温上升速率显著高于同期全球平均水平,其中中部地区升温速率为0.23 ℃·(10 a)−1,同时伴有1.24 mm·(10 a)−1 的降水量增加速率[17]。三峡库区地处北亚热带,夏季高温多雨,是长江中下游重要的生态屏障,但同时也是我国典型的生态系统稳定性较为脆弱的地区[18]。但目前针对该地区森林生态系统在全球气温升高和降水增多背景下的土壤氮矿化特征鲜有报道。本研究以三峡库区马尾松-栓皮栎针阔混交林为研究对象,研究土壤水分、温度及二者的交互作用对土壤净氮转化作用的影响,以期为预测该地区温室效应和降水增多背景下的土壤养分循环提供数据支撑和为当地人工林生产力的提升提供理论依据。
-
样地位于湖北省秭归县茅坪镇泗溪村(30′46′ N,110°56′ E,海拔 825 m),属亚热带季风气候,年平均气温14 ~22 ℃,年平均降水量1400 mm。研究区森林类型为针阔混交林,优势种为马尾松和栓皮栎,为20世纪80年代飞播造林,平均林龄40 a,平均树高和胸径分别为16.1 m和18.3 cm,样地内林木分布均匀。地带性土壤为黄壤,深度约40 cm。林下植物主要有茶(Camellia sinensis (L.) Kuntze)、细齿叶柃(Eurya nitida Korth.)、宜昌荚蒾(Viburnum erosum Thunb.)、黑足鳞毛蕨(Dryopteris fuscipes)、鱼腥草(Houttuynia cordata)等。
-
2018年8月,在马尾松-栓皮栎针阔混交人工林内设置3个20 m×20 m的样方。2021年8月,在样方中去除地表凋落物,并随机取50个采样点,用直径5 cm,高10 cm的PVC管取样方中0~20 cm的原状土,用保鲜膜封存,置于冷藏箱中运回实验室4 ℃冰箱保存。在实验室将PVC管中的土样小心取出,将来自同个样方采集的土样混合,顺着缝隙将土壤掰成8 000 μm以下的小块。根据Bach[19]出的筛分法将土壤进行粒径分级,得到大团聚体(2 000~8 000 μm)、中团聚体(1 000~2 000 μm)、小团聚体(250~1 000 μm)和微团聚体(<250 μm)。分级后的团聚体样品分成两部分,分别用于土壤 pH等土壤性质检测以及矿化试验。
-
将部分团聚体自然风干后分别过2 mm和0.149 mm的筛子,用于土壤基本性质的检测。使用电极法测定土壤 pH (水土比为2.5∶1.0,pH211 酸度计);高温外热重铬酸钾氧化-容重法测定土壤有机碳,凯氏定氮法测定全氮,盐酸、氟化铵浸提-钼锑抗比色法测定速效磷,乙酸铵浸提-火焰光度法测定速效钾。土壤氨态氮和硝态氮含量使用氯化钾浸提法和流动分析仪完成检测。
-
设置3个处理:0.4 FCW (40% 田间最大持水量)、0.6 FCW (60% 田间最大持水量)和0.8 FCW (80% 田间最大持水量),使用纯水完成水分调整,将样品置于25 ℃的培养箱中预培养7 d以恢复微生物活性。称取相当于20 g干土质量的团聚体置于350 mL梅森瓶中,用无菌透气封口膜封住瓶口,分别置于5、10、25和35 ℃的培养箱中培养28 d,期间每24 h采用称重法补充团聚体水分。培养结束后的样品用于测定矿化实验结束后的氨态氮和硝态氮含量测定。
土壤净氮矿化速率的计算[8]:
$R_{\min } \mathrm{NH}_4^{+}{\text{-}}\mathrm{N}=\left(N_{\mathrm{B}} \mathrm{NH}_4^{+}{\text{-}}\mathrm{N}-N_{\mathrm{A}} \mathrm{NH}_4^{+}{\text{-}}\mathrm{N}\right) / \mathrm{d} $
$R_{\min } \mathrm{NO}_3^{-}{\text{-}}\mathrm{N}=\left(N_{\mathrm{B}} \mathrm{NO}_3^{-}{\text{-}}\mathrm{N}-N_{\mathrm{A}} \mathrm{NO}_3^{-}{\text{-}}\mathrm{N}\right) / \mathrm{d} $
$R_{\min } \mathrm{N}=R_{\min } \mathrm{NH}_4^{+}{\text{-}}\mathrm{N}+R_{\min } \mathrm{NO}_3^{-}{\text{-}}\mathrm{N}$
式中:
$R_{\min } \mathrm{NH}_4^{+}{\text{-}}\mathrm{N} $ 为净氨化速率;$R_{\min } \mathrm{NO}_3^{-}{\text{-}}\mathrm{N} $ 为净硝化速率;$N_{\mathrm{B}} \mathrm{NH}_4^{+}{\text{-}}\mathrm{N} $ 为培养后氨态氮量;$N_{\mathrm{B}} \mathrm{NO}_3^{-}{\text{-}}\mathrm{N} $ 为培养后硝态氮量;$N_{\mathrm{A}} \mathrm{NH}_4^{+}{\text{-}}\mathrm{N} $ 为培养前氨态氮量;$N_{\mathrm{A}} \mathrm{NO}_3^{-}{\text{-}}\mathrm{N} $ 为培养前硝态氮量;RminN为净氮矿化速率;d 为培养时间(28 d)。每个处理重复3次。
-
用 Excel 2016 和 SPSS 22.0 进行试验数据统计分析,采用单因素方差分析(Oneway ANOVA)和 Duncan 多重比较对团聚体在不同培养条件下各指标进行显著性检验,检验水平 P= 0.05。 用双因素方差分析( two-way ANOVA) 研究水分和温度及其交互作用对团聚体净氮矿化的影响。图表中数据为“平均值 ± 标准差”(n=3)。使用Origin 2021绘图。
-
马尾松-栓皮栎针阔混交林土壤不同粒径团聚体的理化性质和质量比例如表1所示。土壤pH和速效钾含量随团聚体粒径变化没有出现显著差变化。土壤全氮、有机质含量以及微生物生物量碳、氮均在大团聚体中最低,在微团聚体中最高;而土壤速效磷含量随粒径变化规律与之相反。团聚体的重量比例随粒径减小呈现倒“V”变化,从高到低为小团聚体>中团聚体>大团聚体>微团聚体。
表 1 不同粒径团聚体理化特征
Table 1. Physicochemical characteristics of aggregates with different particle sizes
大团聚体
Large macroaggregates中团聚体
Coarse aggregate小团聚体
Small macroaggregates微团聚体
Microaggregate全氮
Total N/ (g·kg−1)1.41 ± 0.14 b 2.55 ± 0.26 a 2.76 ± 0.28 a 2.81 ± 0.57 a 土壤有机质
Soil organic matter/(g·kg−1)17.79 ± 2.61 c 24.85 ± 3.12 b 29 ± 5.01 ab 32.13 ± 3.38 a 速效磷
Available phosphorus/(mg·kg−1)13.11 ± 0.55 a 13.61 ± 0.48 a 13.35 ± 0.33 ab 11.81 ± 1.15 b 速效钾
Available potassium/(mg·kg−1)103.47 ± 1.21 a 102.7 ± 1.35 a 102.3 ± 2.15 a 103 ± 1.18 a 土壤酸碱度
pH (H2O)5.35 ± 0.26 a 5.02 ± 0.08 a 5.12 ± 0.1 a 5.28 ± 0.34 a 微生物量碳
Microbial biomass C/(mg·kg−1)244.69 ± 41.15 c 252.76 ± 15.97 bc 321.72 ± 62.34 ab 376.61 ± 7.15 a 微生物量氮
Microbial biomass N/(mg·kg−1)40.49 ± 0.16 b 44.64 ± 4.34 b 56.97 ± 5.65 a 64.38 ± 6.47 a 质量比例
Weight proportions
of aggregates in soil0.21 ± 0.04 c 0.29 ± 0.29 b 0.39 ± 0.04 a 0.11 ± 0.01 d 注:不同小写字母标注代表在0.05检验水平上差异显著,下同
Notes: Different small letters indicate significant difference at 0.05 test level, the same below -
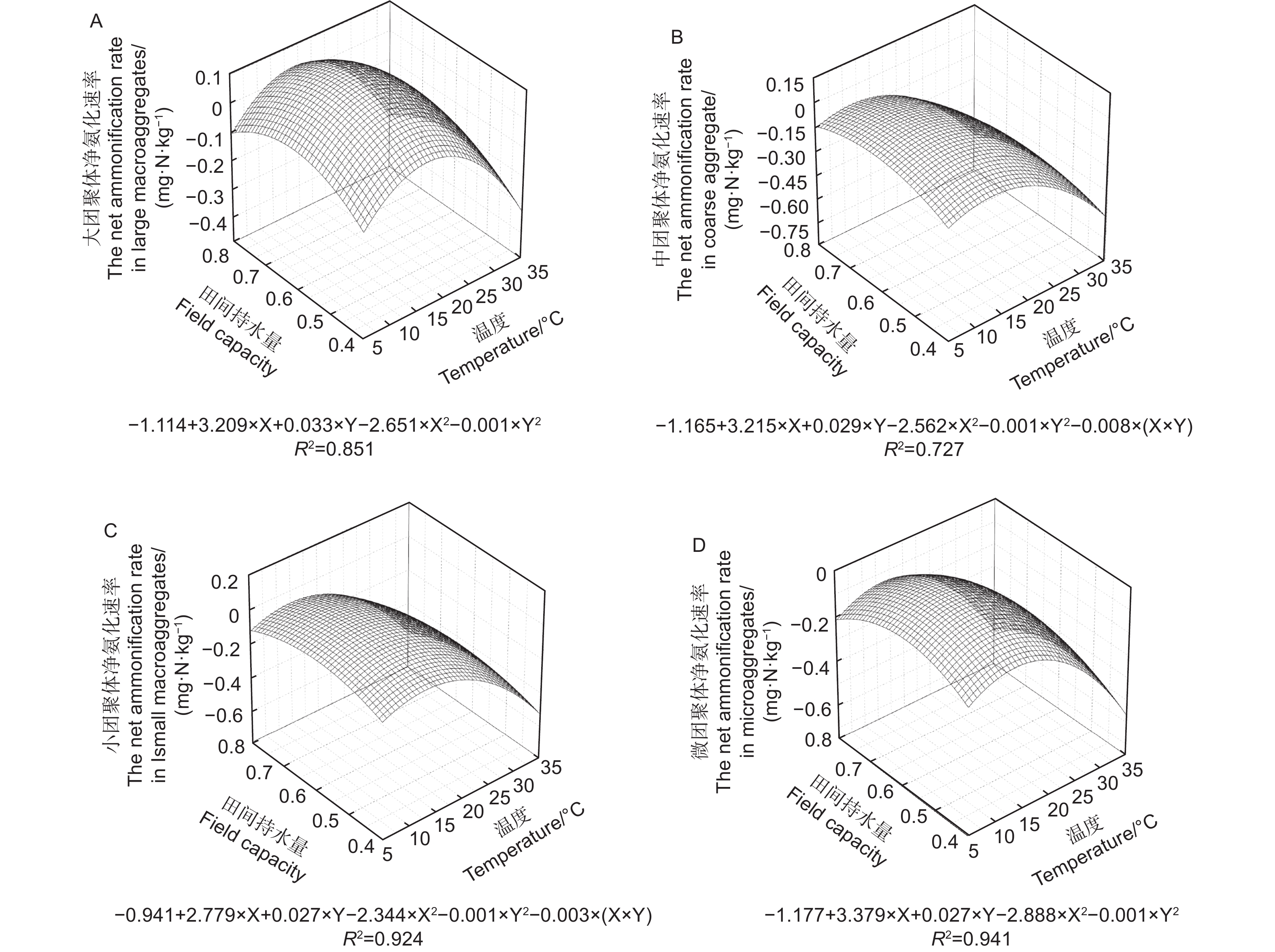
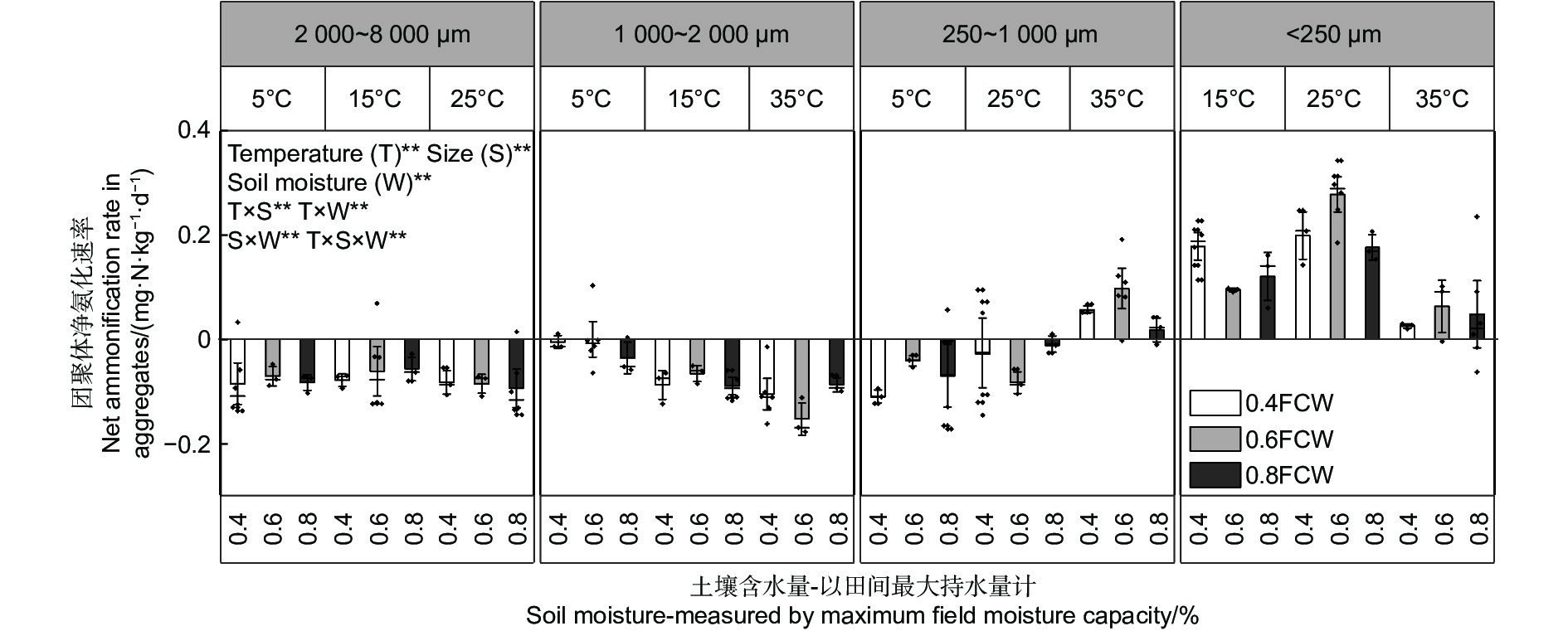
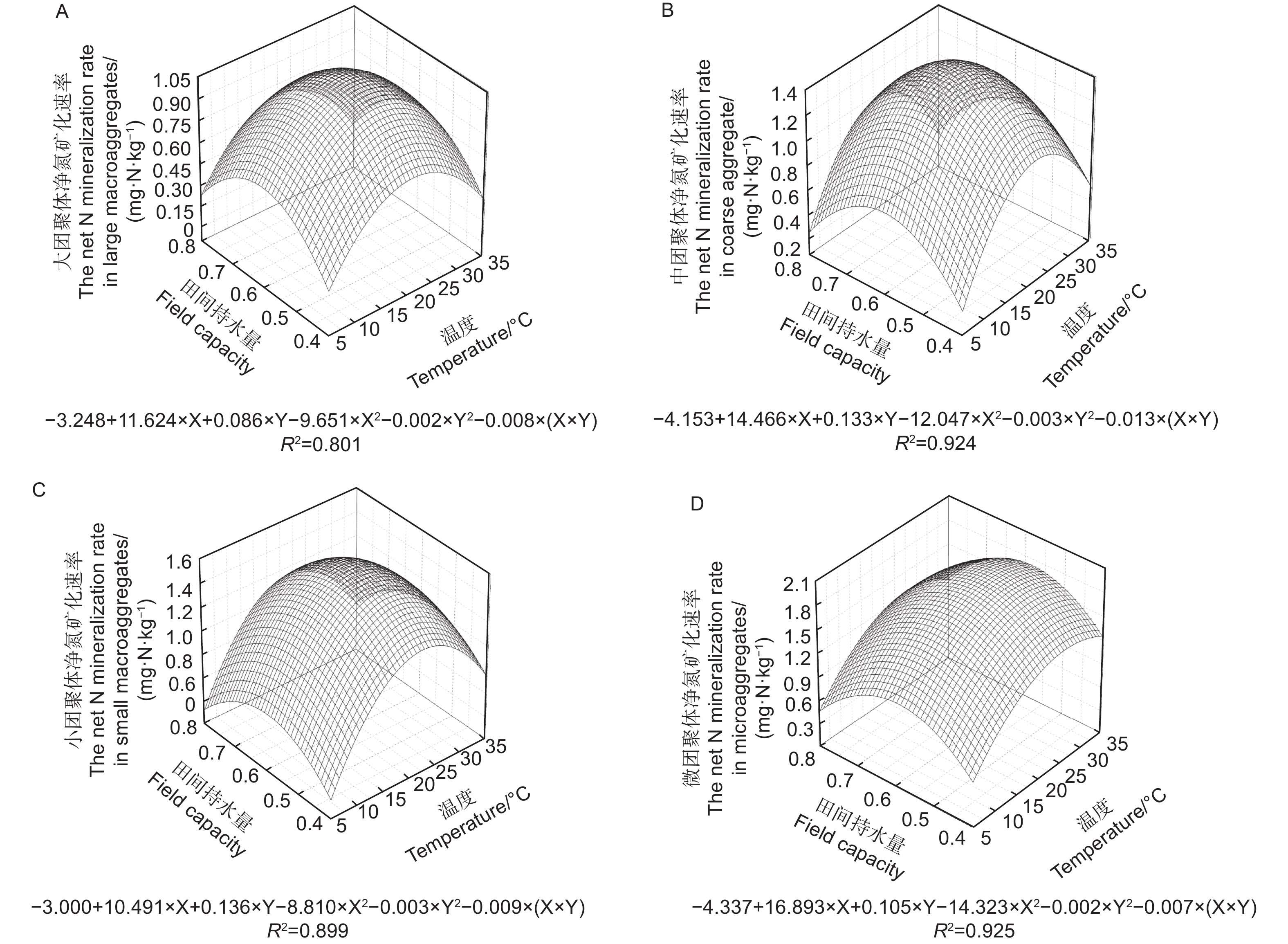
不同粒径团聚体净氨化速率( RminNH4+-N)随水热变化的响应如图1所示,温度、土壤含水量以及粒径均对团聚体净氨化速率产生了显著影响。整体分析,微团聚体的净氨化速率整体最高。当土壤含水量为0.6 FCW、培养温度为25 ℃时,土壤团聚体净氨化速率均>0 mg N kg−1·d−1,说明在该条件下,4个粒径的团聚体中均净增了NH4+-N。由图2A~D可知,4个粒径的团聚体净氨化速率随水热条件变化响应拟合出的二元二次方程并不相同。由图2可估算出大团聚体、中团聚体、小团聚体和微团聚体的净氨化速率最大值分别出现在含水量为0.63 FCW、0.68 FCW、0.70 FCW和0.75 FCW;温度为22.3、20.5、23.0和23.5 ℃。
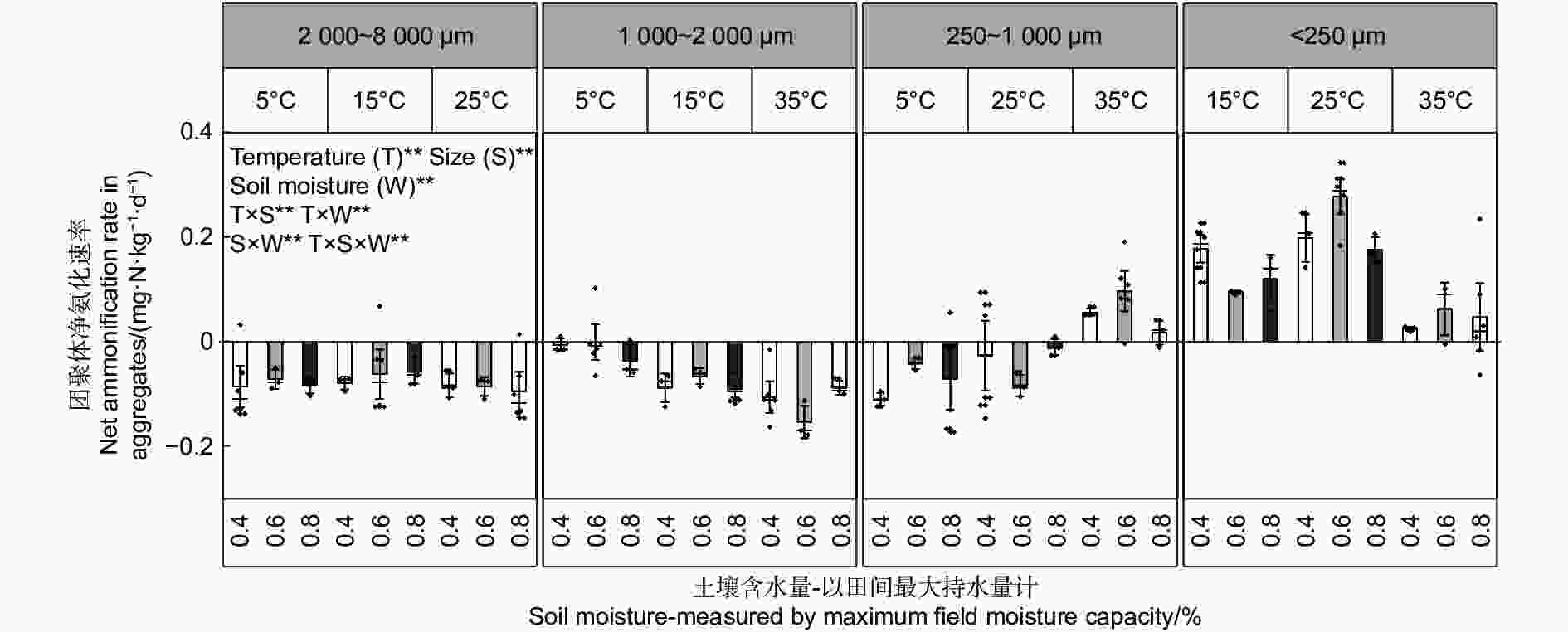
图 1 团聚体在不同水热条件下净氨化速率特征
Figure 1. Characteristics of net ammonification rate of agglomerates under different soil moisture and temperature conditions.
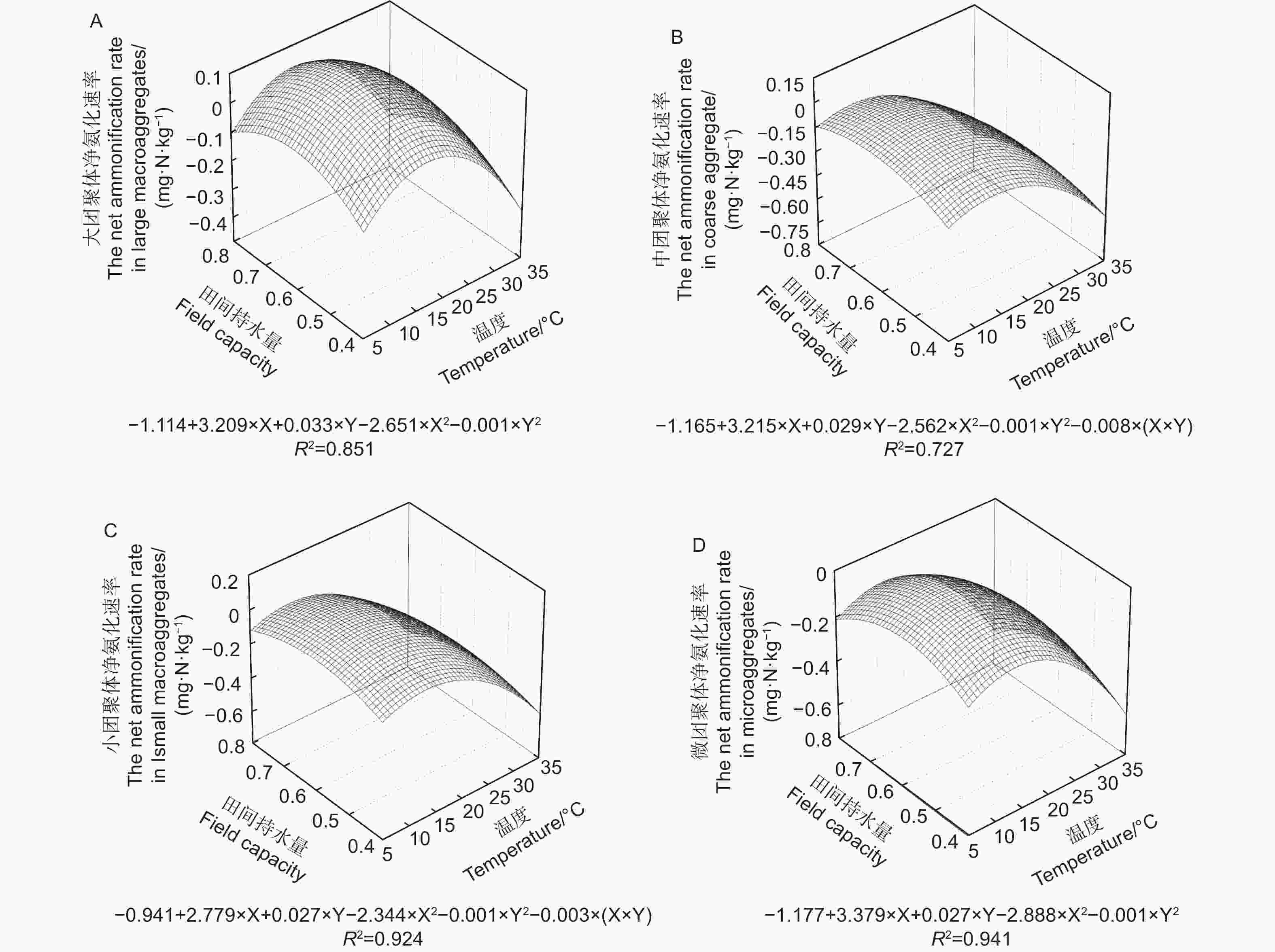
图 2 团聚体净氨化速率对土壤水分和温度变化响应模式
Figure 2. Response pattern of net ammonification rate of aggregates to changes in soil moisture and temperature
-
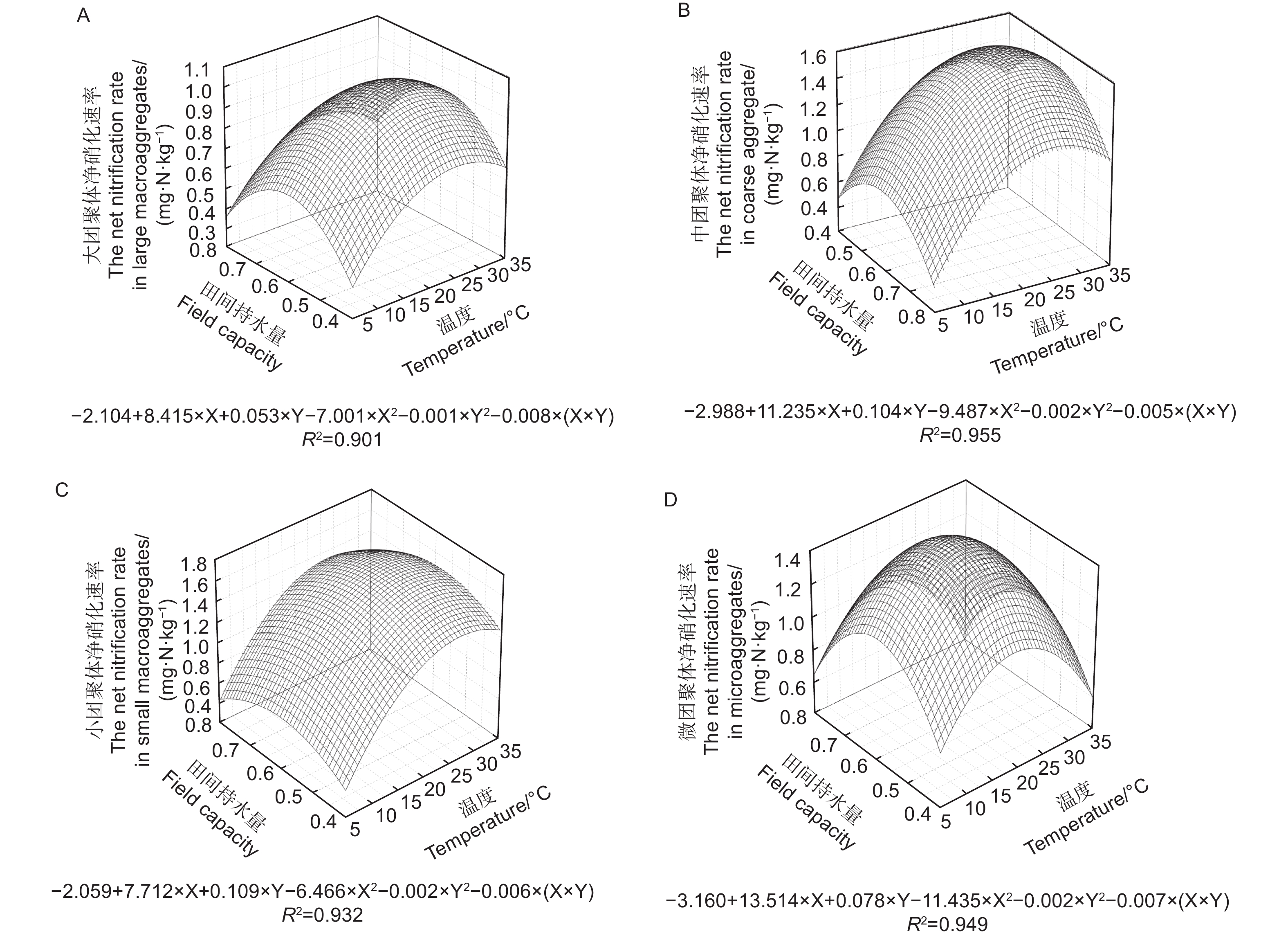
不同粒径团聚体净硝化速率( RminNO3−-N)在12种培养条件下随水热变化的响应如图3与净氨化速率相似,温度、土壤含水量以及粒径均对团聚体净硝化速率产生了显著影响( p<0.01)。整体分析,当土壤含水量为0.6 FCW、培养温度为25 ℃时,土壤团聚体净硝化速率最高。随团聚体粒径减小,净硝化速率存在上升趋势。由图4A~D可估算出大团聚体、中团聚体、小团聚体和微团聚体的净硝化速率最大值分别出现在含水量为0.58 FCW、0.57 FCW、0.60 FCW和0.61 FCW以及温度为24.9、25.5、27.5和25.7 ℃。
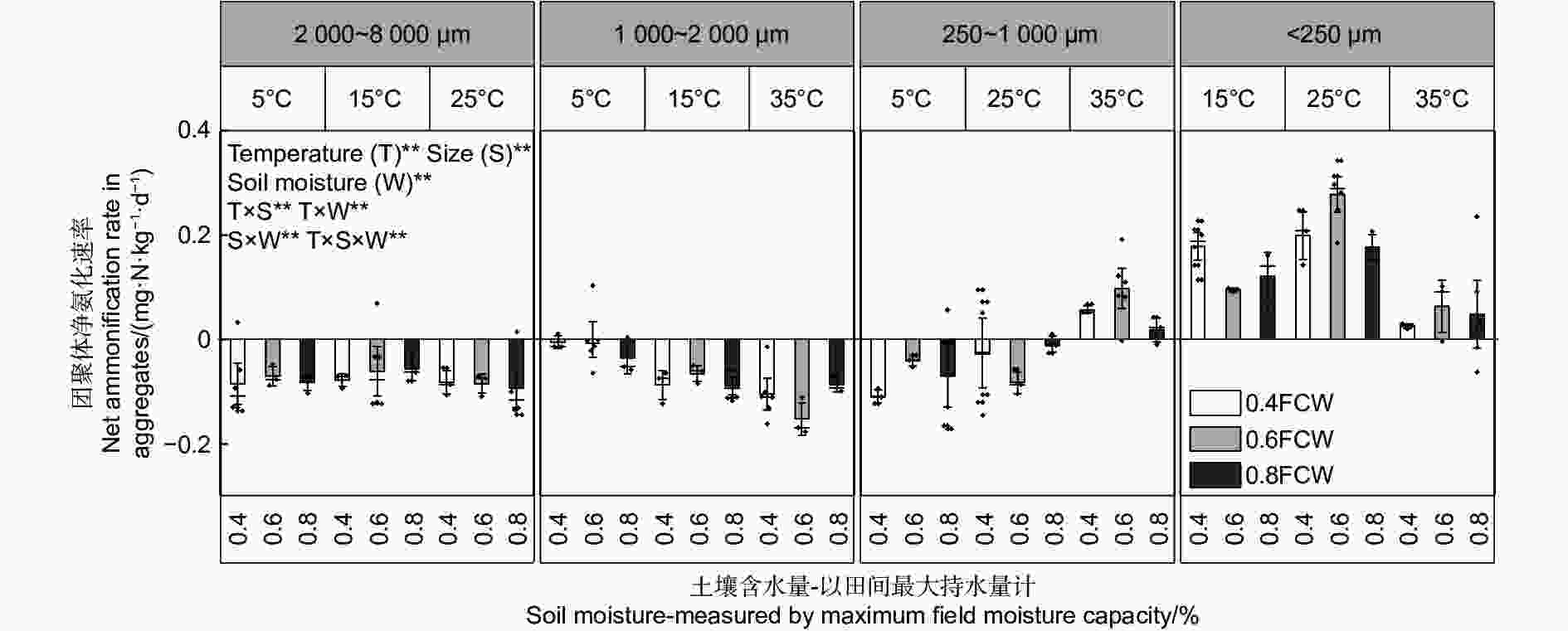
图 3 团聚体在不同水热条件下净硝化速率特征
Figure 3. Characteristics of net nitrification rate of aggregates under different soil moisture and temperature conditions
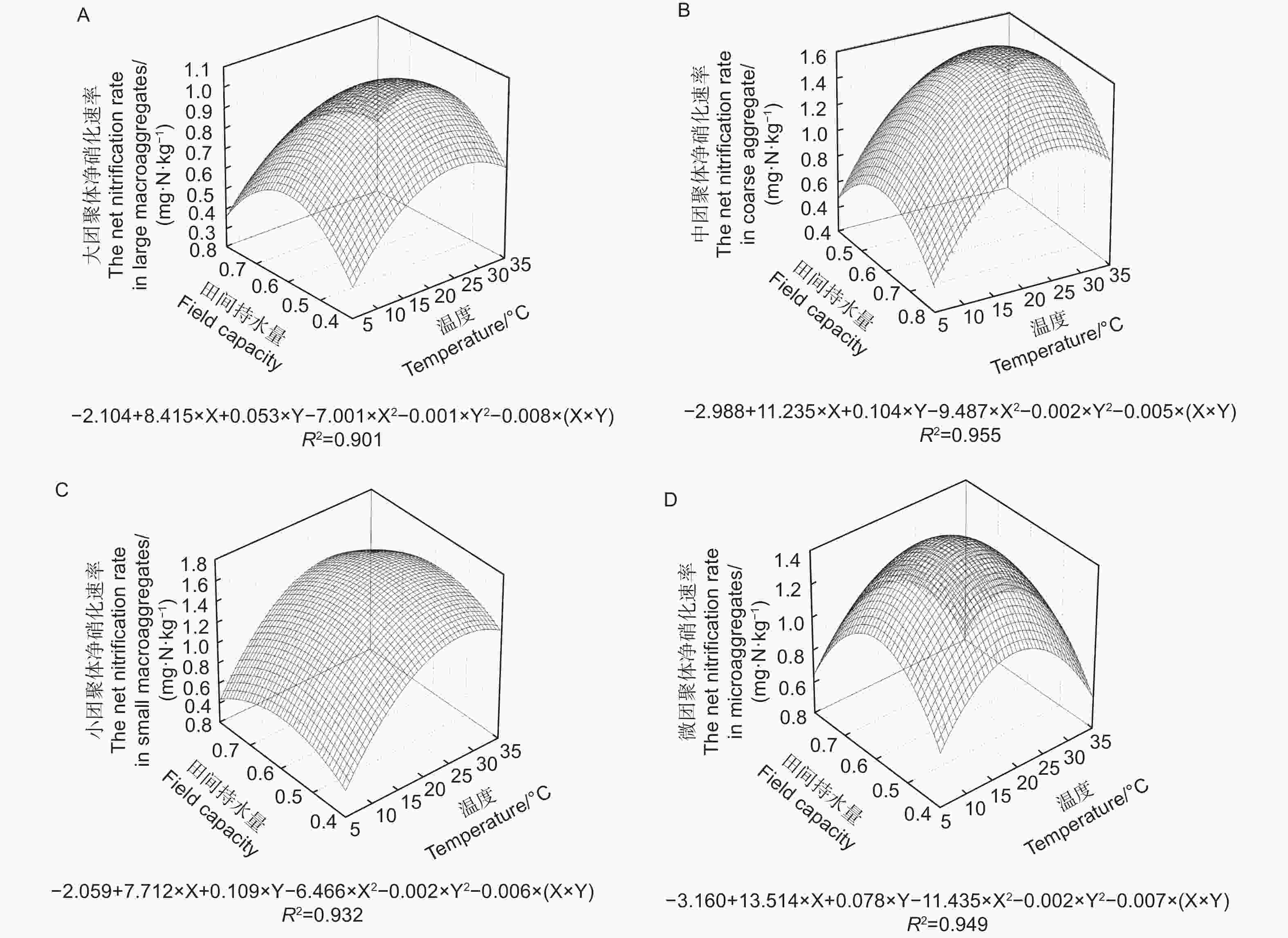
图 4 团聚体净氨化速率对土壤水分和温度变化响应模式
Figure 4. Response pattern of net nitrification rate of aggregates to changes in soil moisture and temperature
-
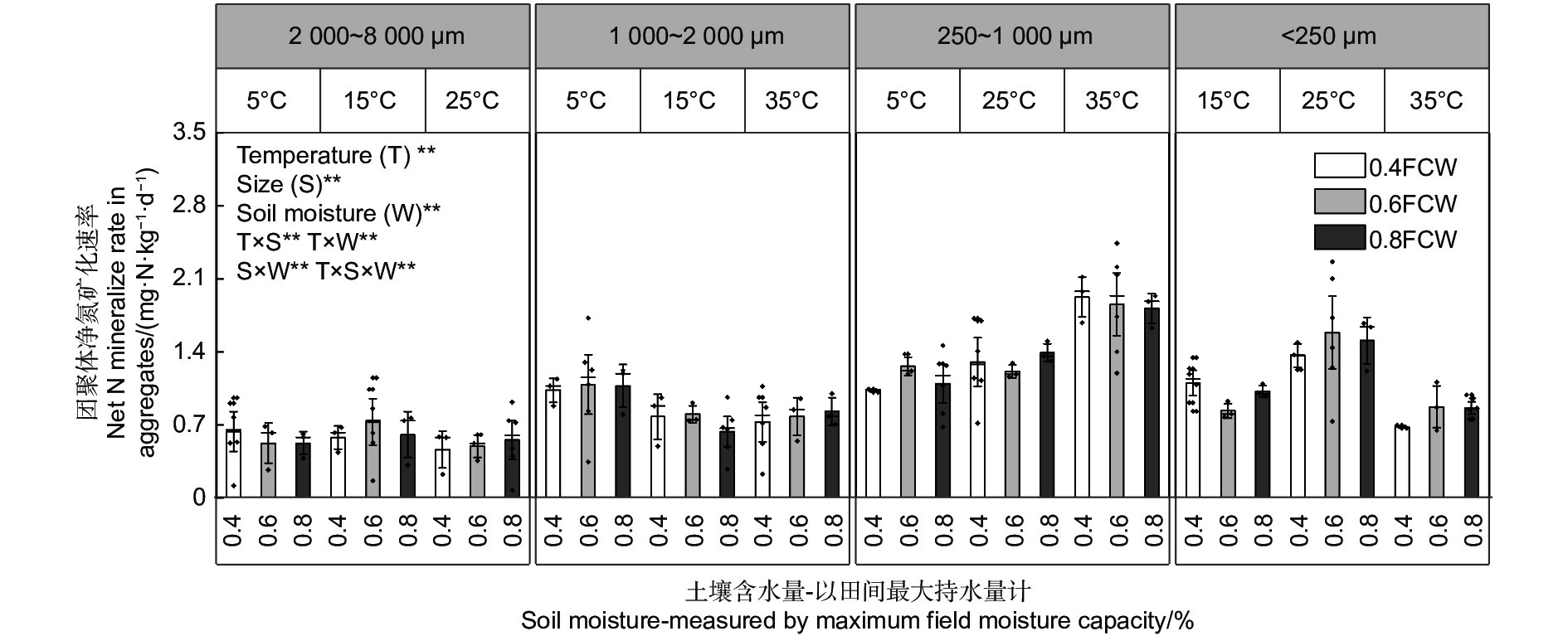
如图5所示,不同粒径团聚体净氮矿化速率(RminN)是净氨化速率与净硝化速率的之和,4种粒径的团聚体在12种水热培养条件下净硝化速率对净氮矿化速率的贡献均超过85%。温度、土壤含水量以及粒径均对团聚体净氮矿化速率产生了显著影响( p <0.01)。当土壤含水量为0.6 FCW、培养温度为25 ℃时,土壤团聚体净氮矿化速率最高。与净硝化速率相似,净氮矿化速率存在随团聚体粒径减小而上升的趋势。由图6A~D可估算出大团聚体、中团聚体、小团聚体和微团聚体的净氮矿化速率最大值分别出现在含水量为0.61 FCW、0.57 FCW、0.58 FCW和0.62 FCW以及温度为23.9、22.7、24.5和26.7 ℃。
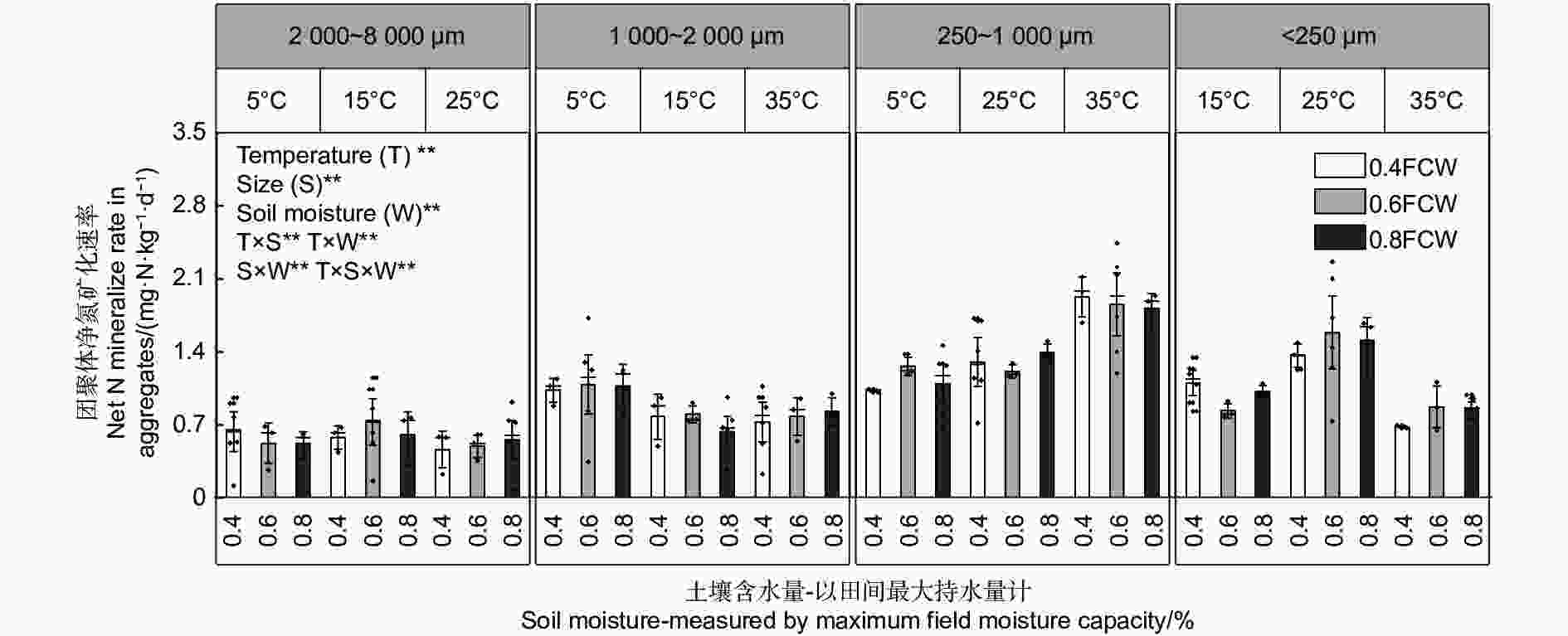
图 5 团聚体在不同水热条件下净氮矿化速率特征
Figure 5. Characteristics of net N mineralization rate of aggregatess under different soil moisture and temperature conditions
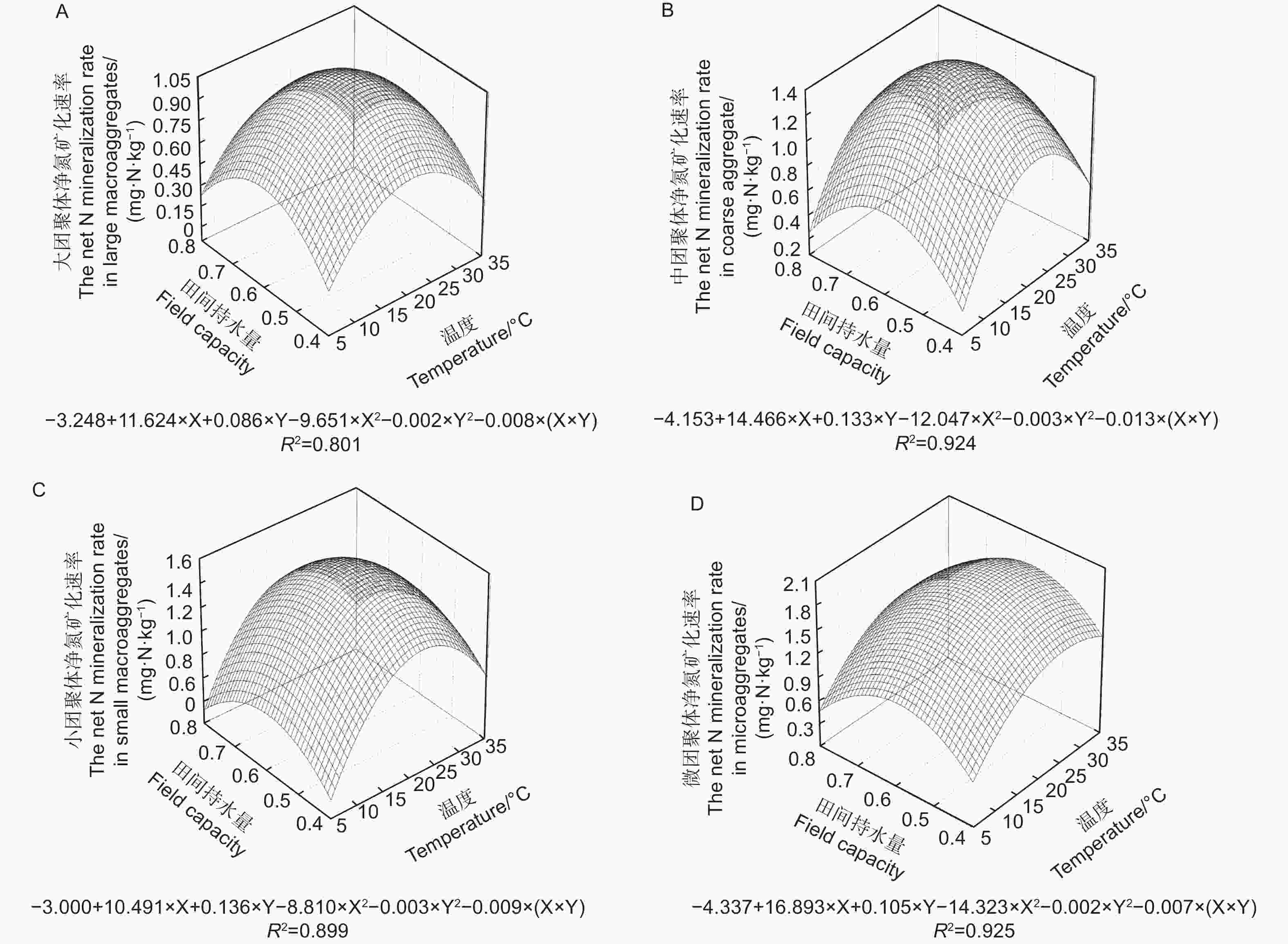
图 6 团聚体净氨化速率对土壤水分和温度变化响应模式
Figure 6. Response pattern of net N mineralization rate of aggregates to changes in soil moisture and temperature
-
4种粒径团聚体的净氨化、净硝化和净氮矿化速率的 Q10随水分变化数值如表2所示。大团聚体在各土壤水分处理下净氨化、净硝化和净氮矿化速率的Q10分别在−1.39~2.75、−4.84~1.96和−0.83~1.84之间;中团聚体在各土壤水分处理下净氨化、净硝化和净氮矿化速率的Q10分别在0.01~5.65、−4.32~1.92和−0.36~1.94之间;小团聚体在各土壤水分处理下净氨化、净硝化和净氮矿化速率的Q10分别在−1.22~2.49、−0.39~3.19和0.10~4.39之间;微团聚体在各土壤水分处理下的净氨化、净硝化和净氮矿化速率的Q10分别在−0.92~2.02、−2.11~9.46和−0.84~1.74之间,并整体在0.6 FWC、15 ℃/25 ℃时温度敏感性最大。
表 2 团聚体净氨化、净硝化和净氮矿化速率Q10在不同土壤水分下的变化
Table 2. Fluctuate in Q10 values in aggregates of net ammonification, net nitrification, and net nitrogen mineralization rates under different soil moisture treatments
0.4 FCW 0.6 FCW 0.8 FCW 5 ℃/15 ℃ 15 ℃/25 ℃ 25 ℃/35 ℃ 5 ℃/15 ℃ 15 ℃/25 ℃ 25 ℃/35 ℃ 5 ℃/15 ℃ 15 ℃/25 ℃ 25 ℃/35 ℃ 净氨化速率
Net
ammonification
rate大团聚体
(2 000~8 000 μm)0.69 ± 0.14 b 2.75 ± 0.93 b −1.39 ± 0.32 b 1.25 ± 0.28 c 1.26 ± 0.28 c 1.25 ± 0.26 c 1.36 ± 0.30 b 1.64 ± 0.29 b 0.83 ± 0.15 cd 中团聚体
(1 000~2 000 μm)0.31 ± 0.07 b 5.65 ± 11.49 a 0.01 ± 0.03 b 1.47 ± 0.24 bc 1.89 ± 0.31 ab 0.76 ± 0.05 de 1.55 ± 0.24 b 2.24 ± 0.36 a 0.65 ± 0.05 d 小团聚体
(250~1 000 μm)1.08 ± 0.11 b 1.97 ± 0.13 b −1.22 ± 0.20 b 1.17 ± 0.08 cd 1.92 ± 0.36 a 0.70 ± 0.11 e 1.18 ± 0.08 bc 2.49 ± 0.52 a 0.48 ± 0.09 d 微团聚体
(<250 μm)0.49 ± 0.2 bc 1.41 ± 0.49 a −0.92 ± 0.05 e 1.58 ± 0.31 a 1.43 ± 0.29 a 0.57 ± 0.08 b 2.02 ± 0.59 a 1.68 ± 0.42 a 0.47 ± 0.07 b 净硝化速率
Net
nitrification
rate大团聚体
(2 000~8 000 μm)−0.62 ± 0.15 de −4.84 ± 1.13 f 0.09 ± 0.00 cd 1.71 ± 0.39 a 1.66 ± 0.21 a 0.74 ± 0.08 b 1.96 ± 0.49 a 1.76 ± 0.23 a 0.68 ± 0.07 b 中团聚体
(1 000~2 000 μm)0.93 ± 0.17 ab 0.33 ± 0.14 bc −4.32 ± 0.24 f 1.69 ± 0.44 a 1.35 ± 0.25 a 0.57 ± 0.06 b 1.92 ± 0.71 a 1.55 ± 0.37 a 0.47 ± 0.07 b 小团聚体
(250~1 000 μm)0.06 ± 0.01 c 3.19 ± 6.10 a −0.39 ± 0.02 c 1.52 ± 0.27 a 1.56 ± 0.25 a 0.64 ± 0.04 b 1.76 ± 0.33 a 1.74 ± 0.28 a 0.54 ± 0.03 b 微团聚体
(<250 μm)9.46 ± 3.71 b −2.11 ± 0.67 c 0.11 ± 0.02 c 1.58 ± 0.06 a 1.81 ± 0.06 a 0.62 ± 0.04 b 1.70 ± 0.04 a 1.83 ± 0.01 a 0.58 ± 0.04 b 净氮矿化速率
Net
N mineralization
rate大团聚体
(2 000~8 000 μm)1.45 ± 0.29 c 1.14 ± 0.13 c −0.83 ± 0.13 c 1.51 ± 0.10 a 1.52 ± 0.25 a 0.62 ± 0.10 b 1.53 ± 0.14 a 1.84 ± 0.28 a 0.51 ± 0.10 b 中团聚体
(1 000~2 000 μm)0.92 ± 0.22 b 1.89 ± 0.31 b −0.36 ± 0.12 b 1.11 ± 0.06 b 1.52 ± 0.13 a 0.69 ± 0.11 c 1.15 ± 0.03 C 1.94 ± 0.26 a 0.57 ± 0.09 d 小团聚体
(250~1 000 μm)0.10 ± 0.02 b 4.39 ± 6.75 a 0.31 ± 0.02 b 1.35 ± 0.18 ab 1.48 ± 0.19 a 0.81 ± 0.04 c 1.49 ± 0.24 b 1.71 ± 0.18 ab 0.74 ± 0.04 d 微团聚体
(<250 μm)0.92 ± 0.17 b 0.19 ± 0.04 b −0.84 ± 0.33 b 1.10 ± 0.20 b 1.49 ± 0.25 a 0.70 ± 0.09 c 1.13 ± 0.20 c 1.74 ± 0.32 ab 0.67 ± 0.08 d -
团聚体是土壤基本的结构和功能单位,其粒径大小直接影响水分滞留、气体交换和养分循环,因此具有自身的亚结构和物质微循环机制,兼有调控土壤水热和养分的重要生态功能[20]。土壤团聚体形成的重要基础是有机-无机复合体,因此粒径对其净氮矿化速率的影响主要来自于其自身的有机物质差异[21]。净氨化与净硝化是土壤净氮转化的直接指标[11]。土壤净氨矿化量是氨态氮在培养实验结束时与在培养初始时含量的差值。团聚体净氨化速率相对净硝化速率较低,在某些处理之下出现负值(图1)。这是由于氨态氮是微生物的优先氮源和后续硝化反应的底物,微生物为满足活动需求以及进行硝化反应会不断消耗氨态氮[22],随着培养的进行,当氨态氮的消耗量大于增加量时,净氨矿化量就会出现负值。净硝矿化贡献了85%以上的净氮矿化,因此净硝化与净氮矿化随粒径变化的规律一致(图5)。团聚体中的全氮、有机质以及微生物生物量碳、氮含量均随团聚体粒径减小而增加,而土壤速效磷含量变化趋势与之相反(表1)。这是由于微生物来源的碳水化合物会首先在比表面积更大、吸附力更强的微团聚体中富集[21],这使得微团聚体中全氮和有机质含量会更高[23];凌小莉等[24]和杨坤等[25]发现较小粒径的团聚体会具有更高的微生物生物量,这一结论与实验结果一致,同时更高的微生物生物量意味着小粒径团聚体具有更高的生物酶活性和生化反应能力,这也反映在相同的培养条件下微团聚体的净氮矿化高于其他3个粒径。但更高的微生物生物量会加快土壤中速效磷的消耗,因此速效磷含量从高到低依次是大团聚体>中团聚体>小团聚体>微团聚体。这种现象预警了土壤氮、磷以及有机质的流失将可能最先在微团聚体中出现[19],因此需要开展长期监测,从经营措施的角度考虑,必要时可以施用磷肥。
-
水分和温度是影响氮矿化速率的两个关键非生物因素,二者对土壤氮矿化作用存在显著的交互作用[26]。温度对土壤净氮转化的调控主要是通过土壤微生物和胞外酶活性来实现。低温环境中,温度升高带来微生物活动加剧和胞外酶活性提高,土壤氨化和硝化作用速率不断升高[27];但随温度的不断升高和反应的进行,微生物对碳、磷等养分的需求加剧,此时微生物的活动强度降低主要受到养分的限制[28]。此外,当温度升高过程中会显著提高反硝化的强度,将加速N2O和NO态的氮流失,最终反映为净氮矿化的降低[29]。土壤水分对氮矿化的影响一般存在两个途径;其一是水通过对土壤水分的可用性和氧气通量对微生物活性实现调控,对氮矿化产生直接效应[30-31],其二是通过水分的增加促进可溶性有机物的溶解,提高可矿化态有机氮含量,对氮矿化产生间接效应。在温度分别为5、15、25和35 ℃和土壤水分分别为0.4 FCW、0.6 FCW、0.8 FCW的条件下开展室内矿化培养试验,4个粒径的团聚体均在25 ℃、0.6 FCW条件下具有最大净氮转化速率,之后随温度和水分增加开始下降(图1、3、5),这与Gao等[32]和Wang等[33]的研究一致,也与马芬等[34]在亚热带3种类型森林中的研究结果一致。通过建立二元二次方程,结合不同粒径团聚体的质量比例进行估算,三峡库区马尾松-栓皮栎针阔混交林土壤净氮转化最高速率出现在温度为22.7~24.5℃、土壤水分在0.57 FWC~0.58 FWC之间(图2、4、6),过高或者过低的土壤水分和温度都将会降低土壤净氮转化,这与周于波等[9]、石薇等[35]及桂慧颖等[36]人的研究一致。根据估算,净氨矿化最适温度(20.5~23.0 ℃)低于净硝矿化和净氮矿化的最适温度(22.7~24.5 ℃)。原因是随着硝化反应的进行,氨态氮作为反应底物被氧化导致净氨累积量不断减少,所以在净硝化速率的最适温度下土壤中氨态氮消耗加快,导致氨态氮净积累降低[22]。而团聚体净氨矿化最适土壤水分(0.63 FCW ~ 0.75 FCW)高于净硝矿化和净氮矿化最适土壤水分(0.58 FCW ~ 0.61 FCW图2、4)。这可能是由于当土壤水分达到一定数值时,土壤通气性和氧气含量下降,硝化细菌的活性被抑制,反硝化作用增强,土壤中硝态氮被加速还原为N2O和NO,因此减少了土壤中的净硝化和净氮矿化积累[37]。刘杏认等[38]对3种草地类型土壤研究时发现,水分与土壤净氮矿化速率之间呈正相关,这与本研究结果并不一致,原因是土壤类型和土地利用方式的不同使微生物群落对环境出现适应性差异。
温度敏感性( Q10)用以描述温度每升高10 ℃时土壤氮矿化速率增加的倍数,Wetterstedt等[39]人证实 Q10在不同条件下存在明显差异,因此了解 Q10对于预测气候变化下土壤有效氮动态具有重要意义。在本研究中,4种粒径的团聚体在不同水分处理下净氮矿化速率的 Q10存在显著差异,结合质量比重,Q10整体在15 ℃/25 ℃时最大,范围在1.75~4.39之间(表2)。在对草甸土壤研究中发现,土壤氮矿化速率Q10在5 ℃/15 ℃时最高,当温度继续升高到25 ℃和35 ℃时,Q10下降并逐渐趋于稳定[40],这与本实验的结果并不一致,原因是不同类型的土壤中主导氮矿化过程的微生物群落结构存在差异。根据活化能理论,氮矿化速率 Q10的不同是矿化底物质量差异造成的[9, 35- 36]。土壤氮矿化是通过微生物介导的酶促反应来实现的一个耗能过程, Q10越高说明土壤氮矿化过程中所需的酶促反应步骤越多,映射了矿化基质的质量越差[41]。因此相比于其他土壤理化性质,Q10与土壤有机质的关系更密切。中国陆地生态系统土壤氮矿化速率 Q 10平均值为1.58,其中森林土壤为1.94[40],结合质量比重,三峡库区马尾松-栓皮栎针阔混交林土壤氮矿化 Q10大于中国森林土壤的平均值,这意味着该地区氮矿化反应底物的基质质量可能低于全国森林土壤平均水平[37, 41-42]。微团聚体氮转化 Q10整体最低,最大值同样出现在15 ℃/25 ℃,范围在0.92~1.74,说明微团聚体中的有机质比其他几个粒径的团聚体更易被矿化,这与微团聚体具有最高的氮矿化速率的实验结果相符。根据IPCC报道,气温升高仍然是未来气候变化的趋势,本世纪末平均气温较上世纪末将上升4.4 ℃,并且由于全球变暖,预计降水量也将同步增多[43]。三峡库区典型人工林土壤氮矿化速率在15~25 ℃之间对温度变化最敏感,而该地区年均温度为14~ 22 ℃,处于土壤氮矿化对温度敏感系数最大值的范围内,这预示着在气温不断升高的未来,该地区土壤氮矿化速率将加快。
-
在三峡库区马尾松-栓皮栎针叶混交人工林中,土壤团聚体超过85%的净氮矿化由净硝化贡献。随土壤团聚体粒径的减小,净硝化和净氮矿化速率逐渐增加。团聚体氮矿化对土壤含水量和温度变化的响应显著。当培养条件设置为5 ~35 ℃、0.4 FCW~ 0.8 FCW之间时,土壤净硝化和净氮矿化速率随温度和含水量增加均呈现倒“V”字趋势,拐点分别出现在25 ℃和0.6 FCW。在气温逐渐升高的全球背景之下,结合三峡库区气候特点,短期来看马尾松-栓皮栎针阔混交人工林的土壤净氮矿化作用将加强,有利于提高当地土壤氮的供应能力,使森林生态系统的生产力得到一定提升;但长期处于这种变化下,土壤氮的淋溶风险和N2O等温室气体的排放风险将增加,并引起土壤磷限制,对当地森林生态系统生产力和区域环境产生不利影响。从人工林经营角度考虑应开展长期监测,必要时可考虑施用磷肥以提高生产力。
三峡库区典型人工林土壤团聚体氮矿化对水热变化的响应
Response of Nitrogen Mineralization in Soil Aggregates to Changes in Soil Moisture and Temperature in the Three Gorges Reservoir Area's Typical Plantation
-
摘要:
目的 气温升高和降水增多已成为全球气候正在面临的挑战,为了探究全球气候变化对三峡库区森林土壤氮动态的影响,以期为预测该地区土壤养分循环变化和制定提升森林生产力的经营措施提供理论依据。 方法 以马尾松-栓皮栎混交人工林为研究对象,将土壤筛分为大团聚体、中团聚体、小团聚体和微团聚体(粒径为:2 000~8 000、1 000~2 000、250~1 000和<250 μm),以室内培养方法观察团聚体净氮转化在土壤水分为40%、60%和80% 田间最大持水量(命名为0.4 FCW、0.6 FCW和0.8 FCW),和温度为5、15、25和35 ℃下的差异。 结果 比表面积较大的小粒径团聚体中有机质、总氮含量和微生物生物量碳、氮更高,而其速效磷含量显著低于大团聚体。随粒径减小,团聚体净硝化和净氮矿化速率逐渐增加,其中净硝化贡献了超过85%的净氮矿化。团聚体净硝化和净氮矿化速率随土壤含水量和温度的增加先上升后下降,最大值出现在0.6 FCW和25 ℃。不同粒径团聚体的净氨化、净硝化和净氮矿化速率整体在0.60 FCW和15 ℃/25 ℃时对温度上升最敏感;通过建立二元二次方程,估算出净氨化最大速率出现在土壤水分和温度为0.70 FCW和23.0 ℃,净硝化和净氮矿化最大速率出现在土壤水分和温度为0.58 FCW ~ 0.60 FCW和24.5 ~ 27.5 ℃。 结论 团聚体的物理结构保护了三峡库区人工林土壤有机氮免受微生物分解,而温度上升和降水增多促进了该地区马尾松-栓皮栎针阔混交人工林土壤净氮转化,这将有利于提高土壤的氮供应能力。但长期来看,这种变化催生土壤有机质和氮的流失,增加磷限制的风险,必要时应开展长期监测和施用磷肥措施。 Abstract:Objective Rising temperatures and precipitation have become challenges. In order to investigate the effects of climate change on soil nitrogen (N) dynamics in forests of the Three Gorges Reservoir area and to provide a basis for predicting changes in soil nutrient cycling in this area and enhancing forest productivity. Method The soil was sieved into large macroaggregates, coarse aggregate, small macroaggregates, and microaggregates (particle sizes: 8000-2000, 2000-1000, 1000-250, and <250 μm) in a mixed plantation of Pinus massoniana-Quercus variabilis. Differences in net N conversion of aggregates were observed at soil moisture of 40%, 60%, and 80% of the maximum water holding capacity in the field (named 0.4 FCW, 0.6 FCW, and 0.8 FCW), and at temperatures of 5°C, 15°C, 25°C, and 35°C using indoor incubation methods. Results The levels of soil organic matter (SOM), total N and microbial biomass carbon (C), and nitrogen were higher in small-size aggregates with larger specific surface areas, while available phosphorus (P) was significantly lower. The rates of net nitrification and net N mineralization of aggregates gradually increased with decreasing particle size, with net nitrification contributing more than 85% of net N mineralization. The rates of net nitrification and net N mineralization of aggregates first increased and then decreased with the increase of soil water moisture and temperature, and the maximum values appeared at 0.6 FCW and 25 ℃. The overall net ammonification, net nitrification, and net N mineralization rates of aggregates of different particle sizes were most sensitive to temperature increase at 0.60 FCW and 15°C/25°C. By developing a quadratic equation, the optimum soil moisture and temperature for the net ammonification rate were 0.70 FCW and 23.0°C, and the optimum soil moisture and temperature for the net nitrification and net nitrogen mineralization rates were 0.58 FCW - 0.60 FCW and 24.5°C - 27.5°C. Conclusion The physical structure of aggregates protects organic N from decomposition in the plantation in the Three Gorges Reservoir area. Rising temperatures and precipitation promote net N transformation in the soils, which is conducive to improving the N supply capacity of the soil; However, there is a risk of soil organic matter, N loss, and P limitation over the long term, so that monitoring and fertilizer measures can be carried out if necessary. -
Key words:
- Three Gorges Reservoir Area
- / aggregates
- / N mineralization
- / temperature
- / moisture
-
图 1 团聚体在不同水热条件下净氨化速率特征
Figure 1. Characteristics of net ammonification rate of agglomerates under different soil moisture and temperature conditions.
图 2 团聚体净氨化速率对土壤水分和温度变化响应模式
Figure 2. Response pattern of net ammonification rate of aggregates to changes in soil moisture and temperature
图 3 团聚体在不同水热条件下净硝化速率特征
Figure 3. Characteristics of net nitrification rate of aggregates under different soil moisture and temperature conditions
图 4 团聚体净氨化速率对土壤水分和温度变化响应模式
Figure 4. Response pattern of net nitrification rate of aggregates to changes in soil moisture and temperature
图 5 团聚体在不同水热条件下净氮矿化速率特征
Figure 5. Characteristics of net N mineralization rate of aggregatess under different soil moisture and temperature conditions
图 6 团聚体净氨化速率对土壤水分和温度变化响应模式
Figure 6. Response pattern of net N mineralization rate of aggregates to changes in soil moisture and temperature
表 1 不同粒径团聚体理化特征
Table 1. Physicochemical characteristics of aggregates with different particle sizes
大团聚体
Large macroaggregates中团聚体
Coarse aggregate小团聚体
Small macroaggregates微团聚体
Microaggregate全氮
Total N/ (g·kg−1)1.41 ± 0.14 b 2.55 ± 0.26 a 2.76 ± 0.28 a 2.81 ± 0.57 a 土壤有机质
Soil organic matter/(g·kg−1)17.79 ± 2.61 c 24.85 ± 3.12 b 29 ± 5.01 ab 32.13 ± 3.38 a 速效磷
Available phosphorus/(mg·kg−1)13.11 ± 0.55 a 13.61 ± 0.48 a 13.35 ± 0.33 ab 11.81 ± 1.15 b 速效钾
Available potassium/(mg·kg−1)103.47 ± 1.21 a 102.7 ± 1.35 a 102.3 ± 2.15 a 103 ± 1.18 a 土壤酸碱度
pH (H2O)5.35 ± 0.26 a 5.02 ± 0.08 a 5.12 ± 0.1 a 5.28 ± 0.34 a 微生物量碳
Microbial biomass C/(mg·kg−1)244.69 ± 41.15 c 252.76 ± 15.97 bc 321.72 ± 62.34 ab 376.61 ± 7.15 a 微生物量氮
Microbial biomass N/(mg·kg−1)40.49 ± 0.16 b 44.64 ± 4.34 b 56.97 ± 5.65 a 64.38 ± 6.47 a 质量比例
Weight proportions
of aggregates in soil0.21 ± 0.04 c 0.29 ± 0.29 b 0.39 ± 0.04 a 0.11 ± 0.01 d 注:不同小写字母标注代表在0.05检验水平上差异显著,下同
Notes: Different small letters indicate significant difference at 0.05 test level, the same below 下载: 导出CSV
下载: 导出CSV
表 2 团聚体净氨化、净硝化和净氮矿化速率Q10在不同土壤水分下的变化
Table 2. Fluctuate in Q10 values in aggregates of net ammonification, net nitrification, and net nitrogen mineralization rates under different soil moisture treatments
0.4 FCW 0.6 FCW 0.8 FCW 5 ℃/15 ℃ 15 ℃/25 ℃ 25 ℃/35 ℃ 5 ℃/15 ℃ 15 ℃/25 ℃ 25 ℃/35 ℃ 5 ℃/15 ℃ 15 ℃/25 ℃ 25 ℃/35 ℃ 净氨化速率
Net
ammonification
rate大团聚体
(2 000~8 000 μm)0.69 ± 0.14 b 2.75 ± 0.93 b −1.39 ± 0.32 b 1.25 ± 0.28 c 1.26 ± 0.28 c 1.25 ± 0.26 c 1.36 ± 0.30 b 1.64 ± 0.29 b 0.83 ± 0.15 cd 中团聚体
(1 000~2 000 μm)0.31 ± 0.07 b 5.65 ± 11.49 a 0.01 ± 0.03 b 1.47 ± 0.24 bc 1.89 ± 0.31 ab 0.76 ± 0.05 de 1.55 ± 0.24 b 2.24 ± 0.36 a 0.65 ± 0.05 d 小团聚体
(250~1 000 μm)1.08 ± 0.11 b 1.97 ± 0.13 b −1.22 ± 0.20 b 1.17 ± 0.08 cd 1.92 ± 0.36 a 0.70 ± 0.11 e 1.18 ± 0.08 bc 2.49 ± 0.52 a 0.48 ± 0.09 d 微团聚体
(<250 μm)0.49 ± 0.2 bc 1.41 ± 0.49 a −0.92 ± 0.05 e 1.58 ± 0.31 a 1.43 ± 0.29 a 0.57 ± 0.08 b 2.02 ± 0.59 a 1.68 ± 0.42 a 0.47 ± 0.07 b 净硝化速率
Net
nitrification
rate大团聚体
(2 000~8 000 μm)−0.62 ± 0.15 de −4.84 ± 1.13 f 0.09 ± 0.00 cd 1.71 ± 0.39 a 1.66 ± 0.21 a 0.74 ± 0.08 b 1.96 ± 0.49 a 1.76 ± 0.23 a 0.68 ± 0.07 b 中团聚体
(1 000~2 000 μm)0.93 ± 0.17 ab 0.33 ± 0.14 bc −4.32 ± 0.24 f 1.69 ± 0.44 a 1.35 ± 0.25 a 0.57 ± 0.06 b 1.92 ± 0.71 a 1.55 ± 0.37 a 0.47 ± 0.07 b 小团聚体
(250~1 000 μm)0.06 ± 0.01 c 3.19 ± 6.10 a −0.39 ± 0.02 c 1.52 ± 0.27 a 1.56 ± 0.25 a 0.64 ± 0.04 b 1.76 ± 0.33 a 1.74 ± 0.28 a 0.54 ± 0.03 b 微团聚体
(<250 μm)9.46 ± 3.71 b −2.11 ± 0.67 c 0.11 ± 0.02 c 1.58 ± 0.06 a 1.81 ± 0.06 a 0.62 ± 0.04 b 1.70 ± 0.04 a 1.83 ± 0.01 a 0.58 ± 0.04 b 净氮矿化速率
Net
N mineralization
rate大团聚体
(2 000~8 000 μm)1.45 ± 0.29 c 1.14 ± 0.13 c −0.83 ± 0.13 c 1.51 ± 0.10 a 1.52 ± 0.25 a 0.62 ± 0.10 b 1.53 ± 0.14 a 1.84 ± 0.28 a 0.51 ± 0.10 b 中团聚体
(1 000~2 000 μm)0.92 ± 0.22 b 1.89 ± 0.31 b −0.36 ± 0.12 b 1.11 ± 0.06 b 1.52 ± 0.13 a 0.69 ± 0.11 c 1.15 ± 0.03 C 1.94 ± 0.26 a 0.57 ± 0.09 d 小团聚体
(250~1 000 μm)0.10 ± 0.02 b 4.39 ± 6.75 a 0.31 ± 0.02 b 1.35 ± 0.18 ab 1.48 ± 0.19 a 0.81 ± 0.04 c 1.49 ± 0.24 b 1.71 ± 0.18 ab 0.74 ± 0.04 d 微团聚体
(<250 μm)0.92 ± 0.17 b 0.19 ± 0.04 b −0.84 ± 0.33 b 1.10 ± 0.20 b 1.49 ± 0.25 a 0.70 ± 0.09 c 1.13 ± 0.20 c 1.74 ± 0.32 ab 0.67 ± 0.08 d
下载: 导出CSV
-
[1] CHENG Y, WANG J, CHANG S X, et al. Nitrogen deposition affects both net and gross soil nitrogen transformations in forest ecosystems: A review[J]. Environmental Pollution, 2019, 244: 608-616. [2] WEN Y, TANG Y, WEN J , et al. Variation of intra‐aggregate organic carbon affects aggregate formation and stability during organic manure fertilization in a fluvo‐aquic soil[J]. Soil Use and Management, 2020, 37(1): 151-163. [3] 陈 天, 程瑞梅, 王丽君, 等. 氮添加对马尾松人工林土壤团聚体氮矿化及土壤酶活性的影响[J]. 生态学报, 2023, 43(16):6528-6538. [4] BUCKA F B, KÖLBL A, UTEAU D, et al. Organic matter input determines structure development and aggregate formation in artificial soils[J]. Geoderma, 2019, 354: 113881. [5] XIAO R, DUAN B, MAN X, et al. Highly sensitive net nitrogen mineralization to soil temperature and moisture during a boreal secondary forest succession[J]. Journal of Soils and Sediments, 2023, 23(3): 1169-1181. [6] PAJUSTE K, FREY J. Nitrogen mineralization in podzol soils under boreal Scots pine and Norway spruce stands[J]. Plant and Soil, 2003, 257(1): 237-247. doi: 10.1023/A:1026222831694 [7] BAI E, LI S, XU W, et al. A meta-analysis of experimental warming effects on terrestrial nitrogen pools and dynamics[J]. New Phytologist, 2013, 199(22): 441-451. doi: 10.1111/nph.12252 [8] 徐星凯, 韩 琳, 罗献宝. 模拟氮沉降对温带阔叶红松林地氮素净矿化量的影响[J]. 气候与环境研究, 2012, 17(5):628-638. [9] 周于波, 王景燕, 黄 帅, 等. 水热条件对华西雨屏区檫木人工林土壤氮矿化的影响[J]. 云南农业大学学报(自然科学), 2019, 34(3):479-485. [10] 王 鸽, 贾志军, 韩 琳. 全球森林土壤净矿化量的影响因子[J]. 环境科学与技术, 2014, 37(120):9-14. [11] STANFORD G, EPSTEIN E. Nitrogen mineralization-water relations in soils[J]. Soil Science Society of America Proceedings, 1974, 38(1): 103-107. [12] KNOEPP J D, SWANK W T. Using soil temperature and moisture to predict forest soil nitrogen mineralization[J]. Biology and Fertility of Soils, 2002, 36(3): 177-182. [13] GUNTIÑAS M E, LEIRÓS M C, TRASAR-CEPEDA C, et al. Effects of moisture and temperature on net soil nitrogen mineralization: A laboratory study[J]. European Journal of Soil Biology, 2012, 48(1): 73-80. [14] KNOEPP J D, VOSE J M. Regulation of nitrogen mineralization and nitrification in southern Appalachian ecosystems: separating the relative importance of biotic vs. abiotic controls[J]. Pedobiologic, 2007, 51(2): 89-97. doi: 10.1016/j.pedobi.2007.02.002 [15] 朱建华, 田 宇, 李 奇, 等. 中国森林生态系统碳汇现状与潜力[J]. 生态学报, 2023, 43(9):3442-3457. [16] 中国气象局气候变化中心. 中国气候变化蓝皮书(2022)[R]. 北京: 中国气象局. 2022. [17] 程书波, 李 冲, 岳 颖, 等. 1961—2020年我国中部地区气温和降水时空变化特征[J]. 水利水电技术(中英文), 2023, 54(6):75-86. [18] 王丽君, 程瑞梅, 肖文发, 等. 氮添加对三峡库区马尾松-栓皮栎混交林土壤微生物生物量和酶活性的影响[J]. 应用生态学, 2022, 33(1):42-50. [19] BACH E M, HOFMOCKEL K S. Soil aggregate isolation method affects measures of intra-aggregate extracellular enzyme activity[J]. Soil Biology and Biochemistry, 2014, 69(1): 54-62. [20] 陈 天, 程瑞梅, 沈雅飞, 等. 氮添加对三峡库区马尾松人工林土壤团聚体有机氮组分和氮矿化的影响[J]. 应用生态学报, 2023, 34(10):2601-2609. [21] CAO R, CHEN L C, HOU X C, et al. Nitrogen addition reduced carbon mineralization of aggregates in forest soils but enhanced in paddy soils in South China[J]. Ecol Processes, 2021, 10(1): 1-11. [22] 刘君政, 王 鹏, 肖汉玉, 等. 中国陆地生态系统土壤氮矿化速率和硝化速率及影响因素——基于文献数据的统计分析[J]. 生态学报, 2020, 40(12):12. [23] POLLÁKOVÁ N, ŠIMANSKÝ V V, KRAVKA M. The influence of soil organic matter fractions on aggregates stabilization in agricultural and forest soils of selected Slovak and Czech hilly lands[J]. Journal of Soils and Sediments, 2017, 18(8): 2790-2800. [24] 凌小莉, 史宝库, 崔海莹, 等. 氮磷添加对松嫩草地土壤团聚体结构及其碳含量的影响[J]. 中国草地学报, 2021, 43(2):54-63. [25] 杨 坤, 陈佳广, 关连珠, 等. 不同利用方式下棕壤及其各级微团聚体中微生物量碳、氮的变化[J]. 中国农学通报, 2006(1):185-187. [26] GUNTIÑAS M E, LEIRÓS M C, TRASAR-CEPEDA C, et al. Effects of moisture and temperature on net soil nitrogen mineralization: A laboratory study[J]. European Journal of Soil Biology, 2012, 48(3): 73-80. [27] CHEN T, CHENG R M, XIAO W F, et al. Nitrogen addition enhances soil nitrogen mineralization through an increase in mineralizable organic nitrogen and the abundance of functional genes[J]. Journal of Soil Science Plant nutrition, 2024, https://doi.org/10.1007/s42729-023-01600-0. [28] MARKLEIN A R, WINBOURNE J B, ENDERS S K, et al. Mineralization ratios of nitrogen and phosphorus from decomposing litter in temperate versus tropical forests[J]. Global Ecology and Biogeography, 2016, 25(3): 335-346. [29] LI Y, LIU Y H, WANG Y L, et al. Interactive effects of soil temperature and moisture on soil N mineralization in a Stipa krylovii grassland in Inner Mongolia, China[J]. Journal of Arid Land, 2014, 6(5): 571-580. doi: 10.1007/s40333-014-0025-5 [30] FUJITA Y, van BODEGOM P M, VENTERINK H O, et al. Towards a proper integration of hydrology in predicting soil nitrogen mineralization rates along natural moisture gradients[J]. Soil Biology and Biochemistry, 2013, 58: 302-312. [31] 朱寒松. 土壤和团聚体氮素矿化对水热条件的响应特征研究[D]. 杨凌, 西北农林科技大学, 2018. [32] GAO J Q, OUYANG H, ZHANG F, et al. The response of soil nitrogen mineralization to soil temperature and soil moisture in Zoige alpine wetland[J]. Wetland Science, 2008, 6(2): 229-234. [33] WANG C, WAN S, XING X, et al. Temperature and soil moisture interactively affected soil net N mineralization in temperate grassland in Northern China[J]. Soil Biology Biochemistry, 2006, 38(5): 1101-1110. [34] WANG C, WAN S, XING X, et al. Temperature and soil moisture interactively affected soil net N mineralization in temperate grassland in Northern China[J]. Soil Biology & Biochemistry, 2006, 38(5): 1101-1110. [35] 马 芬, 马红亮, 邱 泓, 等. 水分状况与不同形态氮添加对亚热带森林土壤氮素净转化速率及N2O排放的影响[J]. 应用生态学报, 2015, 26(2):379-387. [36] 石 薇, 王景燕, 魏有波, 等. 水热条件对华西雨屏区柳杉人工林土壤氮矿化的影响[J]. 土壤通报, 2014, 45(6):1430-1436. [37] 桂慧颖, 李雪江, 王景燕, 等. 温度和水分对华西雨屏区毛竹林土壤氮矿化的影响[J]. 四川农业大学学报, 2018, 36(6):758-764. [38] 魏丽云. 土壤水分动态对氮素净矿化的影响[D]. 杨凌, 西北农林科技大学, 2019. [39] 刘杏认, 董云社, 齐玉春, 等. 温带典型草地土壤净氮矿化作用研究[J]. 环境科学, 2007, 28(3):633-639. [40] LIU Y, TAN X P, Wang Y Y, et al. Responses of litter, organic and mineral soil enzyme kinetics to 6 years of canopy and understory nitrogen additions in a temperate forest[J]. Science of The Total Environment, 2020, 712: 136383. [41] 赵 宁, 张洪轩, 王若梦, 等. 放牧对若尔盖高寒草甸土壤氮矿化及其温度敏感性的影响[J]. 生态学报, 2014, 34(15):4234-4241. [42] ZANG H D, BLAGODATSKAYA E, WEN Y, et al. Temperature sensitivity of soil organic matter mineralization decreases with long-term N fertilization: Evidence from four Q 10 estimation approaches[J]. Land Degradation & Development, 2020, 31(6): 683-693. [43] IPCC. Climate change 2021: The physical science basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change[M]. Cambridge and New York: Cambridge University Press (In Press), 2021. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1102
- HTML全文浏览量: 240
- PDF下载量: 24
- 被引次数: 0









