

-
木麻黄(Casuarina equisetifolia Forst.)属于木麻黄科,是一种常绿乔木植物,最初产自大洋洲、太平洋岛屿以及东南亚地区,是亚热带地区沿海防护林的首选树种,因其耐干旱、耐盐碱、耐贫瘠特性,在保持水土、防风固沙、减少海浪侵蚀等方面发挥着重要作用[1-2]。自上世纪20年代从澳大利亚引种到广东,目前我国东南沿海7 230 km长的海岸线上已营造了16万公顷木麻黄人工林[3-4]。东山、惠安、平潭等地建立多条防护林带,在防风固沙、改善环境方面产生了巨大的生态效益。木麻黄生长快、材积高,材质坚硬,满足人们对用材林需求的同时,还能显著提升农田作物产量,具有显著的经济效益[5]。在生态服务价值方面,木麻黄林属于人为干扰下达到相对稳定的偏途顶级群落,在脆弱的滨海生态系统中尚无其它树种可以替代[6]。因此,木麻黄资源的可持续利用在我国现代林业的健康发展以及维持滨海生态系统的稳定方面起着至关重要的作用。然而,随着木麻黄人工林的多代连续种植,连栽障碍问题(Continuous planting obstacle)近年来引起林业和生态学者的广泛关注。国内外有关人工林连栽障碍屡有报道,国外的报道涉及到了辐射松、火炬松、湿地松、红皮云杉和日本柳杉等树种,而国内大面积种植的杨树、杉木、桉树和落叶松等树种也有相关的研究[7]。连栽作为木麻黄人工林最主要的经营方式,随着连栽代数的增加,会导致植株生长发育不良,病虫害严重[8-9]。据调查显示,连栽引起福建东山、惠安等地木麻黄林的生长量显著下降,14年生木麻黄二代林与头代相比,树高、胸径和材积分别降低了23.7%、24.4%和29.0%,19年生木麻黄连栽与头栽相比,平均树高、胸径和材积生长分别减少了8.36%、7.61%和10.30%,且存在显著差异[10-11]。目前,我国南方20世纪80~90年代营造的大面积木麻黄人工林,均面临二代、三代更新问题。连栽木麻黄的高产高质是一项科学性难题,解决连栽障碍机制及其调控是木麻黄人工林栽培的一个重要科学问题,在国内外同行的研究中备受关注。
根际土壤是植物体的重要生境,根际微生物群落的宏基因组是植物微生物组的重要组成部分[12-13]。目前,植物根际微生物组研究的重点主要是细菌和真菌,而古菌经常被忽略。古菌被认为仅分布于地球上的极端自然环境中,随着研究的不断深入,在土壤、湿地、海水等不同的生态系统中都发现古菌的存在,且对元素的生物地球化学循环具有重要的驱动作用[14-15]。古菌是微生物群落中的一部分,适应各种条件,从而定居土壤、动植物和人体[16]。在土壤中,受土壤类型和土层的影响,古菌丰度、群落结构和分布特征可能有所不同[17-18]。目前有关森林生态系统中人工林连栽障碍下根际土壤古菌群落变化的研究鲜见报道。以往对木麻黄人工林连栽障碍的研究表明,随着连栽代数的增加,根际细菌和放线菌的数量显著减少,而真菌的数量明显增加,这导致了土壤微生物群落类型从以细菌为主向以真菌为主过渡[19-22]。然而,连栽条件下木麻黄人工林的根际土壤古菌群落是如何变化的还未见报道。因此,本研究以3个不同连栽代数(第一代First continuous plantation,FCP;第二代Second continuous plantation,SCP;第三代Third continuous plantation,TCP)的木麻黄根际土壤为材料,以未种植木麻黄的天然乔木林根际土壤作为对照(CK),采用高通量测序技术对古菌群落进行16S rDNA扩增子测序,通过物种注释和相对丰度分析,探究连栽木麻黄根际土壤古菌群落变化规律,以期为探明木麻黄连栽障碍机制提供基础理论依据。
-
研究样地所处地点在福建省泉州市惠安县赤湖国有防护林场,占地面积约为433 hm2,属南亚热带海洋性气候,年降雨量为1 029 mm,年蒸发量达2 000 mm,年平均气温为19.8℃,极高气温和极低气温分别为35℃和1℃[23]。林地上种植了3个不同代数的木麻黄人工林,分别是一代林(FCP)、二代林(SCP)和三代林(TCP)。在每一代林内,木麻黄的林龄是统一的,而不同代数间的林龄是不同的。每一代的样地未经过炼山整地,栽植苗为木麻黄嫩枝扦插培育的幼苗,伴生树种与林下植被基本一致。基本信息如表1所示。
表 1 木麻黄人工林样地基本情况
Table 1. Basic information of the C. equisetifolia plantation sample plots
样地
Sample plot栽植时间
Planting time方位
Position伴生树种
Associated tree species林下植被
Underforest vegetation土壤质地Soil texture CK — 118°90′43″ E,24°91′38″ N 台湾栾树(Koelreuteria elegans (Seem.) A. C. Smith ssp. formosana (Hayata) Meyer)和潺槁木姜子(Litsea glutinosa (Lour.) C. B. Rob.) 霍香蓟(Ageratum conyzoides L.)和鬼针草(Bidens pilosa L.) 风积黄沙土 FCP 1987年 118°90′52″ E,24°91′45″ N SCP 2011年 118°90′56″ E,24°91′47″ N TCP 2014年 118°90′31″ E,24°91′56″ N 注:CK:对照;FCP:一代林;SCP:二代林;TCP:三代林。下同。
Note: CK: Control; FCP: First continuous plantation; SCP: Second continuous plantation; TCP: Third continuous plantation. The same below. -
2023年2月,在赤湖国有防护林场内对不同代数木麻黄林地和天然乔木林设置实验样地,即在CK、FCP、SCP和TCP分别设立一个20 m × 20 m的实验样地,每个样地内随机划分3个5 m × 5 m的样方,共计12个样方。木麻黄根际土壤的采集参照王圳等的方法进行[24]。具体为,在每个样方内沿“S”形路径随机选择长势相近的木麻黄,移除土壤表面的树叶和杂质,用铁锹挖去周围土壤,用毛刷轻轻刷取并收集粘附在细根上的土壤,即木麻黄根际土壤。每个样方内的根际土壤摇匀后得到1份土壤样品,共计获得12份。过2 mm筛后,一部分立即用于土壤DNA的提取,一部分自然风干用于测定土壤基本理化性质,剩余的土样保存于−80℃冰箱中备用。土壤基本理化性质如表2所示。
表 2 研究区样地土壤基本理化指标
Table 2. Basic physical and chemical indexes of soil in the study area
指标Index CK FCP SCP TCP pH 5.17 ± 0.00a 5.15 ± 0.00a 4.94 ± 0.01b 4.93 ± 0.01b 全氮Total nitrogen(TN)(mg·g−1) 0.45 ± 0.01a 0.50 ± 0.02a 0.38 ± 0.02b 0.33 ± 0.02b 全磷Total phosphorus(TP)(mg·g−1) 0.39 ± 0.02ab 0.41 ± 0.00a 0.36 ± 0.01bc 0.33 ± 0.01c 全钾Total potassium(TK)(mg·g−1) 1.08 ± 0.05a 1.04 ± 0.02a 0.74 ± 0.03b 0.57 ± 0.03c 速效氮Alkaline nitrogen(AN)(mg·kg−1) 65.57 ± 0.28c 72.71 ± 0.52a 67.93 ± 0.24b 66.34 ± 0.30c 速效磷Available phosphorus(AP)(mg·kg−1) 6.39 ± 0.02c 6.70 ± 0.01a 6.45 ± 0.01b 6.26 ± 0.01d 速效钾Available potassium(AK)(mg·kg−1) 50.65 ± 0.78a 49.45 ± 0.29a 50.45 ± 0.11a 50.38 ± 0.19a 注:同行不同字母表示差异显著(P<0.05)。
Note: Different letters in the same line meant significant difference at 0.05 level. -
土壤微生物基因组总DNA采用BioFast Soil Genomic DNA Extraction Kit试剂盒(BioFlux公司,中国)进行提取。样品DNA浓度采用Nanodrop紫外分光光度计测定,并用1.2%琼脂糖凝胶电泳检测样品DNA的质量。以稀释至1 ng·μL−1的样品DNA为模板,采用特异性引物对古菌16S V4~V5区进行PCR扩增,引物序列分别为524F(5-barcode + TGYCAGCCGCCGCGGTAA-3)和958R(5-CCGGCGTTGAVTCCAATT-3)。使用2%琼脂糖凝胶电泳来检验PCR扩增产物的质量,选择合格的PCR产物送到北京诺禾至源科技有限公司进行16S rDNA扩增子高通量测序。
-
为了完成整个文库制备工作,PCR产物需要经过末端修复、加A尾、加测序接头、纯化等一系列步骤,然后利用Illumina NovaSeq测序平台对文库进行双末端测序。首先,根据barcode对样品进行拆分,以获取每个样品的原始数据,并去除其中的barcode和引物。使用FLASH软件将R1和R2序列数据进行拼接。对于拼接后的Tags,我们进行质控处理,得到Clean Tags。然后,对Clean Tags进行嵌合体过滤,以获得可用于后续分析的有效数据,即Effective Tags。最后,我们基于这些有效数据使用DADA2进行降噪处理,从而得到了最终的ASVs[25]。随后对得到的有效数据进行物种注释和丰度分析,比较不同样本之间的群落结构差异。
-
采用QIIME2软件绘制样品稀释曲线图,并分析 Chao1、Simpson和Shannon多样性指数。花瓣图、柱形图采用Perl 5.26.2 软件进行绘制。热图采用R 4.0.3进行绘制。利用
https://www.bioincloud.tech/ 云平台进行相关性分析。利用DPS 7.05软件进行单因素方差分析和LSD多重比较,并进行组间差异显著性检验。 -
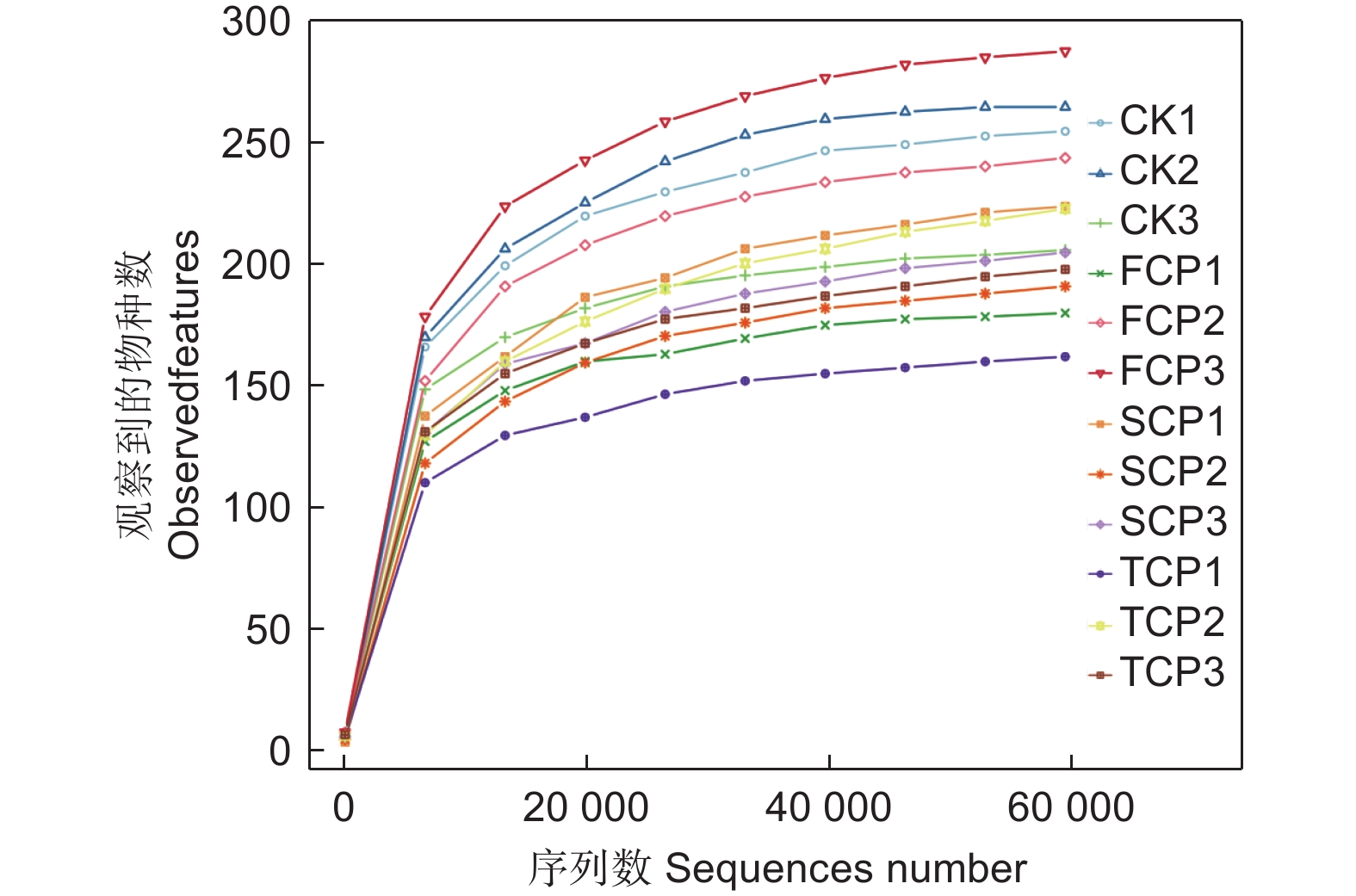
采用Illumina NovaSeq测序平台对不同连栽代数木麻黄根际土壤和未种植木麻黄的天然乔木林土壤的古菌群落进行高通量测序。总共得到1 146 390条有效序列,平均每个样品的有效序列为95 533条。使用DADA2方法进行降噪,并以100%序列相似度聚类,共得到998 515个ASV,其中CK、FCP、SCP、TCP的平均ASV数量分别为63 362、65 824、97 647、106 006。由图1可知,抽取的序列条数将近60 000条,曲线趋向平坦,表明不同土壤样品所测的序列都能够较好地反映古菌群落种类数量,测序数据量渐进合理。
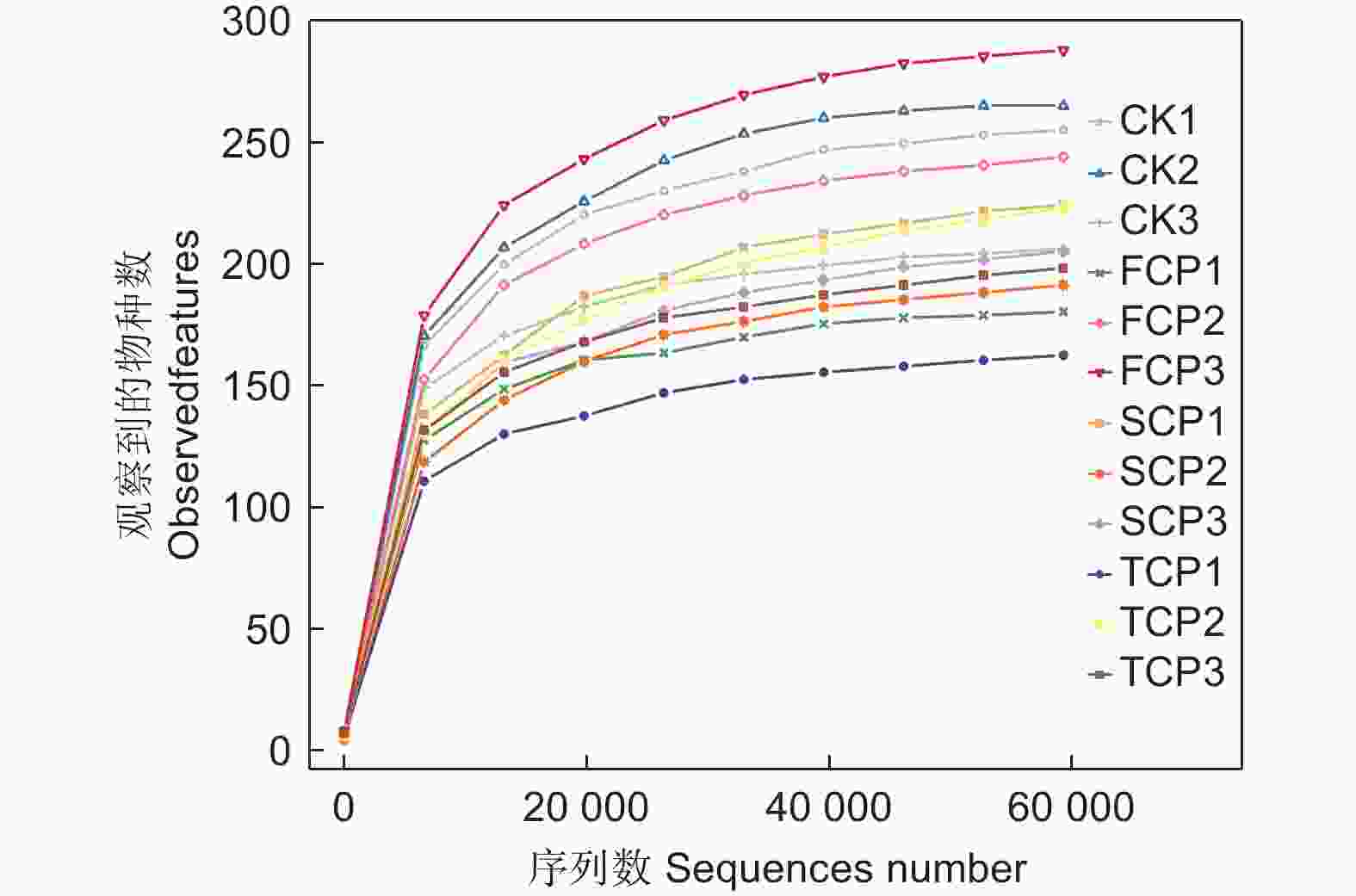
图 1 木麻黄根际土壤古菌群落稀释曲线
Figure 1. Rarefaction curves of the archaeal communities in rhizosphere soil of C. equisetifolia
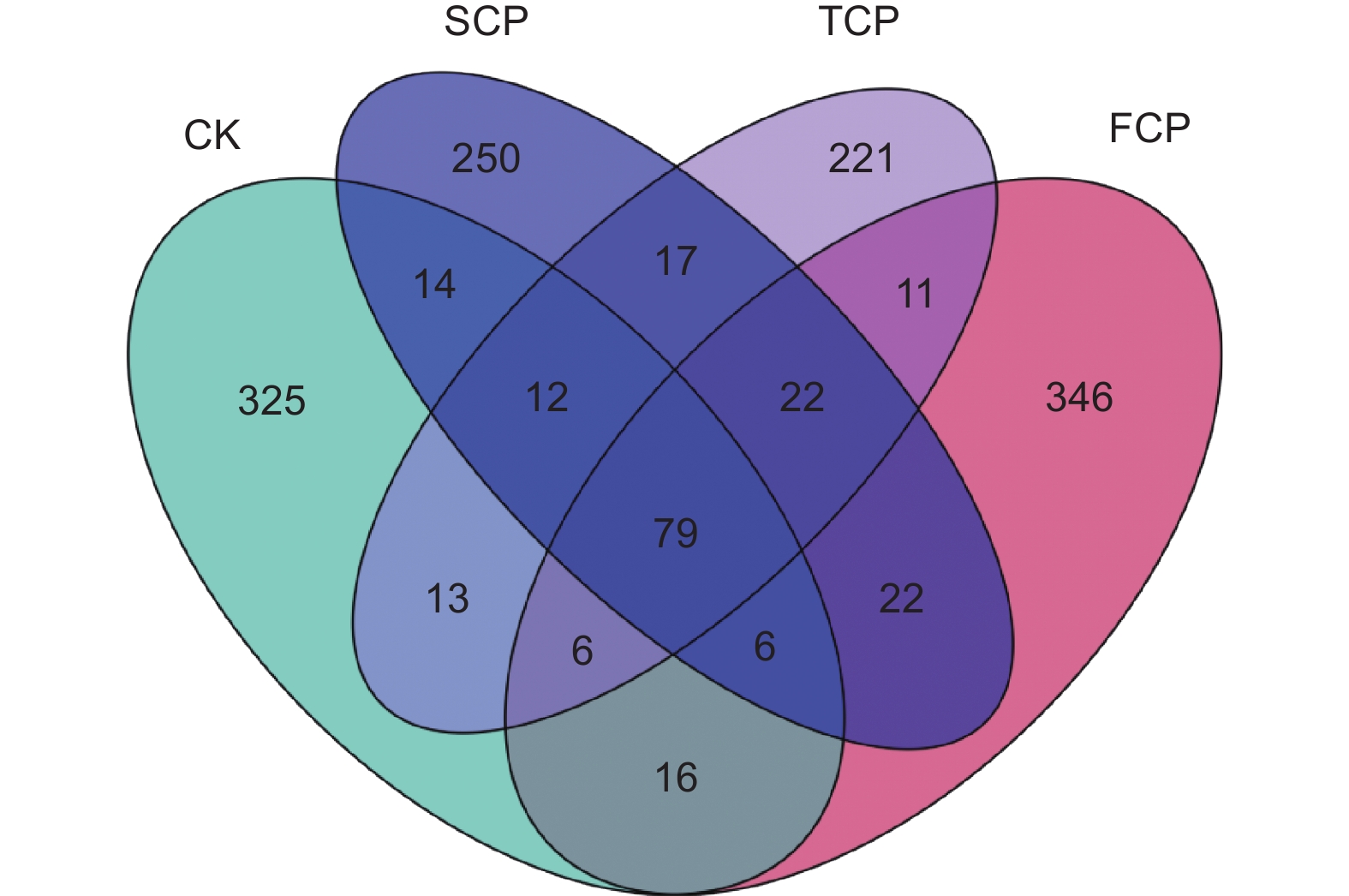
花瓣图可以直观地体现不同连栽代数木麻黄根际土壤古菌群落ASV组成的差异性及重叠情况(图2)。分析结果表明,CK、FCP、SCP和TCP中特异性古菌ASV分别占总ASV序列数的23.90%(325)、25.44%(346)、18.38%(250)和16.25%(221)。CK与FCP、SCP、TCP共有的ASV数量为107(7.87%)、111(8.16%)、110(8.09%),FCP与SCP、TCP共有的ASV数量为129(9.49%)、118(8.68%),SCP与TCP共有的ASV数量为130(9.56%),CK、FCP、SCP和TCP土壤中共有的古菌ASV数量为79(5.81%)。除此之外,随着连栽代数的增加,特异ASV数量呈现下降的趋势,表明木麻黄根际土壤古菌群落结构发生了变化,且在不同代数之间存在明显差异。
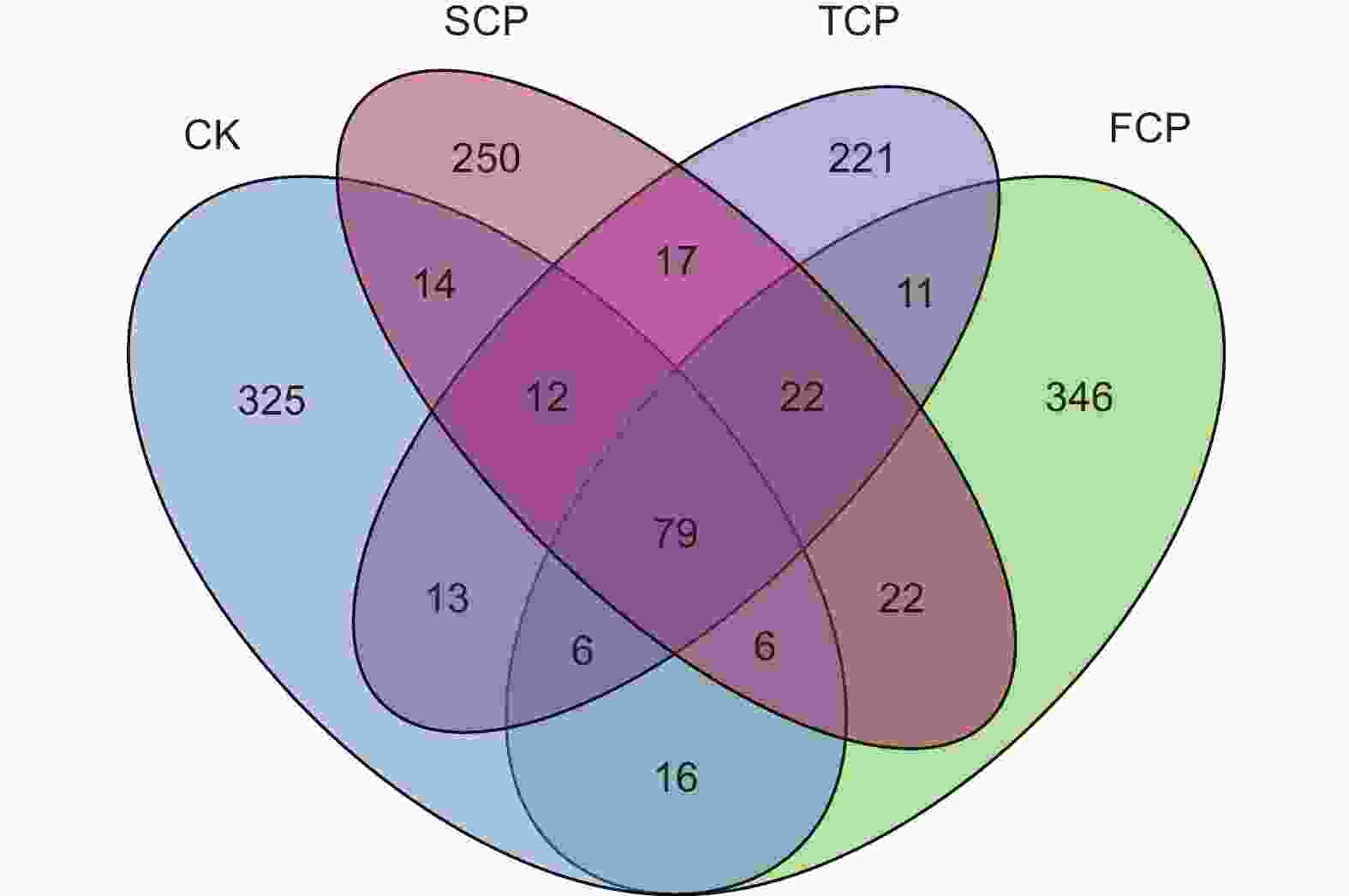
图 2 基于ASVs丰度的木麻黄根际土壤古菌群落花瓣图
Figure 2. Petal plot of archaeal communities in rhizosphere soil of C. equisetifolia based on ASVs abundance
-
对不同连栽代数木麻黄根际土壤古菌群落进行Alpha多样性分析,评估微生物群落的丰富度和多样性的差异。分析结果表明,所观察到的物种数和Chao1指数均随着连栽代数的增加呈现下降的趋势,表明物种丰富度发生了明显变化,但不同代数之间均没有显著差异(表3)。Shannon指数、Simpson指数和pielou_e均以FCP最高,TCP次之,SCP最低,且不同代数之间差异均显著(P<0.05),表明不同代数木麻黄根际土壤古菌群落的多样性和均匀度存在显著差异(P<0.05),且均呈现先下降后上升的趋势。
表 3 连栽木麻黄根际土壤古菌群落丰富度和多样性指数
Table 3. Richness and diversity indices of archaeal communities in rhizosphere soils of continuous rotations of C. equisetifolia
土壤样品
Soil sample观察到的物种
Observed speciesChao1指数
Chao1 index香农指数
Shannon index辛普森指数
Simpson index均匀度指数
Pielou_eCK 243 ± 18a 247.341 ± 17.159a 4.789 ± 0.062a 0.905 ± 0.006a 0.605 ± 0.004a FCP 238 ± 31a 243.231 ± 31.191a 5.001 ± 0.043a 0.943 ± 0.001a 0.636 ± 0.011a SCP 208 ± 10a 221.002 ± 8.894a 3.226 ± 0.143c 0.695 ± 0.027c 0.419 ± 0.018c TCP 195 ± 18a 208.878 ± 23.102a 4.164 ± 0.040b 0.858 ± 0.004b 0.548 ± 0.012b 注:同列不同字母表示差异显著(P<0.05)。
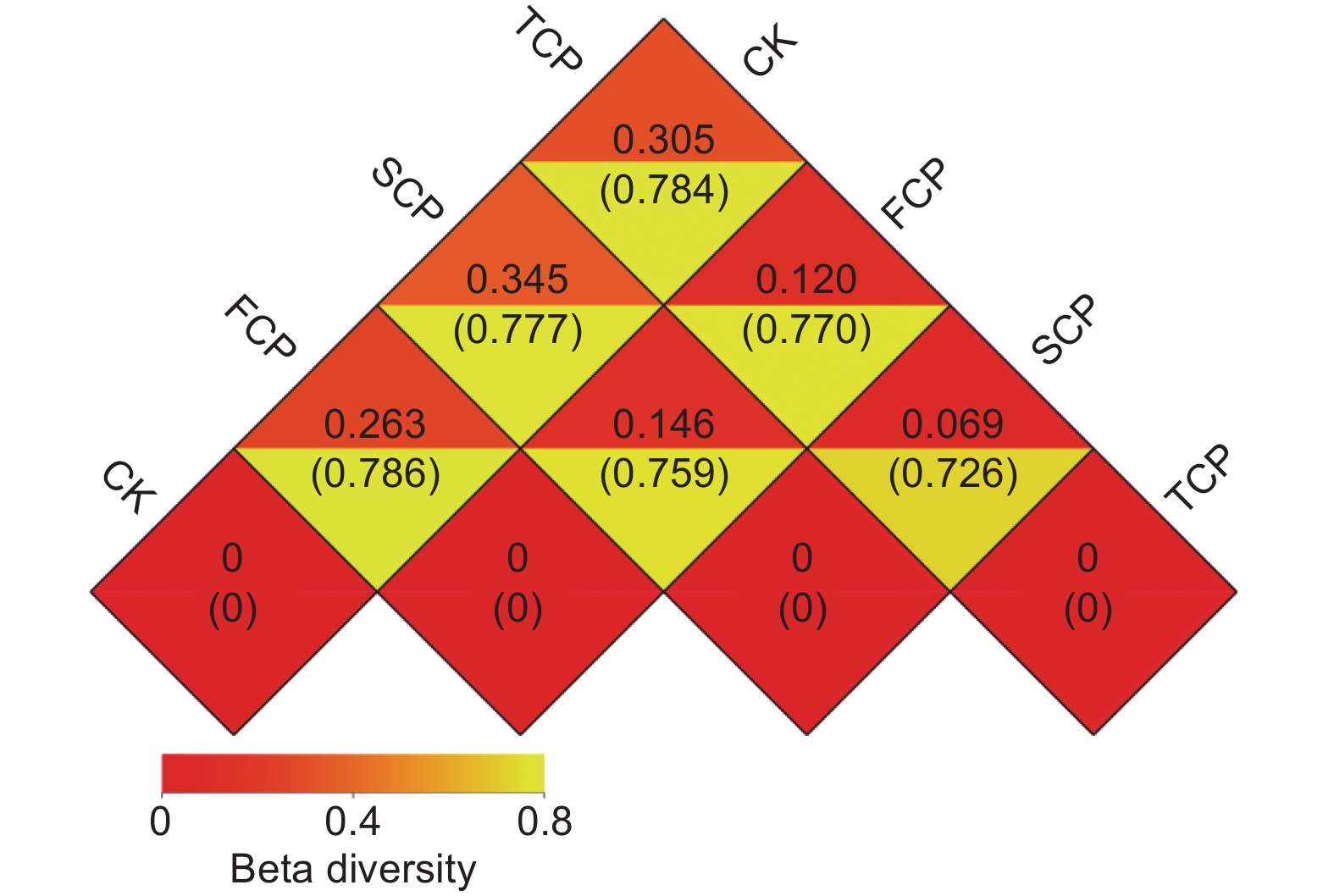
Note: Different letters in the same column meant significant difference at 0.05 level.使用Weighted Unifrac距离和Unweighted Unifrac距离来计算样本之间的相异系数,这些距离的值越小,表示两个样本在物种多样性方面的差异越小。分析结果显示,CK和FCP、SCP、TCP之间的Weighted Unifrac距离分别为0.263、0.345、0.305,呈现先上升后下降的趋势,而Unweighted Unifrac距离分别为0.786、0.777、0.784,呈现先下降后上升的趋势(图3)。FCP和SCP、TCP之间的Weighted Unifrac距离和Unweighted Unifrac距离分别为0.146和0.759、0.120和0.770,SCP和TCP之间分别为0.069和0.726,表明不同连栽代数木麻黄根际土壤古菌群落的Beta多样性存在差异,但不同组间差异不显著(图3)。
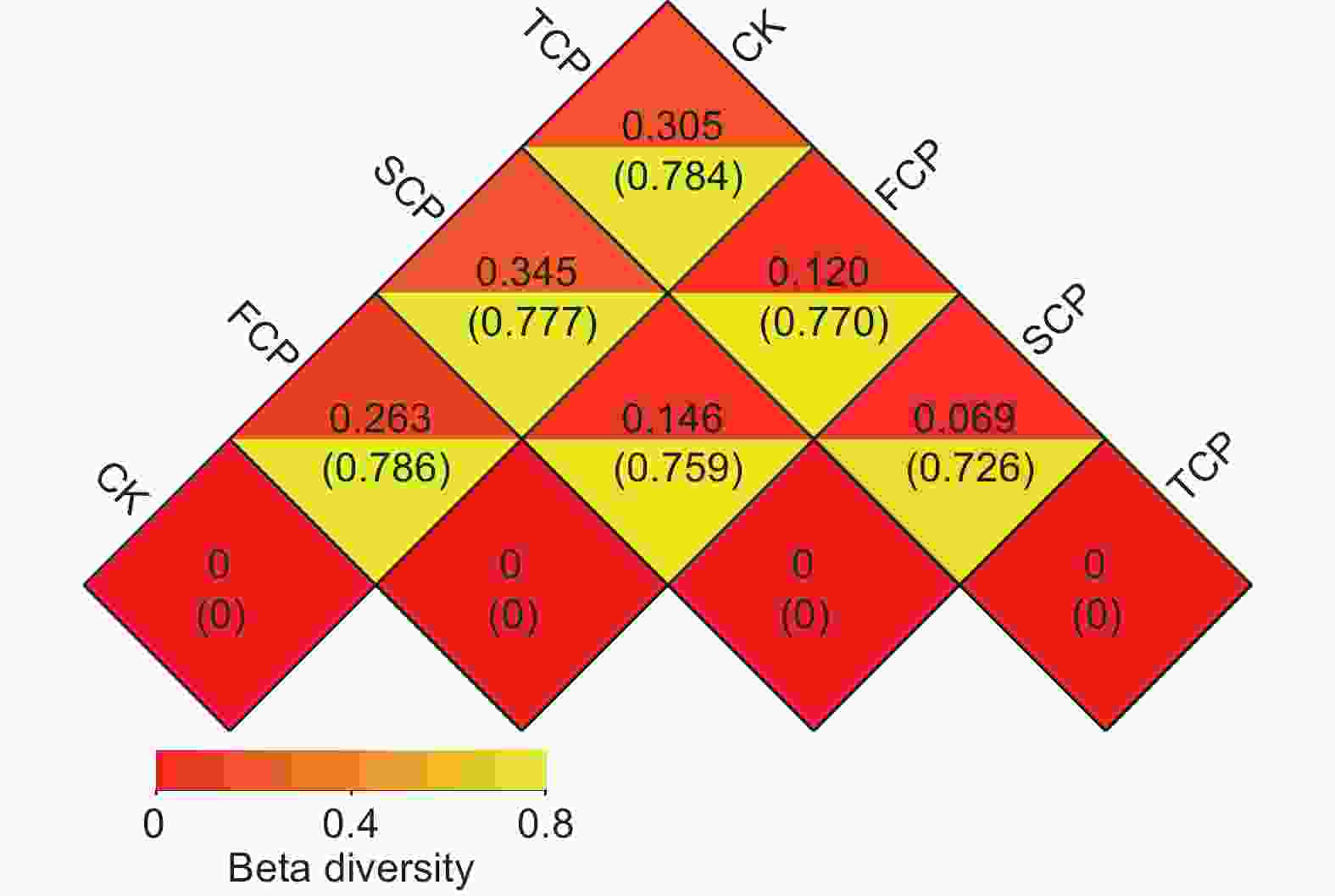
图 3 不同代数木麻黄根际土壤样本间的Weighted Unifrac距离和Unweighted Unifrac距离
Figure 3. Weighted and unweighted Unifrac distances between rhizosphere soils samples of different rotations of C. equisetifolia
-
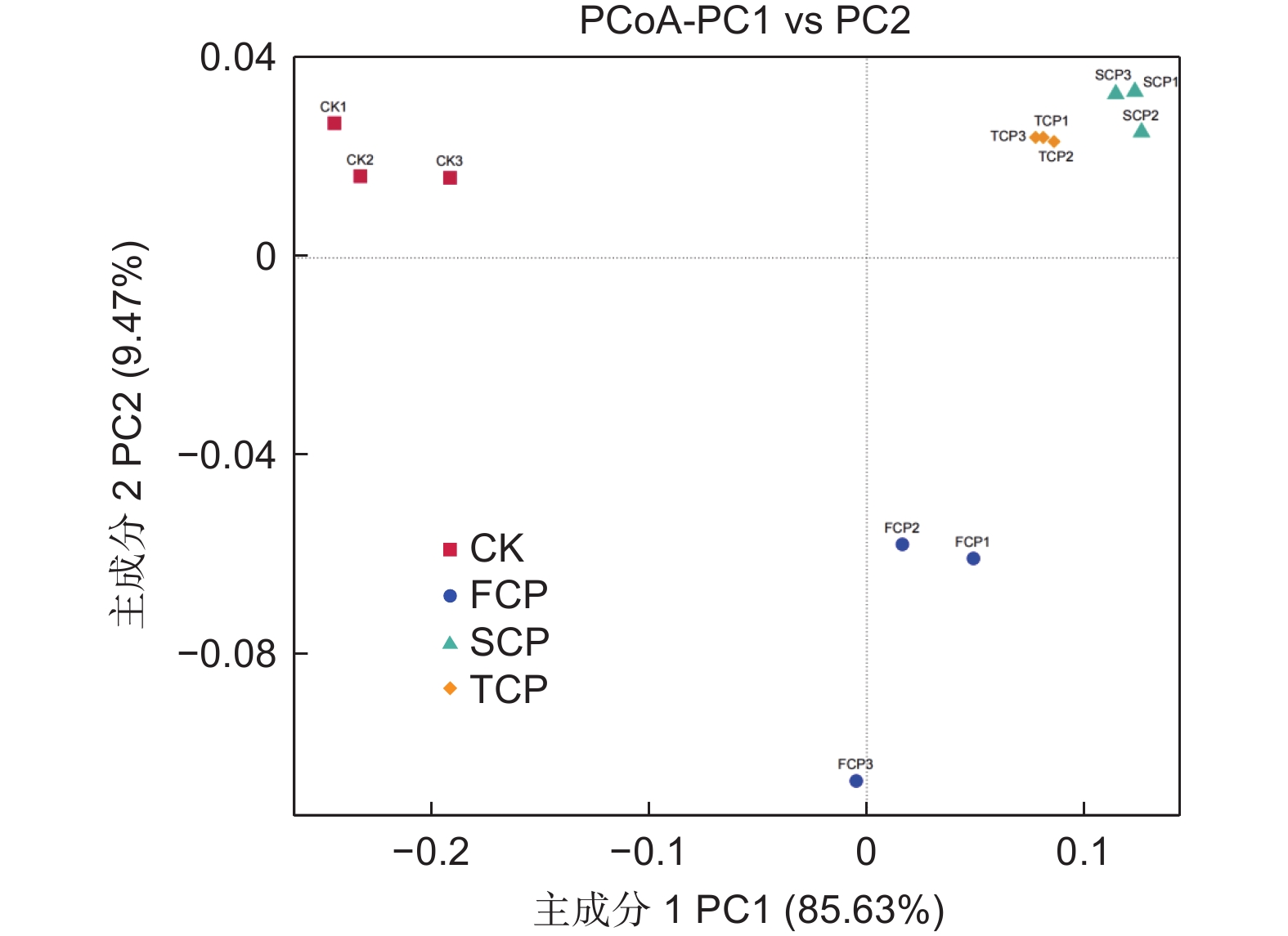
基于Weighted unifrac距离和Unweighted unifrac距离对连栽木麻黄根际土壤古菌群落进行主坐标分析(PCoA,Principal Co-ordinates Analysis)。结果表明,同组的3个平行样本彼此之间距离相近,说明样本重复性较好。分析结果显示,主成分1(PC1)解释变量方差的85.63%,主成分2(PC2)解释变量方差的9.47%,两者累计贡献率达95.10%,主成分1和主成分2有效将CK、FCP、SCP和TCP区别开来(图4)。通过Weighted Unifrac距离矩阵对不同土壤古菌群落进行UPGMA聚类分析,结果显示,SCP和TCP的古菌群落结构聚集为一个群体,两者再与FCP聚在一起,最后与CK形成系统发生树(图5)。进一步通过Anosim分析方法分析连栽木麻黄根际土壤古菌组间群落结构差异性是否显著。分析结果显示,不同组间的R值均大于0,表明组间差异大于组内差异,但是组间差异均不显著(P>0.05)(表4)。
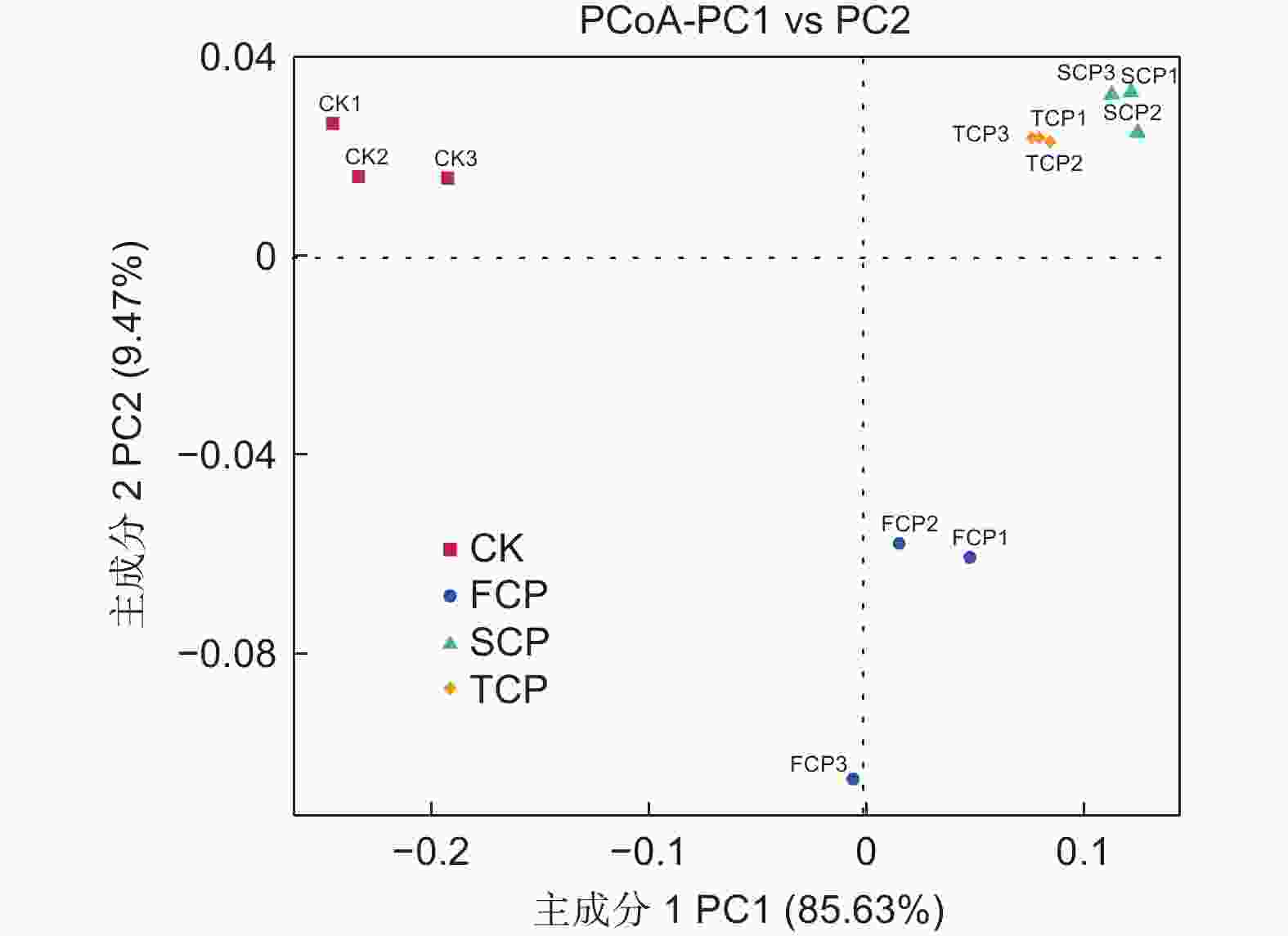
图 4 连栽木麻黄根际土壤古菌群落PcoA分析
Figure 4. PcoA analysis of archaeal communities from rhizosphere soils of continuous rotations of C. equisetifolia
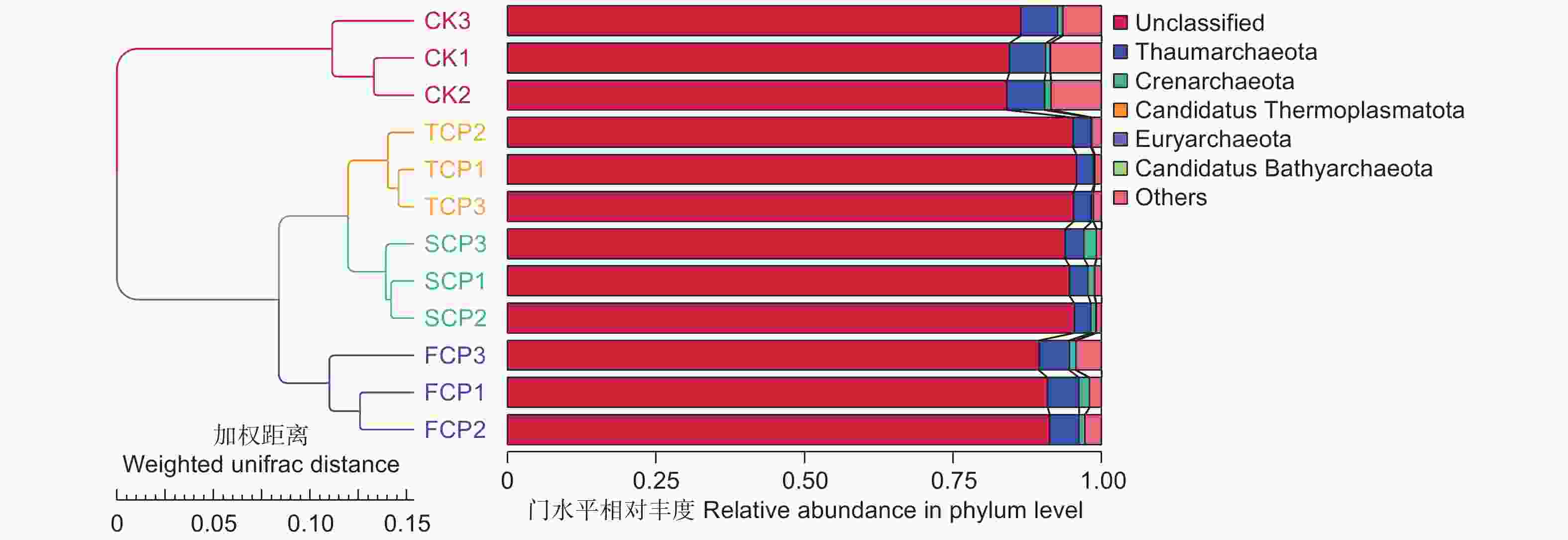
图 5 连栽木麻黄根际土壤古菌群落UPGMA聚类分析
Figure 5. UPGMA analysis of archaeal communities from rhizosphere soils of continuous rotations of C. equisetifolia
表 4 连栽木麻黄根际土壤古菌群落Anosim组间差异分析
Table 4. Analysis of Anosim group differences of archaeal communities in rhizosphere soils of continuous rotations of C. equisetifolia
组间
GroupR值
R-valueP值
P-valueCK-FCP 1 0.1 CK-SCP 1 0.1 CK-TCP 1 0.1 FCP-SCP 1 0.1 FCP-TCP 1 0.1 SCP-TCP 1 0.1 -
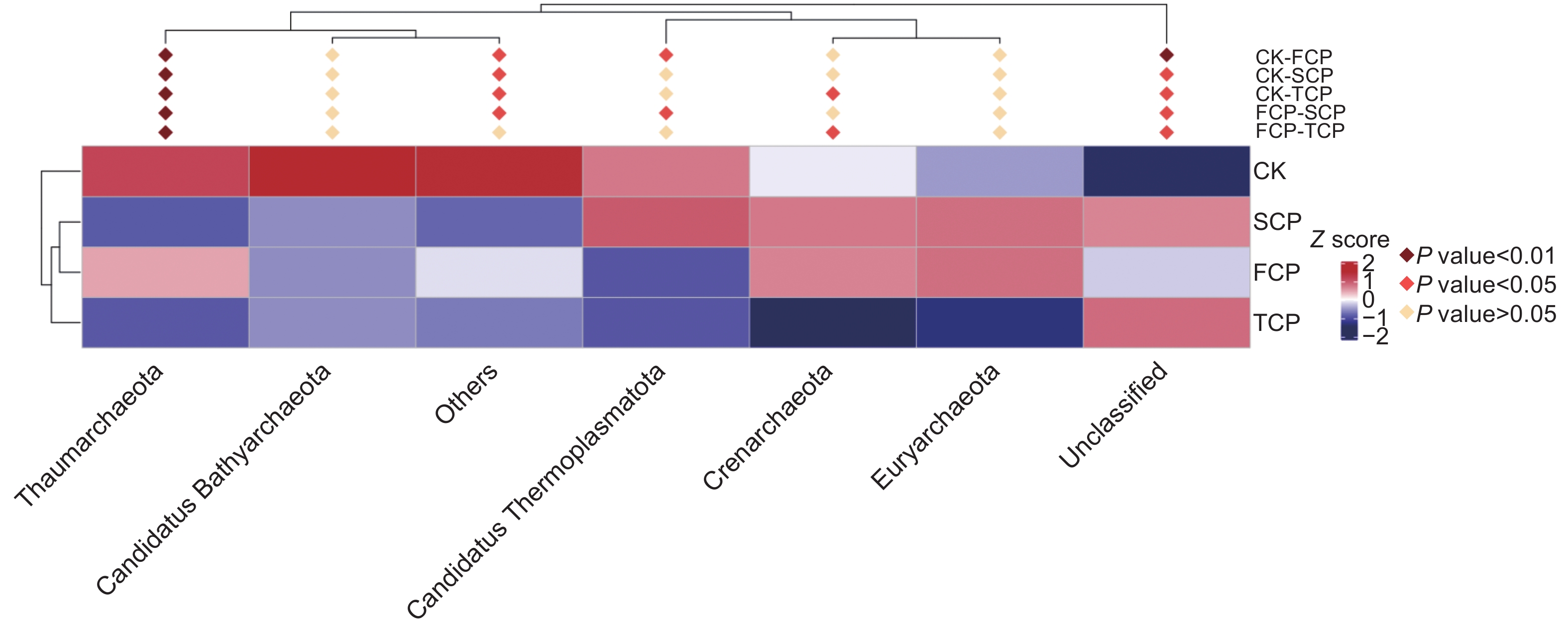
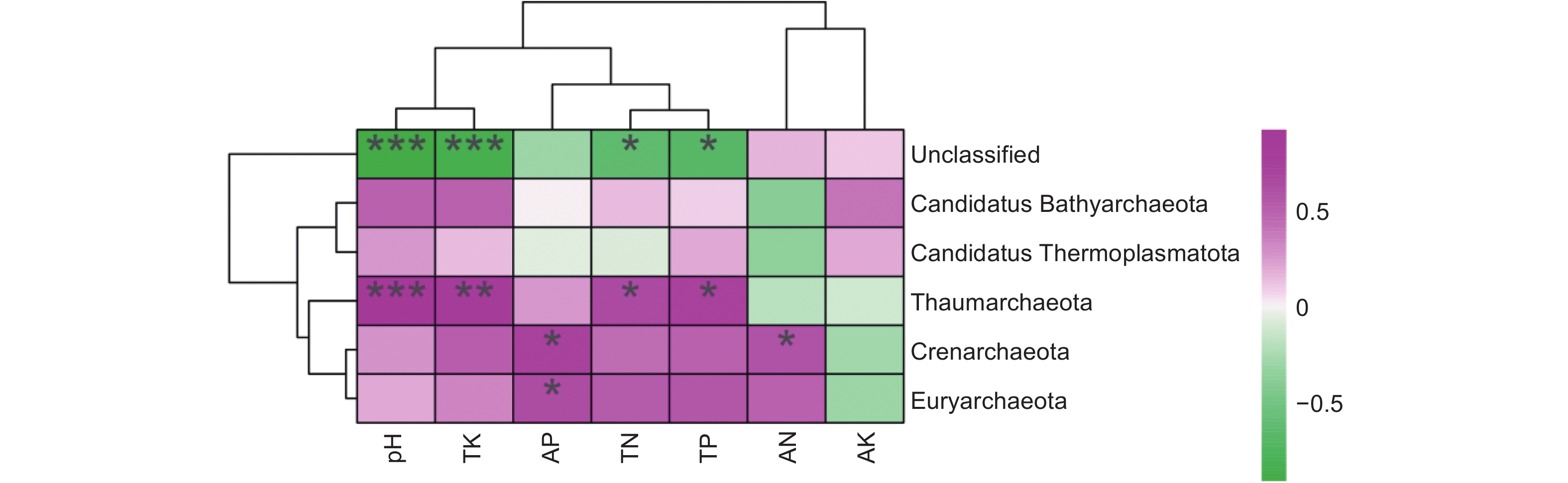
利用MetaStat方法进一步统计分析,有针对性地找出组间丰度变化差异显著的物种,并得到差异物种在不同分组内的富集情况,绘制热图。结果显示,奇古菌门(Thaumarchaeota)的物种差异在不同组间均达到了极显著水平(P<0.01),表明连栽木麻黄根际土壤奇古菌门的古菌发生了显著变化,泉古菌门(Crenarchaeota)在CK-TCP、FCP-TCP两个组间存在显著差异(P<0.05),Candidatus Thermoplasmatota在CK-FCP、FCP-SCP两个组间存在显著差异(P<0.05),广古菌门(Euryarchaeota)和Candidatus Bathyarchaeota在不同组间均未达到显著水平(图6)。
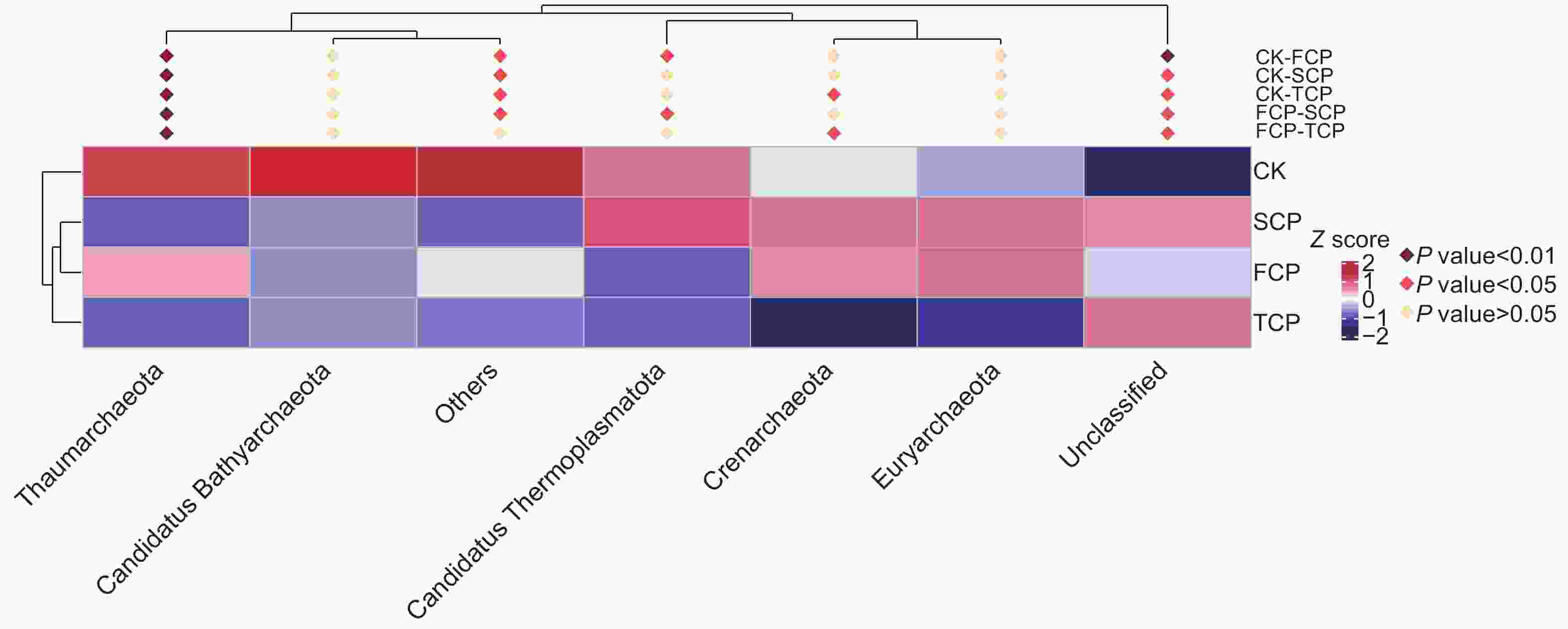
图 6 基于MetaStat方法的连栽木麻黄根际土壤古菌群落注释热图
Figure 6. Annotation heat map of archaeal communities in rhizosphere soils of continuous rotations of C. equisetifolia based on the MetaStat method
-
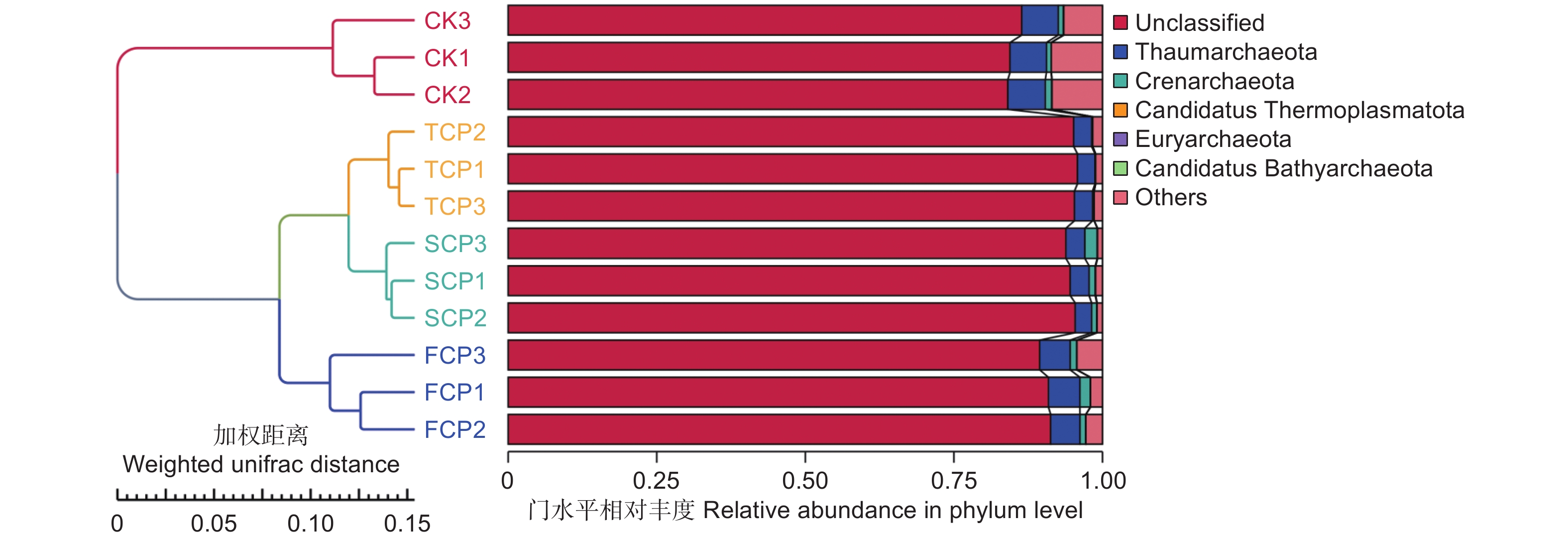
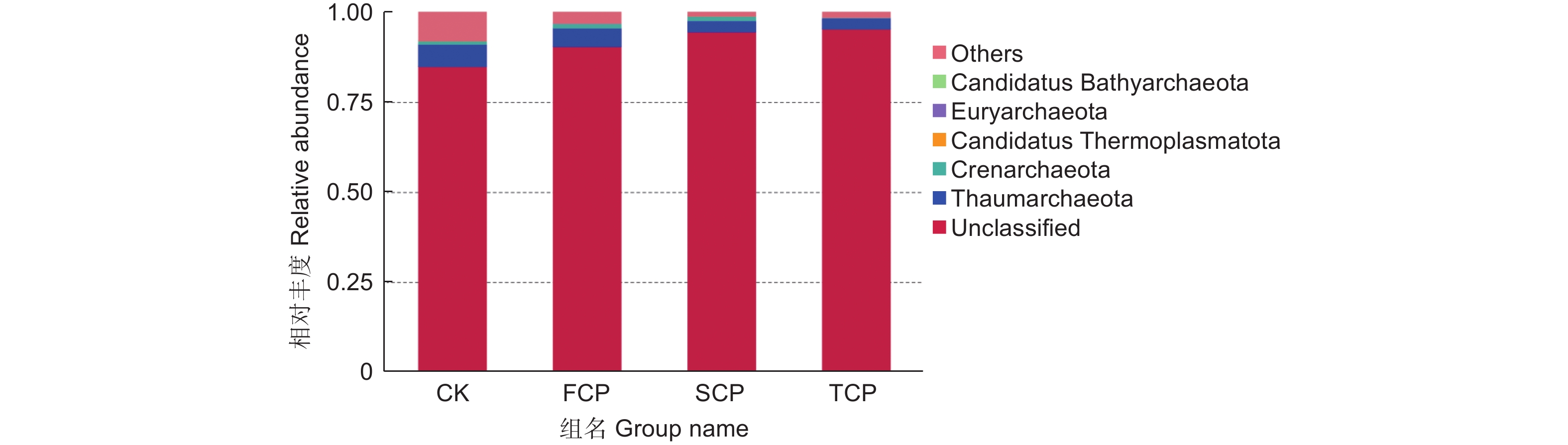
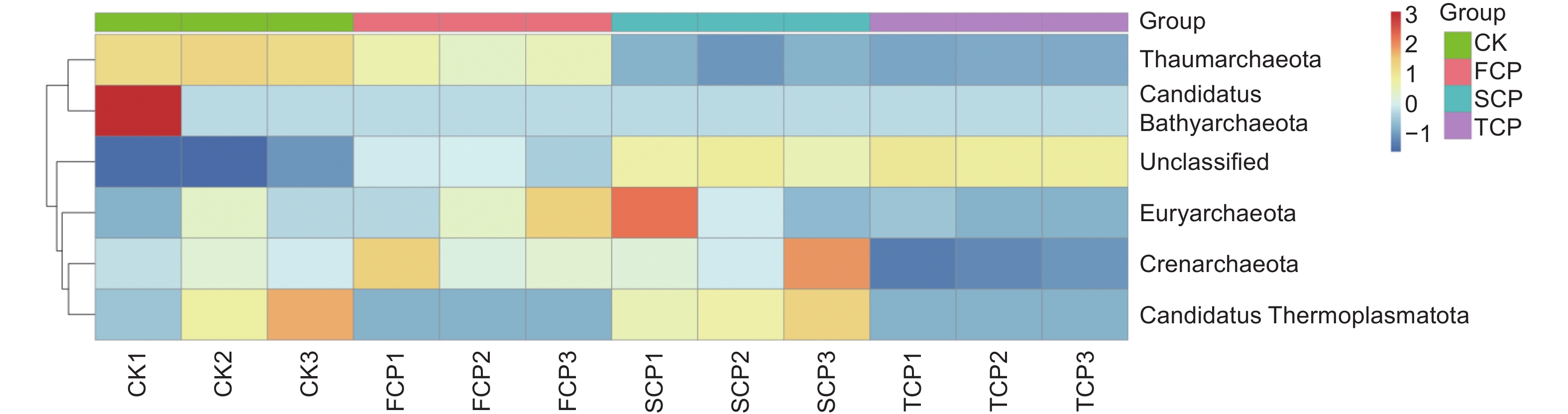
在不同连栽代数木麻黄根际土壤古菌群落中共检测到5个门,分别为奇古菌门(Thaumarchaeota)、泉古菌门(Crenarchaeota)、Candidatus Thermoplasmatota、广古菌门(Euryarchaeota)和Candidatus Bathyarchaeota。优势菌门为奇古菌门,占比3.00%-6.19%;次优势菌门为泉古菌门,占比0.19%-1.33%(图7)。除此之外,大部分的古菌是未分类的,占比高达84.07%-95.80%,说明木麻黄根际土壤古菌群落在很大程度上是未知的。进一步分析发现,奇古菌门占比随着栽植代数的增加呈现下降的趋势,与对照相比,FCP、SCP、TCP分别下降了17.42%、50.97%、51.51%,且除SCP和TCP之间未达到显著水平外,其余不同组分之间均存在显著差异(P<0.05),而泉古菌门占比随着栽植代数的增加呈现先上升后下降的趋势,FCP与SCP之间无显著差异,FCP、SCP均与TCP之间存在显著差异(P<0.05)。门水平上的群落热图分析也表明,不同连栽代数木麻黄根际土壤的古菌群落结构发生了显著变化(图8)。
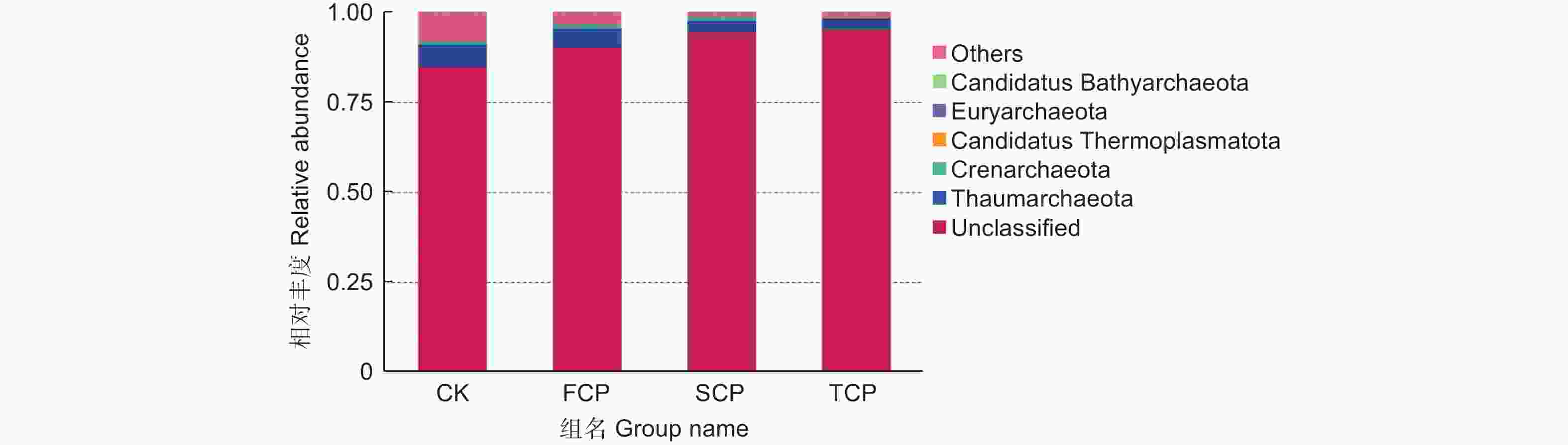
图 7 连栽木麻黄根际土壤优势古菌在门水平的群落相对丰度
Figure 7. The relative abundance of predominant archaea in rhizospheric soil samples of continuous rotations of C. equisetifolia at the phylum level
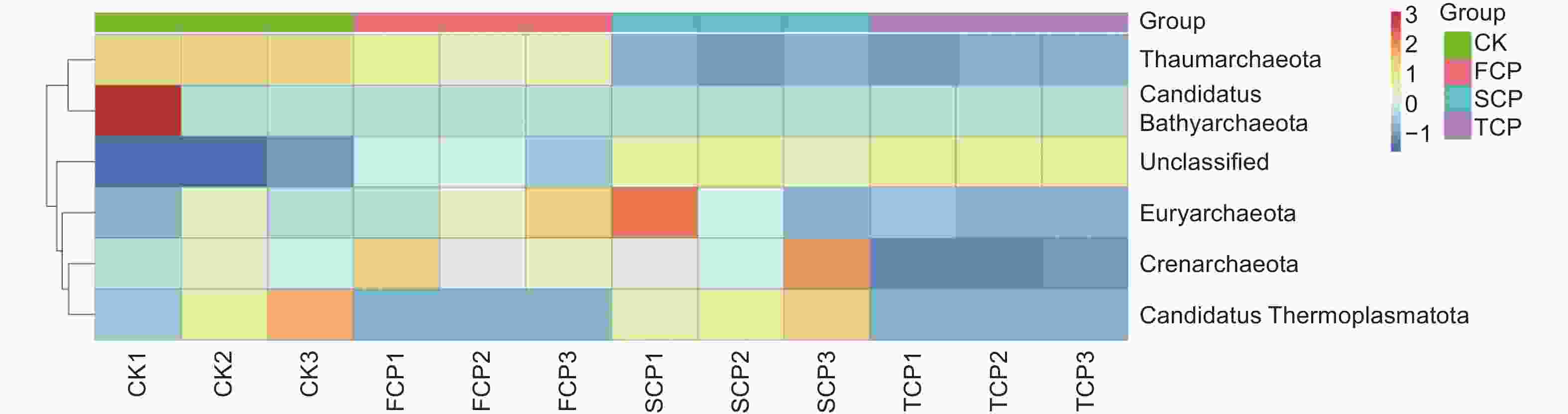
图 8 连栽木麻黄根际土壤古菌在门水平的群落热图分析
Figure 8. Heat map analysis of archaea in rhizospheric soil of continuous rotations of C. equisetifolia at the phylum level
-
为了更好地了解不同连栽代数的木麻黄根际土壤古菌群落与土壤理化因子的关系,对古菌群落与土壤理化因子进行了相关性分析(图9)。分析结果显示,优势菌门奇古菌门(Thaumarchaeota)与pH和TK呈极显著正相关,与TN和TP呈显著正相关,次优势菌门泉古菌门(Crenarchaeota)与AP和AN呈显著正相关,广古菌门(Euryarchaeota)与AP呈显著正相关,Candidatus Thermoplasmatota和Candidatus Bathyarchaeota与各土壤理化因子之间均无显著相关性,而Unclassified古菌与pH和TK呈极显著负相关,与TN和TP呈显著负相关,表明不同连栽代数的木麻黄根际土壤古菌群落与土壤理化性质具有密切的联系,而不同古菌群落与土壤理化因子之间的相关性存在显著差异。
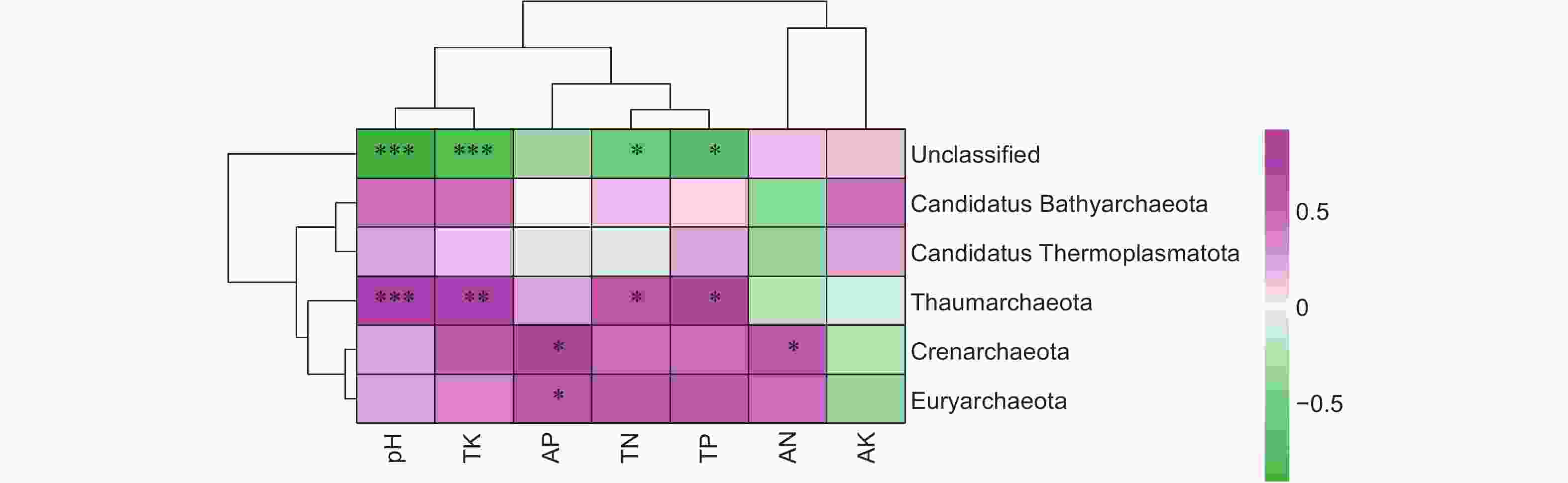
图 9 古菌群落与土壤理化因子相关性分析
Figure 9. Correlation analysis between archaeal communities and soil physicochemical factors
-
根际微生物在维持土壤生产力方面占有重要地位,促进土壤生态系统中的能量流动、物质循环和信息传递[26]。古菌作为地球进化最早期的生命体之一,分布的生态类型多样,在极端环境和非极端环境中均有分布,驱动着土壤中C、N、S、Fe等营养元素的生物地球化学循环[27-28]。由于古菌的培养条件比较特殊,对该群落结构生态功能的认识受到了极大限制。近年来,扩增子高通量测序技术成为了研究古菌群落结构的重要手段[29]。本研究对连栽木麻黄人工林根际土壤古菌群落进行16S rDNA扩增子高通量测序,发现随着木麻黄人工林连栽代数的增加,特异ASV数量呈现下降的趋势,根际土壤古菌群落所观察到的物种数和Chao1指数均呈现下降的趋势,Shannon指数、Simpson指数和pielou_e均以FCP最高,呈现先下降后上升的趋势,表明不同代数木麻黄根际土壤古菌群落的丰富度、多样性和均匀度发生了明显变化,PCoA分析、UPGMA聚类分析和Anosim分析显示根际土壤古菌群落存在差异,但不同组间差异不显著。推测原因可能是在连栽过程中土壤理化性质、林木生长发育、化感自毒物质、森林凋落物降解等因素存在差异所导致的。有研究表明,天然林向杉木人工林的转变导致土壤古菌的多样性和丰富度显著下降,并发现杉木人工林土壤中古菌的减少可能与土壤中氨离子浓度的变化有关[30]。除此之外,连栽杉木土壤中氨氧化古菌的丰度与多样性大致呈现逐代降低趋势,杉木多代连栽抑制了氨氧化古菌的生长,并证实了土壤养分含量与氨氧化古菌群落之间联系密切,氨氧化古菌对杉木人工林土壤氮素循环硝化过程起着十分重要的作用[31]。唐楚珺等[32]发现不同连栽代数杉木林土壤氨氧化古菌的基因丰度随连栽代数增加呈显著递减趋势。这表明连栽对人工林土壤古菌群落产生了显著影响,且在不同种类的人工林中普遍存在此现象。林木根系的生长发育也可能是造成人工林古菌群落结构存在差异的原因之一。朱启良等[33]对杨树人工林细根不同生长时期根际土壤进行古菌高通量测序分析,发现不同生长阶段细根根际土壤的古菌群落结构存在显著差异,且随着细根生长发育,占绝对优势的氨氧化古菌Candidatus_Nitrososphaera在根际土壤中的丰度呈现上升趋势,推测其可能与细根的生长发育具有密切的关系。此外,作物的连作同样对土壤古菌群落结构产生影响。张伟等[34]研究探讨了在新疆干旱半干旱的土壤环境下,棉花连作对土壤古菌群落结构的影响,结果表明在棉花长期连作过程中,促使形成了新的古菌群落结构组成,且棉花和玉米轮作能快速调整一些古菌属的丰度。这与连栽人工林的结果不一致,推测可能是物种差异、土壤异质性、地域差别等因素造成的。
-
土壤环境中包含着丰富的古菌资源,优势物种是微生物群落的一个重要特征,已有研究报道奇古菌门、广古菌门、泉古菌门为主要的优势菌门[35-39]。在本研究中,不同连栽代数木麻黄根际土壤和未种植木麻黄的对照土壤的古菌群落优势菌门均为奇古菌门。这与De Chaves等[40]研究结果一致。De Chaves等[40]收集了亚马逊原始森林、次生林、农业系统和牛牧场中的土壤样本,发现原始森林和次生林中的古菌群落以奇古菌类为主,且与次生林相比,原始森林中具有更多的古菌种类。Isoda等[41]比较了芬兰北方森林和日本寒温带森林之间矿质土壤中的古菌群落,发现在芬兰北方森林土壤中,奇古菌门占主导地位。Shi等[42]对青藏高原东部94个土壤样本的古菌群落进行了表征,发现奇古菌门是所有土壤中占优势的古菌门。以往的研究表明,奇古菌门是浅层土壤中最主要的古菌群落[43],本研究所取的土壤样品为根际土壤,得到了相似的结果。在本研究中,随着木麻黄连栽代数的增加,与对照相比,奇古菌门的占比分别下降了17.42%、50.97%、51.51%,奇古菌门的物种差异在不同组间均达到了极显著水平(P<0.01),说明奇古菌门在木麻黄连栽过程中发挥着重要作用。与不同代数的木麻黄人工林相比,作为对照的天然乔木林中含有更丰富的奇古菌门古菌,这与Pedrinho等人[44]的研究一致。然而,Tsang等[37]研究发现四种健康和被褐根病菌感染的树木(榕树(Ficus microcarpa Linn. f.)、朴树(Celtis sinensis Pers.)、白楸(Mallotus paniculatus (Lam.) Muell. Arg.)和樟树(Cinnamomum Camphora (L.) Presl.))中的优势古菌门为泉古菌门和广古菌门。殷萌清等[45]比较了天然及人工红树林土壤微生物的群落结构差异,发现相比于细菌,古菌的多样性明显偏低,所检测到的古菌在门的水平上主要为广古菌门。土壤类型的不同、土壤理化性质的差异、土壤深度以及植物根系空间距离,可能是影响土壤古菌优势类群的重要因素[46]。此外,本研究所测得的古菌群落大多数是未分类的,意味着对于古菌资源的挖掘需要进一步研究。
-
根际土壤微生物与土壤中的养分之间关系密切,彼此促进或制约,从而对植物的生长产生影响[47]。土壤理化因子的改变会干扰土壤微生物群落的变化,进而影响微生物的活性和生态功能[48]。本研究发现,不同连栽代数的木麻黄根际土壤古菌群落的不同门水平菌落与土壤理化因子之间的相关性存在显著差异,优势菌门奇古菌门与pH、TK、TN、TP呈显著正相关,次优势菌门泉古菌门与AP和AN呈显著正相关,结果表明土壤理化因子与古菌群落关系密切,这与Zhang等[49]的研究结果相似。陈雯雯等[50]发现连栽杉木土壤中NH4 + -N含量与奇古菌门呈正相关,与广古菌门呈负相关。因此,人工林土壤中古菌的减少可能与森林类型转换后土壤中氨离子浓度的变化有关[51]。也有证据表明,土壤NH4 + -N含量对氨氧化古菌的奇古菌门和广古菌门有直接影响作用,且NH4 + -N含量变化与之存在密切关系[52]。在本研究中,优势菌门奇古菌门与pH呈极显著正相关关系(P<0.001)。pH是影响土壤古菌群落结构的关键决定因素[53]。Zhou等[54]发现pH值是影响红树林和潮间带湿地泥滩中的分层古菌群落形成的最重要因素。在大兴安岭天然林演替中,pH对土壤古菌群落组成几乎没有影响,而在人工林生长过程中,pH对土壤古菌群落组成有影响[55]。在中国东北黑土区,古菌群落在具有明显的生物地理分布模式,土壤pH值是构建群落组成的关键土壤因素[56]。pH同样是影响土壤细菌群落结构的决定因素。Wei等[57]分析了太湖地区落叶常绿林根际细菌群落结构及多样性,发现细菌群落组成和多样性在很大程度上受土壤pH值和树种的影响,土壤pH值是影响细菌多样性的关键因素,较低的pH值与较少的群落多样性相关。综上,土壤理化因子的不同是导致古菌群落差异的重要原因,在人工林连栽过程中,土壤理化指标发生改变,进而影响根际土壤古菌群落。
-
本研究通过16S rDNA扩增子高通量测序研究连栽条件下木麻黄人工林根际土壤古菌群落变化规律,结果发现木麻黄人工林经过多代连续栽植后,根际土壤的古菌群落结构发生显著变化,优势菌门奇古菌门占比显著下降,推测古菌群落结构失衡是导致土壤微生态失衡的重要原因之一。因此,土壤微生态失衡可能是导致木麻黄人工林连栽障碍问题的关键原因。然而,连栽障碍的形成和加重是一个复杂的过程,不是由单一或孤立的因素所导致的,而是由植物、土壤和微生物系统内多种胁迫因子相互关联和相互影响的综合结果产生的。因此,今后从植物—土壤—微生物相互作用的根际对话过程为切入点,锁定关键功能古菌微生物,深入揭示木麻黄连栽障碍形成机理,阐明连栽木麻黄根际土壤微生态失衡规律,对于缓解连栽障碍难题具有重要意义。
连栽对木麻黄人工林根际土壤古菌群落变化的影响
Effect of Continuous Planting on Archaeal Community Changes in Rhizosphere of Casuarina equisetifolia Plantations
-
摘要:
目的 探究连栽对木麻黄人工林根际土壤古菌群落变化的影响,为探明木麻黄连栽障碍机制提供基础理论依据。 方法 以3个不同连栽代数(第一代First continuous plantation,FCP;第二代Second continuous plantation,SCP;第三代Third continuous plantation,TCP)的木麻黄人工林根际土壤为材料,以未种植木麻黄的天然乔木林根际土壤作为对照(CK),通过Illumina NovaSeq测序平台对古菌群落进行高通量测序。 结果 从3个不同连栽代数木麻黄人工林根际土壤和对照土壤中总共得到1 146 390条有效序列,以100%序列相似度聚类得到998 515个ASV。Alpha多样性分析结果表明,所观察到的物种数和Chao1指数均随着连栽代数的增加呈现下降的趋势,而Shannon指数、Simpson指数和pielou_e均以FCP最高,TCP次之,SCP最低。Beta多样性分析、PCoA分析、UPGMA聚类分析和Anosim分析结果表明,不同连栽代数木麻黄根际土壤古菌群落物种多样性存在差异,但不同组间差异不显著。共检测到5个门,优势菌门为奇古菌门(Thaumarchaeota),且奇古菌门占比随着栽植代数的增加呈现下降的趋势,与对照相比,FCP、SCP、TCP分别下降了17.42%、50.97%、51.51%。古菌群落与土壤理化性质相关性分析结果显示,奇古菌门与pH、全钾(TK)、全氮(TN)、全磷(TP)呈显著正相关,泉古菌门与速效磷(AP)和速效氮(AN)呈显著正相关,不同古菌群落与土壤理化因子之间的相关性存在显著差异。 结论 本研究证实了木麻黄人工林经过多代连续栽植后,根际土壤古菌群落结构发生显著变化,推测古菌群落结构失衡是导致土壤微生态失衡的重要原因之一。研究结果为揭示木麻黄人工林连栽障碍形成的原因提供了理论依据。 Abstract:Objective To investigate the impact of continuous planting on the changes in the archaeal community in the rhizosphere soil of Casuarina equisetifolia Forst. plantations for understanding of the obstacle mechanism of continuous planting of C. equisetifolia and providing a solid theoretical basis for further exploration. Methods We collected the rhizosphere soil from three C. equisetifolia plantations with varying generations of continuous planting (FCP, SCP, TCP). The rhizosphere soil from a natural arbor forest without C. equisetifolia was used as the control (CK). High-throughput sequencing of the archaeal community was performed using the Illumina NovaSeq sequencing platform. Results A total of 1 146 390 effective sequences were obtained from the rhizosphere soil samples and bare land control soil, resulting in the identification of 998 515 ASVs with 100% sequence similarity. The analysis of Alpha diversity indicated that the observed number of species and Chao1 index decreased with successive generations. However, the Shannon index, Simpson index, and Pielou_e were highest in FCP, followed by TCP, and lowest in SCP. The results of the Beta diversity analysis, Principal Coordinate Analysis (PCoA), UPGMA cluster analysis, and Anosim analysis indicated variations in the species diversity of the archaeal community in the rhizosphere soil of C. equisetifolia across different continuous planting generations. However, the differences between different groups were not significant. A total of 5 phyla were detected, and the dominant bacterial phylum was Thaumarchaeota and the proportion of Thaumarchaeota showed a decreasing trend with the increase of planting generations. Compared with the control, FCP, SCP and TCP decreased by 17.42%, 50.97% and 51.51% respectively. The results of the correlation analysis between archaea community and soil physical and chemical properties revealed significant positive correlations between Thaumarchaeota and pH, TK, TN and TP. Additionally, Crenarchaeota showed significant correlations with AP and AN. There were significant differences in the correlations between different archaeal communities and soil physical and chemical factors. Conclusion This study confirms that the archaeal community structure of the rhizosphere soil change significantly after multiple generations of continuous planting in C. equisetifolia plantations. It is speculated that the imbalance of the archaeal community structure is one of the key factors contributing to the imbalance of soil microecology. The study provides a theoretical foundation for understanding the causes of continuous planting obstacles in C. equisetifolia plantations. -
图 1 木麻黄根际土壤古菌群落稀释曲线
Figure 1. Rarefaction curves of the archaeal communities in rhizosphere soil of C. equisetifolia
图 2 基于ASVs丰度的木麻黄根际土壤古菌群落花瓣图
Figure 2. Petal plot of archaeal communities in rhizosphere soil of C. equisetifolia based on ASVs abundance
图 3 不同代数木麻黄根际土壤样本间的Weighted Unifrac距离和Unweighted Unifrac距离
Figure 3. Weighted and unweighted Unifrac distances between rhizosphere soils samples of different rotations of C. equisetifolia
图 4 连栽木麻黄根际土壤古菌群落PcoA分析
Figure 4. PcoA analysis of archaeal communities from rhizosphere soils of continuous rotations of C. equisetifolia
图 5 连栽木麻黄根际土壤古菌群落UPGMA聚类分析
Figure 5. UPGMA analysis of archaeal communities from rhizosphere soils of continuous rotations of C. equisetifolia
图 6 基于MetaStat方法的连栽木麻黄根际土壤古菌群落注释热图
Figure 6. Annotation heat map of archaeal communities in rhizosphere soils of continuous rotations of C. equisetifolia based on the MetaStat method
图 7 连栽木麻黄根际土壤优势古菌在门水平的群落相对丰度
Figure 7. The relative abundance of predominant archaea in rhizospheric soil samples of continuous rotations of C. equisetifolia at the phylum level
图 8 连栽木麻黄根际土壤古菌在门水平的群落热图分析
Figure 8. Heat map analysis of archaea in rhizospheric soil of continuous rotations of C. equisetifolia at the phylum level
图 9 古菌群落与土壤理化因子相关性分析
Figure 9. Correlation analysis between archaeal communities and soil physicochemical factors
表 1 木麻黄人工林样地基本情况
Table 1. Basic information of the C. equisetifolia plantation sample plots
样地
Sample plot栽植时间
Planting time方位
Position伴生树种
Associated tree species林下植被
Underforest vegetation土壤质地Soil texture CK — 118°90′43″ E,24°91′38″ N 台湾栾树(Koelreuteria elegans (Seem.) A. C. Smith ssp. formosana (Hayata) Meyer)和潺槁木姜子(Litsea glutinosa (Lour.) C. B. Rob.) 霍香蓟(Ageratum conyzoides L.)和鬼针草(Bidens pilosa L.) 风积黄沙土 FCP 1987年 118°90′52″ E,24°91′45″ N SCP 2011年 118°90′56″ E,24°91′47″ N TCP 2014年 118°90′31″ E,24°91′56″ N 注:CK:对照;FCP:一代林;SCP:二代林;TCP:三代林。下同。
Note: CK: Control; FCP: First continuous plantation; SCP: Second continuous plantation; TCP: Third continuous plantation. The same below. 下载: 导出CSV
下载: 导出CSV
表 2 研究区样地土壤基本理化指标
Table 2. Basic physical and chemical indexes of soil in the study area
指标Index CK FCP SCP TCP pH 5.17 ± 0.00a 5.15 ± 0.00a 4.94 ± 0.01b 4.93 ± 0.01b 全氮Total nitrogen(TN)(mg·g−1) 0.45 ± 0.01a 0.50 ± 0.02a 0.38 ± 0.02b 0.33 ± 0.02b 全磷Total phosphorus(TP)(mg·g−1) 0.39 ± 0.02ab 0.41 ± 0.00a 0.36 ± 0.01bc 0.33 ± 0.01c 全钾Total potassium(TK)(mg·g−1) 1.08 ± 0.05a 1.04 ± 0.02a 0.74 ± 0.03b 0.57 ± 0.03c 速效氮Alkaline nitrogen(AN)(mg·kg−1) 65.57 ± 0.28c 72.71 ± 0.52a 67.93 ± 0.24b 66.34 ± 0.30c 速效磷Available phosphorus(AP)(mg·kg−1) 6.39 ± 0.02c 6.70 ± 0.01a 6.45 ± 0.01b 6.26 ± 0.01d 速效钾Available potassium(AK)(mg·kg−1) 50.65 ± 0.78a 49.45 ± 0.29a 50.45 ± 0.11a 50.38 ± 0.19a 注:同行不同字母表示差异显著(P<0.05)。
Note: Different letters in the same line meant significant difference at 0.05 level.
下载: 导出CSV
表 3 连栽木麻黄根际土壤古菌群落丰富度和多样性指数
Table 3. Richness and diversity indices of archaeal communities in rhizosphere soils of continuous rotations of C. equisetifolia
土壤样品
Soil sample观察到的物种
Observed speciesChao1指数
Chao1 index香农指数
Shannon index辛普森指数
Simpson index均匀度指数
Pielou_eCK 243 ± 18a 247.341 ± 17.159a 4.789 ± 0.062a 0.905 ± 0.006a 0.605 ± 0.004a FCP 238 ± 31a 243.231 ± 31.191a 5.001 ± 0.043a 0.943 ± 0.001a 0.636 ± 0.011a SCP 208 ± 10a 221.002 ± 8.894a 3.226 ± 0.143c 0.695 ± 0.027c 0.419 ± 0.018c TCP 195 ± 18a 208.878 ± 23.102a 4.164 ± 0.040b 0.858 ± 0.004b 0.548 ± 0.012b 注:同列不同字母表示差异显著(P<0.05)。
Note: Different letters in the same column meant significant difference at 0.05 level.
下载: 导出CSV
表 4 连栽木麻黄根际土壤古菌群落Anosim组间差异分析
Table 4. Analysis of Anosim group differences of archaeal communities in rhizosphere soils of continuous rotations of C. equisetifolia
组间
GroupR值
R-valueP值
P-valueCK-FCP 1 0.1 CK-SCP 1 0.1 CK-TCP 1 0.1 FCP-SCP 1 0.1 FCP-TCP 1 0.1 SCP-TCP 1 0.1
下载: 导出CSV
-
[1] 魏永成, 张 勇, 孟景祥, 等. 不同种源短枝木麻黄对青枯病的生理生化响应及早期选择[J]. 林业科学,2021,57(11):134-141 doi: 10.11707/j.1001-7488.20211113 [2] CHANDARANA H, SENTHIL KUMAR P, SEENUVASAN M, et al. Kinetics, equilibrium and thermodynamic investigations of methylene blue dye removal using Casuarina equisetifolia pines[J]. Chemosphere,2021,285: 131480 doi: 10.1016/j.chemosphere.2021.131480 [3] 左林芝, 黄 蕊, 张雅倩, 等. 海南岛不同林龄短枝木麻黄凋落物内外真菌多样性分析[J]. 生态学报,2020,40(17):6086-6095 [4] 杨 彬, 郝清玉. 基于天然更新性能筛选海南岛木麻黄海防林混交树种[J]. 植物科学学报,2020,38(02):221-232 [5] PAPE IBRAHIMA D, NATHALIE D, MARIAMA N, et al. Selection of arbuscular mycorrhizal fungal strains to improve Casuarina equisetifolia L. and Casuarina glauca Sieb. tolerance to salinity[J]. Annals of Forest Science,2018,75(3): 72 doi: 10.1007/s13595-018-0747-1 [6] ZHOU X, WANG Y, Li C Q, et al. Differential expression pattern of pathogenicity-related genes of Ralstonia pseudosolanacearum responding to Tissue Debris of Casuarina equisetifolia[J]. Phytopathology,2021,111(11): 1918-1926 doi: 10.1094/PHYTO-11-20-0490-R [7] WU Z Y, LI J J, ZHENG J, et al. Soil microbial community structure and catabolic activity are significantly degenerated in successive rotations of Chinese fir plantations[J]. Scientific Reports,2017,7(1): 6691 doi: 10.1038/s41598-017-06768-x [8] CHEN D Z, YE G F, GAO W, et al. Ecological response of Casuarina equisetifolia to environmental stress in coastal dunes in China[J]. Journal of Forest Research,2018,23(3): 173-182 doi: 10.1080/13416979.2018.1469209 [9] 王玉. 海南岛木麻黄人工海防林天然更新困难的障碍机制[D]. 海南: 海南师范大学, 2020 [10] 叶功富, 张水松, 黄传英, 等. 木麻黄人工林地持续利用问题的探讨[J]. 林业科技开发,1994(04):18-19 [11] 叶功富, 徐俊森, 林武星, 等. 木麻黄连栽林地土壤肥力动态与地力维持[J]. 防护林科技,1996(S1):49-53 + 89 [12] KAŠTOVSKÁ E, CÁRDENAS-HERNÁNDEZ J, KUZYAKOV Y. Priming effects in the rhizosphere and root detritusphere of two wet-grassland graminoids[J]. Plant and Soil,2022,472(1-2): 105-126 doi: 10.1007/s11104-021-05191-6 [13] BENITEZ M S, EWING P M, OSBORNE S L, et al. Rhizosphere microbial communities explain positive effects of diverse crop rotations on maize and soybean performance[J]. Soil Biology & Biochemistry,2021,159: 108309 [14] BAKER B J, DE ANDA V, SEITZ K W, et al. Diversity, ecology and evolution of Archaea[J]. Nature Microbiology,2020,5(7): 887-900 doi: 10.1038/s41564-020-0715-z [15] 东秀珠, 李 猛, 向 华, 等. 探秘生命的第三种形式——我国古菌研究之回顾与展望[J]. 中国科学:生命科学,2019,49(11):1520-1542 [16] MOISSL-EICHINGER C, PAUSAN M, TAFFNER J, et al. Archaea are interactive components of complex microbiomes[J]. Trends in Microbiology,2018,26(1): 70-85 doi: 10.1016/j.tim.2017.07.004 [17] AZZIZ G, TRASANTE T, MONZA J, et al. The effect of soil type, rice cultivar and water management on ammonia-oxidizing archaea andbacteria populations[J]. Applied Soil Ecology,2016,100(1): 8-17 [18] CHEN X, ZHANG L M, SHEN J P, et al. Soil type determines the abundance and community structure of ammonia-oxidizingbacteria and archaea in flooded paddy soils[J]. Journal of Soils & Sediments,2010,10(8): 1510-1516 [19] ZHOU L T, LI J J, LUO Y, et al. Variation in soil fungal community structure during successive rotations of Casuarina equisetifolia plantations as determined by high-throughput sequencing analysis[J]. Plant Growth Regulation,2019,87(3): 445-453 doi: 10.1007/s10725-019-00483-5 [20] ZHOU L T, LI J J, POKHREL G R, et al. nifH gene sequencing reveals the effects of successive monoculture on soil diazotrophic microbial community in Casuarina equisetifolia plantations[J]. Frontiers in Plant Science, 2020, 11: 578812 [21] ZHOU L T, LI J J, POKHREL G R, et al. Effects of monoculture regime on the soil nirK- and nosZ-denitrifying bacterial communities of Casuarina equisetifolia[J]. Applied Soil Ecology,2022,171: 104326 doi: 10.1016/j.apsoil.2021.104326 [22] 吴则焰, 赵紫檀, 林文雄, 等. 基于T-RFLP方法的连栽杉木根际土壤细菌群落变化研究[J]. 生态学报,2019,39(19):1-11 [23] 郭清明. 滨海沙地3种相思生长防护效果及改土能力研究[J]. 安徽农学通报(上半月刊),2010,16(13):202-203 [24] 王 圳, 张金池, 崔晓晓, 等. VA菌根对喀斯特草本群落植物根际养分影响研究[J]. 生态环境学报,2010,19(7):1574-1577 [25] LI M J, SHAO D T, ZHOU J C, et al. Signatures within esophageal microbiota with progression of esophageal squamous cell carcinoma[J]. Chinese Journal of Cancer Research,2020,32(6): 755-767 doi: 10.21147/j.issn.1000-9604.2020.06.09 [26] 万 盼, 胡艳波, 张弓乔, 等. 甘肃小陇山油松与柴胡栽培土壤细菌群落特征[J]. 生态学报,2018,38(17):6016-6024 [27] ANGEL R, SOARES M I, UNGAR E D, et al. Biogeography of soil archaea and bacteria along a steep precipitation gradient[J]. The ISME Journal,2010,4(4): 553-563 doi: 10.1038/ismej.2009.136 [28] WANG H L, BIER R, ZGLESZEWSKI L, et al. Distinct distribution of archaea from soil to freshwater to estuary: implications of archaeal composition and function in different environments[J]. Frontiers in Microbiology,2020,11: 576661 doi: 10.3389/fmicb.2020.576661 [29] 李曙光, 皮昀丹, Zhang Chuan-lun. 古菌研究及其展望[J]. 中国科学技术大学学报,2007,37(08):830-838 [30] 郭佳欢. 杉木人工林土壤肥力及微生物群落结构和功能研究[D]. 南京林业大学, 2022 [31] 陈雯雯, 王淑真, 姜宇杰, 等. 杉木连栽氨氧化古菌群落结构与硝态氮含量的关系[J]. 林业科学研究,2023,36(02):79-90 [32] 唐楚珺, 高李文, 彭紫薇, 等. 连栽杉木人工林土壤氮循环功能基因丰度变化[J]. 应用与环境生物学报,2023,29(1):154-161 [33] 朱启良, 刘洪凯, 陈 旭, 等. 杨树人工林根际古菌群落随细根生长的演变[J]. 应用生态学报,2019,30(03):849-856 [34] 张 伟, 杜 钰. 棉花长期连作对新疆农田土壤古菌群落演替的影响[J]. 生态环境学报,2019,28(4):769-775 [35] NICOL G W, GLOVER L A, PROSSER J I. Spatial analysis of archaeal community structure in grassland soil[J]. Applied and Environmental Microbiology,2003,69(12): 7420-7429 doi: 10.1128/AEM.69.12.7420-7429.2003 [36] NICOL G W, TSCHERKO D, EMBLEY TM, et al. Primary succession of soil Crenarchaeota across a receding glacier foreland[J]. Environmental Microbiology,2005,7: 337-347 doi: 10.1111/j.1462-2920.2005.00698.x [37] TSANG K S W, CHEUNG M K, LAM R Y C, et al. A preliminary examination of the bacterial, archaeal, and fungal rhizosphere microbiome in healthy and Phellinus noxius-infected trees[J]. Microbiologyopen,2020,9(10): e1115 doi: 10.1002/mbo3.1115 [38] 戴瑞卿, 林明辉, 赖宝春, 等. 辣椒健康植株与患枯萎病植株根际土壤古菌群落多样性的比较研究[J]. 福建热作科技,2019,44(04):10-14 doi: 10.3969/j.issn.1006-2327.2019.04.004 [39] 闫慧贞, 徐邹洋, 陈文杰, 等. 环梅山岛海域春季浮游古菌群落空间分布特征研究[J]. 微生物学报,2021,61(9):2709-2725 [40] DE CHAVES M G, MERLOTI L F, DE SOUZA L F, et al. Ecological co-occurrence and soil physicochemical factors drive the archaeal community in Amazonian soils[J]. Archives of Microbiology,2022,205(1): 31 [41] ISODA R, HARA S, TAHVANAINEN T, et al. Comparison of archaeal communities in mineral soils at a boreal forest in Finland and a cold-temperate forest in Japan[J]. Microbes and Environments,2017,32(4): 390-393 doi: 10.1264/jsme2.ME17100 [42] SHI Y, ADAMS J M, NI Y Y, et al. The biogeography of soil archaeal communities on the eastern Tibetan Plateau[J]. Scientific Reports,2016,6: 38893 doi: 10.1038/srep38893 [43] MIKKONEN A, SANTALAHTI M, LAPPI K, et al. Bacterial and archaeal communities in long-term contaminated surface and subsurface soil evaluated through coextracted RNA and DNA[J]. FEMS Microbiology Ecology,2014,90(1): 103-114 doi: 10.1111/1574-6941.12376 [44] PEDRINHO A, MENDES L W, MERLOTI L F, et al. Forest-to-pasture conversion and recovery based on assessment of microbial communities in Eastern Amazon rainforest[J]. FEMS Microbiology Ecology,2019,95(3): fiy236 [45] 殷萌清, 冯建祥, 黄小芳, 等. 天然及人工红树林土壤微生物群落结构分析[J]. 生态科学,2017,36(5):1-10 [46] 刘建利, 王英娜, 未 丽, 等. 荒漠孑遗植物四合木对土壤古菌群落的影响[J]. 生态学报,2021,41(9):3548-3563 [47] 刘会芳, 韩宏伟, 王 强, 等. 不同蔬菜与番茄轮作对设施土壤微生物多样性、酶活性及土壤理化性质的影响[J]. 微生物学报,2021,61(01):167-182 [48] 李 娟, 赵秉强, 李秀英, 等. 长期不同施肥制度下几种土壤微生物学特征变化[J]. 植物生态学报,2008,32(4):891-899 [49] ZHANG M J, CHAI L W, HUANG M K, et al. Deciphering the archaeal communities in tree rhizosphere of the Qinghai-Tibetan plateau[J]. BMC Microbiology,2020,20(1): 235 doi: 10.1186/s12866-020-01913-5 [50] 陈雯雯, 王淑真, 姜宇杰, 等. 杉木连栽氨氧化古菌群落结构与硝态氮含量的关系[J]. 林业科学研究,2023,36(02):79-90 doi: 10.12403/j.1001-1498.20220369 [51] 郭佳欢. 杉木人工林土壤肥力及微生物群落结构和功能研究[D]. 南京林业大学, 2022 [52] JIAO S, XU Y Q, ZHANG J, et al. Environmental filtering drives distinct continental atlases of soil archaea between dryland and wetland agricultural ecosystems[J]. Microbiome,2019,7(1): 15 doi: 10.1186/s40168-019-0630-9 [53] TRIPATHI B M, KIM M, TATENO R, et al. Soil pH and biome are both key determinants of soil archaeal community structure[J]. Soil Biology and Biochemistry,2015,88: 1-8 doi: 10.1016/j.soilbio.2015.05.004 [54] ZHOU Z C, MENG H, LIU Y, et al. Stratified bacterial and archaeal community in mangrove and intertidal wetland mudflats revealed by high throughput 16S rRNA gene sequencing[J]. Frontiers in Microbiology,2017,8: 2148 doi: 10.3389/fmicb.2017.02148 [55] 牛晓燕. 大兴安岭天然林演替和人工林生长对于土壤微生物多样性动态的影响[D]. 内蒙古农业大学, 2018 [56] LIU J J, YU Z H, YAO Q, et al. Biogeographic distribution patterns of the archaeal communities across the black soil zone of northeast China[J]. Frontiers in Microbiology,2019,10: 23 doi: 10.3389/fmicb.2019.00023 [57] WEI Z W, HU X L, LI X H, et al. The rhizospheric microbial community structure and diversity of deciduous and evergreen forests in Taihu Lake area, China[J]. PloS One,2017,12(4): e0174411 doi: 10.1371/journal.pone.0174411 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 396
- HTML全文浏览量: 80
- PDF下载量: 2
- 被引次数: 0
