



-
光是影响植物生存和生长的重要环境因子,不同植物对光的利用能力不同,如阳生植物具有较强的光合同化能力、较高的光饱和点和光补偿点[1-2];而阴生植物适应弱光环境,具有较低的光补偿点和表观量子效率等特征[3]。植物对光的响应特征反映了植物对环境的适应能力,是决定其物种丰度和分布模式的重要因素。幼苗是树木更新的重要阶段,幼苗对不同光照条件的适应性对森林植被的恢复和重建具有重要影响[4]。我国栎属(Quercus L.)植物中的很多树种,如麻栎(Q. acutissima Carruth.)、白栎(Q. fabri Hance)和青冈栎(Cyclobalanopsis glauca (Thunb.) Oerst.)等是东部常绿落叶阔叶林的优势种或建群种,其中光照强度是影响其天然林更新的重要生态因子之一,国内很多学者在栎树对光照强度的响应方面作了大量研究[4-7],对阐明栎属植物天然林更新的机制提供了重要参考。
弗吉尼亚栎(Q. virginiana Mill.)原产美国,为壳斗科(Fagaceae)栎属常绿乔木,其根系深而发达,具有很强的抗风能力,同时能耐受干旱、高浓度盐雾和土壤盐分,是美国沿海硬阔林和灌丛林地的顶级优势树种[8-9]。本所于2000年将弗吉尼亚栎引种至长江中下游地区,在浙江慈溪和上虞以及上海、江苏等地沿海滩涂表现出很强的适应能力,成为该地区沿海防护林建设的重要树种[10-11]。弗吉尼亚栎种子无休眠期,落地后只要条件合适即萌发,然而在多年生人工林中,很难发现林下更新苗,而林下更新苗多见于林分外围地带,因此推测光环境可能是影响弗吉尼亚栎林下更新的重要因子。国外有研究认为[12-13],弗吉尼亚栎属于阳生植物,对光的需求较大,不具有耐荫性。Spector等[13]研究发现,林分的过分拥挤会导致弗吉尼亚栎生长所需的空间和光线减少,从而导致弗吉尼亚栎群落的衰退。同时也发现,成年弗吉尼亚栎遮荫叶片和向阳叶片的净光合速率和光补偿点并没有显著差异。弗吉尼亚栎在国内引种以来,研究多集中在繁育技术和抗逆性方面,发现弗吉尼亚栎对盐胁迫、高温、干旱等逆境均具有很强的耐受性[14-17],但其对光环境的响应并不清楚。因此,本研究采用人工遮荫方法,研究弗吉尼亚栎苗期在不同遮荫环境下的生长、光合作用以及叶绿素荧光响应,以期阐明弗吉尼亚栎引种后对光环境的适应能力,为弗吉尼亚栎的引种驯化和高效栽培提供科学依据。
-
试验采用弗吉尼亚栎2年生实生苗,种子来源于浙江上虞弗吉尼亚栎母树林。试验地点位于中国林业科学研究院亚热带林业研究所苗圃,地理位置为30.03′ N, 119.57′ E,该地块地处亚热带季风气候,夏季平均气温为26.5℃,降水量546.60 mm;冬季平均气温6.4℃,降水量339.60 mm。
-
采用盆栽实验方法,选取生长一致的苗木于2016年3月移栽于花盆(24 cm×16 cm),培养介质为60%黄泥土+30%泥炭+10%珍珠岩。试验采用随机区组设计,共3个处理:全光(FL)、50%自然光(中度遮荫,MS;以双层遮荫网覆盖)和25%自然光(强度遮荫,HS;以3层遮荫网覆盖),每个处理15株苗木作为1个区组,共3个区组,处理时间为2016年7—10月。试验期间,根据气温变化(图 1)定期浇水。试验前后测定苗木生长量,试验期间定期测量(共测定5次)叶绿素相对含量、叶绿素荧光参数的变化,并于试验结束时测定光合作用参数。

图 1 试验期间日均气温变化
Figure 1. The temperature changes during the experiment period
-
(1) 苗木生长测定:分别于试验前及收获苗木时测定苗高(H)和地径(D),并计算相对苗高(地径)生长率,计算公式如下:
相对生长率(%)=(试验后测量值-试验前测量值/试验前测量值)×100%
(2) 叶绿素含量指数(CCI)测定:采用CCM-200叶绿素测定仪(美国Opti-Sciences公司)测定叶片叶绿素含量指数[18]。每个处理随机选取5株苗木,测量从顶部起第510片叶,每个叶片分别在叶基部、中部和叶尖处避开叶脉选取3个测量点,最后取平均值。
(3) 光响应曲线测定:采用Licor-6400便携式光合测定系统(美国LI-COR公司)进行光响应曲线测定。安装红蓝光源,光照强度(PAR)梯度自高到低设定为2 000、1 800、1 600、1 400、1 200、1 000、800、600、400、200、100、50、20、0 μmol·m-2·s-1。在不同处理的各个重复中,选择长势最好的幼苗各3株,每株从第510片叶中选择3片功能叶作为测定叶,测定不同光照梯度下的叶片净光合速率(Pn),取3片功能叶的平均值作为单株光合作用的测量值。测定当天平均气温为27 ℃,相对湿度为70.5%,CO2浓度为300 μmol·mol-1,设定流速为500 μmol·s-1;测定前对叶片进行15 min光诱导(1 000 μmol·m-2·s-1),每个不同光照强度值设定数据采集时间3 min。根据光照强度及测得的相应净光合速率绘制Pn-PAR响应曲线。
光响应曲线模拟:采用叶子飘构建的光响应模型进行拟合[19-20],其模型表达式为:
$ Pn\left( I \right) = \alpha \frac{{1 - \beta I}}{{1 + \gamma I}}I - Rd $
其中:Pn为净光合速率(μmol·m-2·s-1),I为光量子通量密度,α为光响应曲线的初始斜率,即表观量子效率,β为修正系数,γ为光响应曲线初始斜率与最大净光合速率(Pmax)之比,Rd为暗呼吸速率(μmol·m-2·s-1)。光补偿点(LCP,μmol·m-2·s-1)和光饱和点(LSP,μmol·m-2·s-1)均基于以上模型进行估算。
(4) 叶绿素荧光参数测定:采用Mini-PAM调制叶绿素荧光测定仪(德国Walz公司)测定叶绿素荧光参数。叶片选取方法同(1),所有数据采用5个叶片的平均值作为单株测量值。测定前以锡箔纸包裹叶片进行暗适应30 min,测定叶绿素初始荧光(F0)和最大荧光(Fm);然后以强度156 μmol·m-2·s-1的光化光测定光下的叶绿素荧光参数,包括光下最大荧光(Fm’)和实时荧光(F),并打开红外光测定光适应下的初始荧光F0’,根据1/2计算以下叶绿素荧光相关参数:
可变荧光(Fv)= Fm-F0
光系统Ⅱ(PSⅡ)最大光化学量子产量(Fv/Fm)= (Fm-F0)/Fm
PSII实际量子产量(Yield)=△F/Fm’=(Fm’-F) / Fm’
光化学淬灭系数(qP)= (Fm’- F)/(Fm’ - F0’)
非光化学淬灭系数(NPQ)=1- (Fm’ - F0’)/(Fm-F0)
-
采用Microsoft Excel 2007、Origin 2016软件进行数据分析和图表处理,利用DPS13.0统计软件对数据进行方差分析及光响应拟合分析。
-
与全光条件相比,经过120 d的中度遮荫(50%自然光)处理后,弗吉尼亚栎苗木的苗高相对生长率明显增加(P < 0.05),而强度遮荫(25%自然光)处理下苗高相对生长率有所下降,但与全光处理下的差异不明显(表 1)。地径生长在遮荫处理下均有所下降,中度遮荫对地径生长的抑制不明显,然而强度遮荫条件下,地径生长明显受到抑制(P < 0.05)。由此可见,强度遮荫对弗吉尼亚栎苗高生长和地径生长均有抑制作用,尤其对地径生长的不利影响更为显著。
表 1 夏季遮阴对弗吉尼亚栎苗木生长的影响
Table 1. Effects of shading on growth of Q. virginiana
生长指标
Growth parameters处理
Treatments处理前
Before treatments处理后
After treatments相对生长率
Relative growth rate/%苗高Height/cm 全光FL 59.25±7.53 a 62.00±7.97 a 4.73±1.55 b 中度遮荫MS 57.75±10.08 a 61.50±7.61 a 7.64±0.29 a 强度遮荫HS 60.58±12.32 a 61.79±12.07 a 2.22±0.58 b 地径Basal diameter/cm 全光FL 3.99±0.37 a 4.96±0.47 a 24.40±3.06a 中度遮荫MS 3.73±0.59 a 4.46±0.62 a 19.90±2.08 a 强度遮荫HS 3.87±0.62 a 4.24±0.63 b 9.79±2.06 b 注:表中数据为平均值±标准差,小写字母不同代表不同光照强度之间差异显著(P < 0.05)。下同。
Notes: Values are means±sd, the different letters represent the significant difference among different light treatment(P < 0.05). The same as below. -
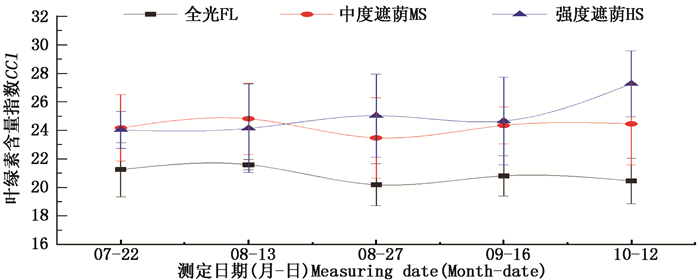
根据观测,全光条件下生长的弗吉尼亚栎苗木叶色淡绿,强度遮荫下(25%自然光)的弗吉尼亚栎叶色深绿。叶绿素含量指数(CCI)测定结果表明(图 2),全光处理与两种遮阴处理之间有明显差异。全光条件下弗吉尼亚栎CCI在2022之间波动,最终降低为20.45。50%自然光处理下,CCI在2426之间波动。25%自然光处理下,叶绿素含量指数最高,最终为27.26,明显高于其它处理。由此可见,遮荫处理对弗吉尼亚栎苗木的叶绿素合成有一定的促进作用,增强了弗吉尼亚栎叶片对光的捕获和吸收能力,是对光环境的一种生理适应,这在很多植物中均发现有类似的响应[2, 6, 23]。
图 2 不同光照处理下弗吉尼亚栎叶绿素含量指数的变化
Figure 2. Changes of chlorophyll content index of Q.virginiana under different light treatments
-
如图 3所示,在设定光照强度下,不同光照处理下弗吉尼亚栎苗木叶片净光合速率(Pn)均表现为随着光照强度(PAR)的增加逐渐增加;从光响应曲线看,50%自然光对弗吉尼亚栎净光合速率的影响不大,其光响应曲线基本接近自然光下的曲线;而25%自然光处理下弗吉尼亚栎净光合速率明显低于全光条件(图 3)。通过对光响应曲线的拟合(表 2)发现,50%自然光导致弗吉尼亚栎叶片光补偿点(LCP)和光饱和点(LSP)明显下降(P < 0.05),说明适度遮荫可以提高弗吉尼亚栎叶片对弱光的利用能力;同时适度遮荫也降低了叶片的暗呼吸速率(Rd),减少了碳水化合物的消耗。尽管大多数情况下,较低的光补偿点反映了植物对遮荫的适应性[24],然而本研究却发现,强度遮荫条件下弗吉尼亚栎光饱和点和光补偿点均明显高于全光条件(P < 0.05),这在白栎中也有类似报道[25],Dai等[26]认为全光条件下光补偿点的降低暗示植物对高光环境的一种驯化,另外也可能是植物在强度遮荫环境下生存与生长之间的一种权衡[27]。
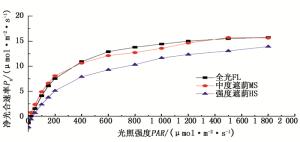
图 3 不同光处理下弗吉尼亚栎叶片光响应曲线
Figure 3. Light response curve of photosynthesis in Q. virginiana leaves under different light treatments
表 2 不同光处理对弗吉尼亚栎叶片光响应参数的影响
Table 2. Effects of light treatments on photosynthesis parameters of Q. virginiana
光响应参数Parameters 全光FL 中度遮荫MS 强度遮荫HS 表观量子效率α / (mmol·mol-1) 0.061±0.02 a 0.063±0.01 a 0.051±0.01 a 最大净光合速率Pmax /(μmol·m-2·s-1) 16.64±1.03 a 16.11±1.22 a 17.32±2.11 a 暗呼吸速率Rd/(μmol·m-2·s-1) 1.33±0.22 b 0.73±0.15 c 2.11±0.32 a 光补偿点LCP/(μmol·m-2·s-1) 21.6±5.33 b 14.4±3.21 c 36.0±10.22 a 光饱和点LSP/(μmol·m-2·s-1) 1 029.6.08 ±288.2 b 957.6±78.3 c 1 756.8±328.4 a -
进一步对弗吉尼亚栎苗木叶片叶绿素荧光参数进行测定发现,从7月底到10月初,不同时间测定的叶绿素荧光参数呈波动状态(图 4)。PSII最大光化学量子产量(Fv/Fm)作为光抑制和PSII复合体受伤的指标,反映了植物潜在的最大光合能力,当植物受到胁迫时,Fv/Fm会显著下降[22]。由图 4A可以看出,遮荫导致了Fv/Fm的小幅增加;实际量子产量(Yield)(图 4B)也呈现类似的趋势。各处理条件下的Fv/Fm和Yield随时间变化的趋势基本一致,但全光处理下的Fv/Fm在8月27日出现明显的下降,通过对试验期间日均气温变化(图 1)的分析发现,在8月底之前,刚刚经历了一次小幅的气温回升过程,推测可能是由于突然升温对植物生长产生了一定影响,导致PSII反应中心原初光能捕获效率下降,从而导致Fv/Fm下降。一些研究指出,中等强度的光照或适度遮荫有利于木本植物幼苗的生长[28-29]。本研究发现,在全光处理下,Fv/Fm值仅为0.77,显著低于其它处理,而遮荫处理的Fv/Fm值一直稳定在0.80以上,表明在光照强烈的夏季,遮荫对弗吉尼亚栎苗木生长有利,而适度遮荫有利于维持弗吉尼亚栎叶片光合器官的正常功能。
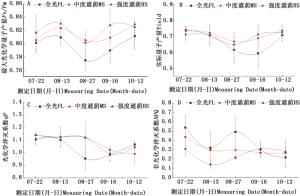
图 4 不同光照处理下弗吉尼亚栎叶绿素荧光参数变化
Figure 4. Changes of chlorophyll fluorenscence of Q.virginiana under different light treatments
光化学淬灭系数(qP)反映植物光合活性的高低,根据图 4C,遮荫条件下的qP在试验初期有小幅的增加,说明遮荫在一定程度上提高了PSII的光合活性;而随着遮荫时间的增加,到10月份时,遮荫条件下qP表现为下降趋势。非光化学猝灭系数(NPQ)作为一种光保护机制,可以将过剩的光能以热耗散的形式释放,图 4D表明,强度遮荫也导致了热耗散(NPQ)的下降(图 4D),这种趋势在8月底表现最为明显。一般来说,在强光下都有过剩光能的产生[30],而植物可以通过热耗散减轻过剩光能的伤害,表现为NPQ的增加,弗吉尼亚栎在8月底自然光下叶片NPQ明显增加,进一步说明此时光照强度已经超过光饱和点,需要通过热耗散减轻对光合机构的伤害,而自然光下生长的叶片通过叶黄素循环耗散过剩光能的能力显然要强于遮荫下生长的叶片。
-
光照是影响植物形态和功能的重要环境因子,对植物的生长、发育和形态建成具有极其重要的作用[4, 31]。研究发现,植物在遭遇遮荫胁迫之后,往往会出现基径变细、株高增加的现象,即表现为“细长”的特征,这可能是植株为了最大程度地获取光照,减少了用于植株基径生长的碳,将同化的碳更多地分配于植株的垂直生长[7, 31]。但栎属植物中的麻栎、蒙古栎(Q. mongolica Fisch. ex Ledeb.)在遮荫条件下株高增长缓慢,在弱光条件下,主要是采用保守策略进行缓慢的资源获取和消耗[7, 32]。本研究发现,弗吉尼亚栎苗木经过120 d的中度遮荫后,苗高相对生长量明显增加,而地径生长变慢;而强度遮荫后,地径和苗高生长均变慢。推测弗吉尼亚栎在不同的光环境下,生长策略也有所不同,中度遮荫条件下,通过株高快速生长的冒险策略进行快速的资源获取和消耗;而在强度遮荫条件下,则倾向于采用保守策略,与麻栎、蒙古栎等类似。
植物光合作用的光响应曲线对于了解植物光化学过程中的光化学效率具有重要意义。表观量子效率(α)反映了叶片在弱光情况下的光合能力,其值越大,表明植物吸收与转换光能的色素蛋白复合体可能越多,利用弱光能力越强[33]。本研究发现,弗吉尼亚栎在中度遮荫条件下,具有较高的表观量子效率,说明弗吉尼亚栎对弱光具有较强的适应性,但与全光下的表观量子效率差异并不明显。光补偿点与光饱和点的高低反映了植物对弱光的利用能力,光补偿点和光饱和点的降低意味着植物在光照有限的条件下能够以最大能力利用低光量子通量,进行最大可能的光合作用,从而利于在弱光下进行有机物的积累,是植物对遮荫环境的适应性表现[4, 34]。弗吉尼亚栎在50%自然光条件下的光饱和点和光补偿点均明显低于对照,说明经过一段时间的遮荫处理,弗吉尼亚栎已经能够适应较低的光照强度,出现阴生植物的光响应特性。遮荫条件下,光补偿点和暗呼吸速率的降低是植物光驯化的一种方式[35],这在很多植物中均有报道[30, 36]。然而也有研究发现,强度遮荫条件下,光补偿点会升高,Dai等[26]在研究遮荫对三叶青(Tetrastigma hemsleyanum Diels et Gilg)光合作用时发现,50%75%的遮荫条件下,光补偿点明显高于全光条件,被认为是三叶青光合作用在高光环境下的一种驯化。本研究也发现,弗吉尼亚栎在强度遮荫条件下光饱和点和光补偿点均高于全光条件,一方面可能与高光驯化有关,另一方面,推测可能与光照强度以及遮荫强度甚至树种自身特性有关,大多数研究采用的遮荫方法均将自然光照的百分比作为遮荫强度,然而不同地区的自然光照本身存在很大差异,由此获得的遮荫强度也各不相同。石玉波等[25]在上海地区研究了白栎对不同强度遮荫的响应,也发现在强度遮荫(60%和80%)条件下,白栎光补偿点和光饱和点也有高于自然光的情况出现。上海与杭州均属长三角地区,气候条件相似,且弗吉尼亚栎与白栎同属于白栎组树种,两者在生物学特性方面存在很多相似,由此推测,不同区域以及不同植物之间,光合作用对遮荫的响应可能存在差异。中度遮荫条件下暗呼吸速率有所下降,反映了弗吉尼亚栎在这种光环境下的适应性,通过降低呼吸作用减少对碳水化合物的消耗;然而强度遮荫以后,弗吉尼亚栎暗呼吸速率明显增加,有悖于光驯化的理论,这可能与夏季高温期间水分因子的干扰有关,有研究表明,植物暗呼吸随水分亏缺程度加大而降低[37],夏季高温情况下,全光照条件下的水分消耗明显高于遮荫条件,这可能会使植物处于暂时缺水状态,因此全光条件下的呼吸速率会有所下降。另一方面,强度遮荫条件下暗呼吸的增强,可能也是导致弗吉尼亚栎苗高和地径生长变慢的重要原因。
叶绿素荧光与光合作用各反应过程密切相关,环境因子对光合作用的影响可通过荧光参数反映出来。PSII最大光化学量子产量(Fv/Fm)反映了光系统II反应中心的光能转换效率,当植物处于非逆境条件下时,Fv/Fm一般为0.750.85,且不受物种的影响,但在逆境或受伤害时会明显降低[21-22]。本研究发现,尽管在不同时间点测定的Fv/Fm有所不同,但基本保持在正常范围,说明弗吉尼亚栎在遮荫条件下并没有受到明显的光胁迫,与表观量子效率的推断一致,同时,PSII实际量子产量(Yield)也表现出与Fv/Fm相似的变化趋势。由此进一步说明,弗吉尼亚栎在遮荫条件下光合器官并未受到严重的损伤,反映了其对遮荫环境的适应。在全光条件下,特别是南方夏季光照强烈时,自然光照强度超过光饱和点,光能利用率下降,因此,全光条件下最大光化学量子产量、实际量子产量以及光化学猝灭系数(qP)会出现下降趋势,这在很多木本植物中均有报道[6-7]。而过多的光能则以热能的形式耗散,因此光照较强时,非光化学猝灭(NPQ)也较高。遮荫条件下,PSⅡ把吸收的光能较多地分配给光合电子传递用于叶片的光合作用,表现出较高的光合作用潜力。陈秋夏等[6]对不同遮荫处理的青冈栎容器苗叶片叶绿素荧光研究发现,随着透光率的降低,青冈栎叶片qP逐渐增加,而NPQ逐渐降低,全光处理下的NPQ明显高于遮荫处理。作者发现,试验开始的1个月内,遮荫使弗吉尼亚栎qP表现出增加趋势,而NPQ呈下降趋势;但8月27日以后,全光处理的叶片qP开始增加,NPQ开始下降。推测这可能与试验期间的温度、光照的季节性变化有关。近年来,有关叶绿素荧光参数季节变化的研究很多,茶树(Camellia sinensis (L.) O. Ktze.)能通过自身调节机制改变NPQ以适应夏秋季节的变化[35],Yang等[36]发现叶绿素荧光参数的季节性变化反映了温带森林植物的光合作用变化。本研究中,遮荫条件下弗吉尼亚栎叶片光合作用以及叶绿素荧光参数的变化,除了与试验处理相关以外,自然环境的季节性变化也是一个重要的影响因素,而目前大多数针对遮荫处理的研究并未考虑光合作用以及荧光参数的动态变化,特别是在一个比较长的生长季节中,忽略了植物自身的调节机制,由此导致相关研究结论的相异。因此,在比较植物光合及荧光参数对遮荫条件的响应研究时,需考虑多方面的影响因子,包括试验区的气候条件、测试时间等;而控制试验本身也需要进一步改良,尽可能使光照条件成为限制因子。
-
本研究表明,弗吉尼亚栎幼苗对遮荫具有一定的耐受性,可以通过生长方式的改变以适应不同的遮荫条件。长三角地区夏季光照强烈,适度遮荫有利于苗木维持正常的光合作用,因此建议在育苗过程中采取适当的遮荫措施;而强度遮荫不利于弗吉尼亚栎干物质的积累,弗吉尼亚栎冠幅较大,人工林中容易形成郁闭,直接影响了其林下幼苗更新,因此弗吉尼亚栎林分密度不宜过大,林下维持50%以上的自然光照有利于幼苗的自然更新。另外,尽管本研究为弗吉尼亚栎对光环境的需求提供了一个大致的参考范围,但要获得全面的光响应研究,依然需要扩大遮荫条件的设置,如设置不同光照强度的轻度遮荫、中度遮荫条件,以期获得弗吉尼亚栎生长最佳的光环境条件。
遮荫对弗吉尼亚栎苗木生长及光合-荧光参数的影响
Effects of Shading on Growth, Photosynthesis and Chlorophyll Fluorescence Parameters in Leaves of Quercus virginiana Seedlings
-
摘要:
目的 揭示美国引进树种弗吉尼亚栎幼苗对光照强度的响应特征,为弗吉尼亚栎的引种驯化和高效栽培提供科学依据。 方法 采用人工遮荫法,研究3种光环境下(全光——100%自然光、中度遮荫——50%自然光和强度遮荫——25%自然光)弗吉尼亚栎苗木生长、光合作用以及叶绿素荧光参数的变化。 结果 表明:中度遮荫明显促进弗吉尼亚栎苗高生长(P < 0.05),但对地径生长影响不明显;而强度遮荫条件下,地径生长受到明显抑制(P < 0.05),苗高虽有所下降,但与对照相比差异不明显。对弗吉尼亚栎光响应曲线的观测表明:与全光条件相比,中度遮荫条件下,弗吉尼亚栎叶片暗呼吸速率(Rd)、光饱和点(LSP)以及光补偿点(LCP)均明显下降(P < 0.05),说明适度的遮荫有助于提高弗吉尼亚栎对光的利用效率,促进干物质的积累;而强度遮荫条件下,光补偿点和光饱和点明显上升,暗呼吸速率明显增加(P < 0.05)。遮荫并未对表观量子效率(α)和最大净光合速率(Pmax)产生明显的影响。进一步对叶绿素荧光参数分析发现,遮荫条件下,PSⅡ最大光化学量子产量(Fv/Fm)和实际光量子产量(Yield)有所增加,但均在正常范围,说明遮荫对PSⅡ活性并未产生胁迫。 结论 弗吉尼亚栎幼苗对遮荫均具有一定的耐受性,中度遮荫有利于苗高生长;而强度遮荫不利于干物质的积累。因此,建议在营林以及人工栽培过程中维持50%以上的自然光照。 Abstract:Objective To discover the response to light intensity of Quercus virginiana, and the effects of different levels of shading on growth, photosynthesis and chlorophyll fluorescence parameters. Method Three light levels (full light-100% natural light, moderate shading-50% natural light and high shading-25% natural light) were set in greenhouse by covering different layers of shading net. The growth, chlorophyll content index, photosynthesis and chlorophyll fluorescence parameters were determined during the experiment. Result The results showed that the height growth of seedlings increased significantly (P < 0.05) under moderate shading condition, while no obvious change was observed in the growth of basal diameter. Under high shading conditions, the height growth changed a little compared with control, whereas the growth of basal diameter was inhibited significantly (P < 0.05). Under moderate shading, the dark respiration rate (Rd), light compensation point (LCP) and light saturation point (LSP) declined and whereas increased significantly under high shading, which showed that moderate shading might be helpful to improve the light use efficiency and increase the dry mass accumulation in leaves of Q. virginiana. There were no obvious changes in apparent quantum yield (α) and the maximum net photosynthetic rate (Pmax). Further analysis of chlorophyll fluorescence parameters showed that PSII primary energy conversion efficiency (Fv/Fm) and photochemical efficiency of PSII in the light (Yield) remained a reasonable range under two light levels, which indicated that the PSII activity might be not damaged. Conclusion These results indicate that Q. virginiana could tolerate certain shading conditions, however, high shading was considered to be detrimental to the accumulation of dry matter, and light condition with more than 50% natural light would be recommended during the silviculture and nursery cultivation. -
Key words:
- Quercus virginiana
- / shading
- / photosynthesis
- / chlorophyll fluorescence
- / chlorophyll content index
-
图 2 不同光照处理下弗吉尼亚栎叶绿素含量指数的变化
Figure 2. Changes of chlorophyll content index of Q.virginiana under different light treatments
图 3 不同光处理下弗吉尼亚栎叶片光响应曲线
Figure 3. Light response curve of photosynthesis in Q. virginiana leaves under different light treatments
图 4 不同光照处理下弗吉尼亚栎叶绿素荧光参数变化
Figure 4. Changes of chlorophyll fluorenscence of Q.virginiana under different light treatments
表 1 夏季遮阴对弗吉尼亚栎苗木生长的影响
Table 1. Effects of shading on growth of Q. virginiana
生长指标
Growth parameters处理
Treatments处理前
Before treatments处理后
After treatments相对生长率
Relative growth rate/%苗高Height/cm 全光FL 59.25±7.53 a 62.00±7.97 a 4.73±1.55 b 中度遮荫MS 57.75±10.08 a 61.50±7.61 a 7.64±0.29 a 强度遮荫HS 60.58±12.32 a 61.79±12.07 a 2.22±0.58 b 地径Basal diameter/cm 全光FL 3.99±0.37 a 4.96±0.47 a 24.40±3.06a 中度遮荫MS 3.73±0.59 a 4.46±0.62 a 19.90±2.08 a 强度遮荫HS 3.87±0.62 a 4.24±0.63 b 9.79±2.06 b 注:表中数据为平均值±标准差,小写字母不同代表不同光照强度之间差异显著(P < 0.05)。下同。
Notes: Values are means±sd, the different letters represent the significant difference among different light treatment(P < 0.05). The same as below. 下载: 导出CSV
下载: 导出CSV
表 2 不同光处理对弗吉尼亚栎叶片光响应参数的影响
Table 2. Effects of light treatments on photosynthesis parameters of Q. virginiana
光响应参数Parameters 全光FL 中度遮荫MS 强度遮荫HS 表观量子效率α / (mmol·mol-1) 0.061±0.02 a 0.063±0.01 a 0.051±0.01 a 最大净光合速率Pmax /(μmol·m-2·s-1) 16.64±1.03 a 16.11±1.22 a 17.32±2.11 a 暗呼吸速率Rd/(μmol·m-2·s-1) 1.33±0.22 b 0.73±0.15 c 2.11±0.32 a 光补偿点LCP/(μmol·m-2·s-1) 21.6±5.33 b 14.4±3.21 c 36.0±10.22 a 光饱和点LSP/(μmol·m-2·s-1) 1 029.6.08 ±288.2 b 957.6±78.3 c 1 756.8±328.4 a
下载: 导出CSV
-
[1] Scholes J D, Press M C, Zipperlen S W. Differences in light energy utilization and dissipation between dipteroearp rain forest tree seedlings[J]. Oecologia, 1997, 109(1):41-48. doi: 10.1007/s004420050056 [2] 刘建锋,杨文娟,江泽平. 等.遮荫对濒危植物崖柏光合作用和叶绿素荧光参数的影响[J]. 生态学报, 2011, 31(20):5999-6004. [3] Senevirathna A M W K, Stirling C M, Rodrigo V H L. Growth, photosynthetic performance and shade adaptation of rubber (Hevea brasiliensis) grown in natural shade[J]. Tree Physiology, 2003, 23(10):705-712. doi: 10.1093/treephys/23.10.705 [4] 李东胜,白庆红,李永杰,等. 光照条件对蒙古栎幼苗生长特性和光合特征的影响[J]. 生态学杂志,2017,36(10):2744-2750. [5] 贺顺钦,王发其. 辽东栎苗木早期生长与光的关系[J]. 林业科学研究,2001, 14(6):697-700. doi: 10.3321/j.issn:1001-1498.2001.06.018 [6] 陈秋夏,廖亮,郑坚,等. 光照强度对青冈栎容器苗生长和生理特征的影响[J]. 林业科学,2011, 47(12):53-58. doi: 10.11707/j.1001-7488.20111208 [7] 徐飞,郭卫华,徐伟红,等. 不同光环境对麻栎和刺槐幼苗生长和光合特征的影响[J]. 生态学报, 2010, 30(12):3098-3107. [8] Haller J M. Quercus virginiana:the southern live oak[J]. Arbor Age,1992, 12(5):30. [9] Harms W R. Quercus virginiana Mill. live oak[M]//Burns R M, Honkala B H. Silvics of North America:Vol.2 hardwoods. Agric. handb. 654. Washington, DC:U.S. Department of Agriculture, Forest Service, 1990:751-754. [10] 陈益泰,陈雨春,黄一青,等. 抗风耐盐常绿树种弗吉尼亚栎引种初步研究[J]. 林业科学研究, 2007,20(4):542-546. doi: 10.3321/j.issn:1001-1498.2007.04.020 [11] 陈益泰,孙海菁,王树凤,等. 5种北美栎树在我国长三角地区的引种生长表现[J]. 林业科学研究, 2013,26(3):344-351. doi: 10.3969/j.issn.1001-1498.2013.03.013 [12] Cavender-Bares J M, Kitajima K, Bazzaz F A. Multiple trait associations in relation to habitat differentiation among 17 oak species in North Central Florida[J]. Ecological Monographs, 2004, 74(4):635-662. doi: 10.1890/03-4007 [13] Spector T, Putz F E. Crown retreat of open-grown Southern live oaks (Quercus virginiana) due to canopy encroachment in Florida, USA[J]. Forest Ecology and Management, 2006, 228(1-3):168-176. doi: 10.1016/j.foreco.2006.03.001 [14] 王树凤,胡韵雪,李志兰,等. 盐胁迫对弗吉尼亚栎生长及矿质离子吸收、运输和分配的影响[J]. 生态学报, 2010, 30(17):4609-4616. [15] 王树凤,胡韵雪,孙海菁,等. 盐胁迫对2种栎树苗期生长和根系生长发育的影响[J].生态学报, 2014, 34(4):1021-1029. [16] 杨舒婷,曲博,李谦盛,等. 弗吉尼亚栎幼苗对高温胁迫的生理响应[J]. 江西农业大学学报,2015,37(1):90-95. doi: 10.3969/j.issn.1000-2286.2015.01.015 [17] 宋以刚,邓敏,王妍婷. 弗吉尼亚栎种子萌发特性[J]. 生态学杂志, 2015, 4(5):1295-1300. [18] Ghasemi M, Arzani K, Yadollahi A, et al. Estimate of leaf chlorophyll and nitrogen content in asian pear (Pyrus serotina Rehd.) by CCM-200[J]. Notulae Scientia Biologicae, 2011, 3(1):91-94. doi: 10.15835/nsb315623 [19] 叶子飘,于强. 一个光合作用光响应新模型与传统模型的比较[J]. 沈阳农业大学学报, 2007, 38(6):771-775. doi: 10.3969/j.issn.1000-1700.2007.06.001 [20] 叶子飘,于强. 光合作用光响应模型的比较[J]. 植物生态学报,2008, 32(6):1356-1361. doi: 10.3773/j.issn.1005-264x.2008.06.016 [21] Maxwell K, Johnson G N. Chlorophyll fluorescence-A practical guide[J]. Journal of Experimental Botany, 2000, 51(345):659-668. doi: 10.1093/jexbot/51.345.659 [22] 张守仁. 叶绿素荧光动力学参数的意义及讨论[J].植物学通报, 1999, 16(4):444-448. doi: 10.3969/j.issn.1674-3466.1999.04.021 [23] 吴飞燕,伊力塔,李修鹏,等. 不同光照强度对石栎幼苗叶绿素含量及叶绿素荧光参数的影响[J].东北农业大学学报,2012, 43(4):88-92. doi: 10.3969/j.issn.1005-9369.2012.04.016 [24] 潘瑞炽. 植物生理学[M].北京:高等教育出版社,2008:61-62. [25] 石玉波,卓丽环,辛雅芬,等.不同遮光处理对白栎光合生理特性的影响[J]. 安徽农业科学, 2010, 38(2):956-958. doi: 10.3969/j.issn.0517-6611.2010.02.168 [26] Dai Y J, Shen Z G, Liu Y, et al. Effects of shade treatments on the photosynthetic capacity, chlorophyll fluorescence, and chlorophyll content of Tetrastigma hemsleyanum Diels et Gilg[J]. Environmental and Experimental Botany, 2009,65(2-3):177-182. doi: 10.1016/j.envexpbot.2008.12.008 [27] Falster D S, Westoby M. Tradeoffs between height growth rate,stem persistence and maximum height among plant species in a post-fire succession[J]. Oikos, 2005, 111(1):57-66. doi: 10.1111/j.0030-1299.2005.13383.x [28] Gardiner E S, Hodges J D. Growth and biomass distribution of cherrybark oak (Quercus pagoda Raf) seedlings as influenced by light availability[J]. Forest Ecology and Management, 1998, 108(1-2):127-134. doi: 10.1016/S0378-1127(98)00220-5 [29] 闫兴富,曹敏.林窗对热带雨林冠层树种绒毛番龙眼幼苗生长的影响[J].应用生态学报,2008,19(2):238-244. [30] 杨兴洪,邹琦,赵世杰. 遮荫和全光下生长的棉花光合作用和叶绿素荧光特征[J]. 植物生态学报,2005, 29(1):8-15 doi: 10.3321/j.issn:1005-264X.2005.01.002 [31] 薛思雷,王庆成,孙欣欣,等. 遮荫对水曲柳和蒙古栎光合、生长和生物量分配的影响[J]. 植物研究, 2012, 32(3):354-359. [32] 郭志华,张旭东,黄玲玲,等. 落叶阔叶树种蒙古栎(Quercus mongolica)对林缘不同光环境光能和水分的利用[J]. 生态学报, 2006, 26(4):1047-1056. doi: 10.3321/j.issn:1000-0933.2006.04.010 [33] Richardson A D, Berlyn G P. Spectral reflectance and photosynthetic properties of Betula papyrifera (Betulaceae) leaves along an elevational gradient on Mt. Mansfield,Vermont, USA[J]. American Journal of Botany, 2005, 89(1):88-94. [34] 梁文斌,聂东伶,吴思政,等. 遮荫对短梗大参苗木光合作用及生长的影响[J]. 生态学杂志, 2015, 34(2):413-419. [35] 唐敏, 翟秀明, 姚永红, 等. 不同茶树品种(系)叶绿素荧光参数季节性差异研究[J]. 西南农业学报, 2015, 28(1):79-83 [36] Yang H L, Yang X, Zhang Y G, et al. Chlorophyll fluorescence tracks seasonal variations of photosynthesis from leaf to canopy in a temperate forest[J]. Global Change Biology, 2017, 23(7):2874-2886. doi: 10.1111/gcb.13590 [37] Crous K Y, Zaragoza-Castells J, Ellsworth D S, et al. Light inhibition of leaf respiration in field-grown Eucalyptus saligna in whole-tree chambers under elevated atmospheric CO2 and summer drought[J]. Plant, Cell and Environment, 2012, 35(5):966-981. doi: 10.1111/j.1365-3040.2011.02465.x -

 点击查看大图
点击查看大图
计量
- 文章访问数: 3993
- HTML全文浏览量: 2018
- PDF下载量: 46
- 被引次数: 0
