


-
新疆野果林树种类型丰富,由野苹果群系、野杏群系、野樱桃李群系、野欧洲李群系、野核桃群系、野山楂群系和稠李群系7个群系组成[1]。天山野果是地质年代第三纪的孑遗物种,已有千万年的历史,是新疆天山生物多样性的重要组成部分,是世界多种栽培果树的起源种[2]。伊犁野果林也被列入中国优先保护生态系统名录。然而近年来,天山野果林的主体部分——新源县、巩留县果树大批量死亡,果林出现衰退现象。当前迫切需要探究其可能的原因及存在的风险,以防野果林衰退面积进一步加大。
疫霉菌(Phytophthora)属卵菌纲(Oomycetes)霜霉目(Peronsporales)腐霉科(Pythiaceae)疫霉属(Phytophthora),是一类世界性分布的重要植物病原菌,该属的大多数种都对寄主植物具有严重的侵害性和破坏性,曾引起马铃薯晚疫病、大豆疫霉根腐病、栎树猝死病等多种严重的植物病害,是世界范围内多次报道的森林衰退的罪魁祸首[3]。
本研究对在2016、2017年于新疆伊犁新源县、巩留县设点,通过林间溪流和土壤悬浮液诱捕方法获得的疫霉菌株(程元,待发表)接种到天山野果林的四种主要群系——苹果、杏、核桃、山楂的叶片上,进行叶片致病性实验,以期探究疫霉对新疆天山野果林衰退的影响及潜在风险。
-
本试验使用于2016、2017年在新疆伊犁巩留县、新源县野果林内通过将2片正面,2片背面共4片野苹果叶片装入网袋放入溪流中,每57天换取叶片,分离叶片病健交界处,置于CARP+选择性培养基上,和通过挖取衰退苹果树下的土壤200 g,装袋,加入纯水静置1天后,放入4片野苹果叶片于土壤悬浮液中,5~7天后取出叶片,分离于选择性培养基CARP+中[4-5],收集了88株疫霉菌株共5种疫霉种,分别为P. gregata、P. sp.、P. plurivora、P. gonapodyides、P. lacustris。13株菌株,编号为GL1710.6,GL1710.9,GLW1717.8,XYW1722.2, S040203,XYS85.1, XYS77.1, XYW1710.1, XYW1723.3, XYS42,XYW1701.3, XYW1711.1, XYW1724.4作为试验菌株(表 1)。
表 1 供试菌株信息
Table 1. The information of tested strains
种名
Species菌株号
Strain no.采集地
Location采集方法
MethodP. gregate GL1710.6 巩留 溪流诱捕 GL1710.9 巩留 溪流诱捕 P. sp. GLW1717.8 巩留 溪流诱捕 XYW1722.2 新源 溪流诱捕 P. plurivora S040203 巩留 土壤诱捕 XYS85.1 新源 土壤诱捕 XYS77.1 新源 土壤诱捕 P. gonapodyides XYW1710.1 新源 溪流诱捕 XYW1723.3 新源 溪流诱捕 XYS42 新源 土壤诱捕 P. lacustris XYW1701.3 新源 溪流诱捕 XYW1711.1 新源 溪流诱捕 XYW1724.4 新源 溪流诱捕 -
试验选取新疆天山野果林中主要的群系树种——苹果、杏、核桃、山楂的健康叶片作为试验材料,摘取叶片时,选择中龄、完整且大小相近的叶片,连同叶柄一起摘取。由于核桃叶片过大,后在实验室进行了剪切。
-
配置5%的水琼脂(5 g琼脂+95 g蒸馏水)灭菌后备用。
-
将实验室存有的菌株活化后,转入V8培养基培养,当菌丝长满培养基时,利用直径1 cm的玻璃打孔器进行打孔备用。
-
在野外采取新鲜健康的苹果、杏、核桃、山楂叶片各50片,带回实验室用蒸馏水冲洗干净,再用沾有75%酒精的脱脂棉擦拭叶片。将叶片放入垫有滤纸的已灭菌的玻璃培养皿中。
-
将装有叶片的玻璃培养皿拿至超净工作台内,利用接种针在叶片中部穿刺伤害,每个叶片穿刺3个伤口。将打孔后的圆形菌丝块放置伤口上,并用水琼脂进行覆盖封口。每个菌株设3组重复,共(13菌株+1对照)×3个重复×4种寄主叶片=168个样本,每个树种设一组对照,对照进行穿刺后,接种无菌种的相同V8培养基,并用水琼脂覆盖。将叶片放入25℃的恒温光照培养箱中培养一周,每日12 h光照,光照度为30 000 lux,12 h黑暗。
-
取出产生病斑的叶片,将病健交界处切成1 cm×1 cm的小块,放入CARP+选择性培养基中进行再分离,观察检验致病菌是否为疫霉菌株。
-
在穿刺部位,利用十字交叉法测量1周后叶片病斑的长度和宽度,根据椭圆面积计算公式(S=πab,π为圆周率,a、b分别为半长轴和半短轴长)计算病斑面积大小,叶片边缘产生的变色不计为病斑大小。取3组重复的平均值作为该菌株号的病斑大小,将同种菌不同菌株号的平均值作为该菌种的病斑大小[7]。并通过R x64 3.5.1.软件对数据进行双因素方差分析,判定树种及菌种间对叶片病斑是否有显著性差异,并绘制直方图。
-
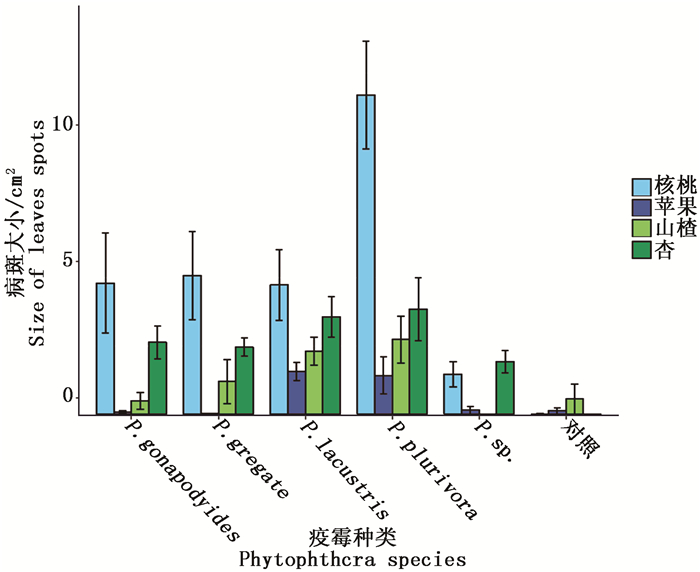
不同疫霉菌株接种不同寄主叶片后,产生的病斑大小如图 1,通过测量计算,病斑面积见表 2。其中病斑最大的为菌株编号XYS85.1(P. plurivora)侵染的核桃叶片,平均病斑面积为13.12 cm2,侵染面积为0的有GL1710.6、GLW1717.8(P. gregate)侵染的苹果叶片和GL1710.9(P. gregate)、XYW1722.2(P. sp.)侵染的山楂叶片。利用R软件对测量计算出的病斑面积进行有重复双因素方差分析,所得的方差分析结果如表 3所示。由此可知,不同疫霉病原菌和不同寄主树种产生的病斑大小都有极显著差异,说明菌种和树种的不同对叶片的病斑大小皆有显著性影响。

图 1 病斑大小
Figure 1. Size of leaves spots
表 2 菌株接种叶片病斑大小
Table 2. The size of leaf spots inoculated by the strain
cm2 种名Species 菌株号Strain no. 苹果Apple 杏Apricot 山楂Hawthorn 核桃Walnut P. gregate GL1710.6 0±0 2.56±0.74 2.39±1.4 1.58±0.35 GL1710.9 0.03±0.03 2.35±0.06 0±0 8.57±0.77 P. sp. GLW1717.8 0±0 1.91±0.68 0.01±0.01 2.07±0.54 XYW1722.2 0.3±0.28 1.94±0.66 0±0 0.87±0.64 P. plurivora S040203 0.67±0.44 3.56±1.71 4.46±2.14 11.88±3.99 XYS85.1 0.4±0.07 4.9±2.36 2.42±0.72 13.12±2.34 XYS77.1 3.19±1.69 3.06±2.62 1.32±1.02 10.08±4.76 P. gonapodyides XYW1710.1 0.02±0.02 0.76±0.34 0.01±0.01 4.25±2.39 XYW1723.3 0.17±0.17 3.72±1.28 0.95±0.94 2.73±1.25 XYS42 0.04±0.01 3.41±0.29 0.5±0.15 7.41±5.21 P. lacustris XYW1701.3 1.28±0.86 3.26±1.8 2.51±1.51 4.02±0.63 XYW1711.1 1.56±0.19 5.31±0.68 2.21±0.85 4.45±0.76 XYW1724.4 1.83±0.72 2.12±0.7 2.21±0.27 5.73±4.29 对照 0.12±0.12 0±0 0.55±0.54 0.01±0.01 表 3 双因素方差分析表
Table 3. ANOVAs of effects of trees and Phytophthora on spots size
因素
Effect自由度
DFF值
FTrees 3 27.631*** Phytophthora 4 11.052** Residuals 12 Sigif. codes: ***0.001. **0.01 -
计算接种5种疫霉后植物叶片产生的病斑大小,标准误差,利用R软件作图如图 2。
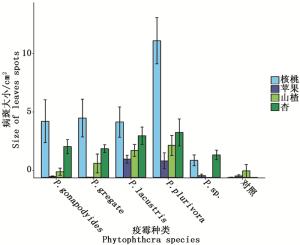
图 2 不同疫霉菌接种叶片后病斑大小
Figure 2. Size of the leaves spots after different Phytophthora inoculated
由图 2可知,5种疫霉菌接种4种寄主植物叶片后皆产生病斑,说明其对该4种植物叶片都有致病性。其中接种P. plurivora后,植物叶片产生的病斑最大,说明在这几种疫霉菌中,P. plurivora对该4种植物叶片的致病性最强。其次为P. lacustris疫霉菌,P. gregate和P. gonapodyides产生的叶片病斑大小接近,说明两者对该4种植物叶片的致病性相近。病斑最小的为P. sp.接种的叶片,说明该疫霉菌在这几种疫霉菌中对4种植物叶片的致病性最小。
对比5种疫霉菌接种叶片后产生的病斑大小,总结其病斑大小依次为:P. plurivora>P. lacustris>P. gregate≈P. gonapodyides>P. sp.,对4种叶片的致病性大小依次为:P. plurivora>P. lacustris>P. gregate≈P. gonapodyides>P. sp.。
-
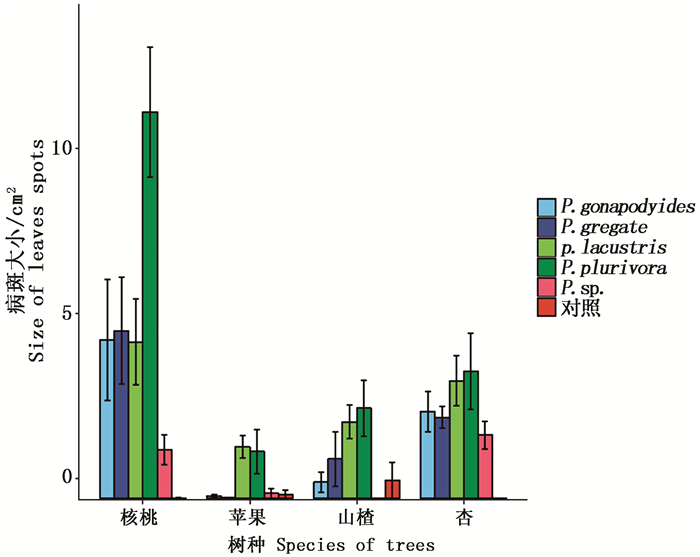
计算4种植物叶片被接种后产生的病斑大小, 标准误差, 利用R软件作图如图 3:
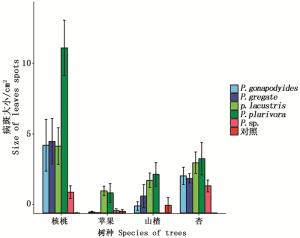
图 3 不同寄主叶片接种疫霉菌后病斑大小
Figure 3. Size of the leaves spots after different leaves inoculated
由图 3可知,4种寄主植物叶片皆产生病斑,说明其叶片对5种疫霉菌皆感病。其中,核桃叶片产生的病斑最大,说明在这4种树种中,核桃叶片最易感病,其次为杏树叶片,在杏树叶片中接种5种疫霉菌后产生的病斑大小差异不大,说明杏树叶片对这5种疫霉菌的感病性差异不大。接种后苹果叶片产生的病斑最小,说明苹果叶片在这4种寄主树种叶片中,不易感病。对比4种植物叶片接种后产生的病斑大小,总结其病斑大小依次为:核桃>杏>山楂>苹果,叶片感病性依次为:核桃>杏>山楂>苹果。
-
当前致病性研究多为1种病原物作用于1种寄主植物,本试验通过5种病原和4种寄主的多维角度对新疆野苹果林内疫霉对叶片的致病性进行差异性探究。P. plurivora于2009年被详细描述,寄主广泛,并造成了许多原生树种甚至生态系统的毁灭性衰退,被认为是目前最具威胁的疫霉属物种,具有强致病性[8-12]。在本试验中,P. plurivora对寄主叶片的致病性最强,与当前研究结果一致。P. lacustris于2012年被详细描述,在栎树和李树的灌根试验中,其引起显著的根系损伤和细根坏死。在对杏树、桃树进行茎段菌丝块接种试验时,具有中等致病性,引起的病斑长分别为3 cm,6 cm[13]。P. gregata属clade6,于2011年被详细描述,该族群大多都有致病性,而P. gregata能抑制植物的生长却很难造成植物死亡,其有致病性但不强[14-15]。P. gonapodyides可寄生蔷薇科、松科等11科寄主植物,导致栎树的根系腐烂和茎部病变,且其致病性总与潮湿的部位有关,但引起的病害程度较小,属于弱致病菌[16]。P. sp.疑似新种,致病性有待探究,就本试验来说,对5种寄主叶片的致病性较弱。
相同的菌株接种不同的树种叶片后,产生的叶片病斑大小不同,可能与叶片的厚薄、有无绒毛有关。就本试验而言,较薄且无柔毛的核桃叶片更易感病,而有柔毛的苹果叶片则较难感病。同时,苹果、杏、山楂为蔷薇科(Rosaceae)植物,核桃为胡桃科(Juglandaceae)植物。据本实验推测新疆野果林内的疫霉菌较蔷薇科植物叶片更易侵染胡桃科植物叶片。
试验中发现部分山楂叶片的基部有大面积病斑,由图 4所示,可能是疫霉菌在伤口处侵入后,沿主叶脉对叶片基部进行了侵染。但本试验中,对照组也出现了微小病斑,说明在试验过程中可能有其他病原菌通过叶片伤口进行了侵染。

图 4 山楂接种疫霉后产生的病斑
Figure 4. The leaves spots of hawthorn
-
通过利用菌丝块接种离体叶片的方法,对新疆野果林疫霉菌对植物叶中致病性的研究可以得出以下结论:①P. plurivora、P. lacustris、P. gregate、P. gonapodyides、P. sp.5种疫霉菌对苹果、杏、山楂、核桃的植物叶片致病性有显著性差异,苹果、杏、山楂、核桃4种植物叶片对以上疫霉菌的感病性亦有显著性差异, 即疫霉的病原菌种类和寄主叶片的植物种类都会影响病斑的大小。②在新疆采集的5种疫霉菌对苹果、杏、山楂、核桃叶片的致病性大小依次为P. plurivora>P. lacustris>P. gregate≈P. gonapodyides>P. sp.。③寄主植物叶片对以上疫霉菌的感病性大小依次为核桃>杏>山楂>苹果。该研究表明在新疆野果林防治疫霉时,应多注重P. plurivora、P. lacustris的防治,同时新疆野核桃林作为重要的经济树种林,应多关注其健康状况,提前做好预防措施,尽量减少疫霉菌对野核桃林的危害。在此研究基础上,未来的试验可通过灌根、菌丝块接种茎部等植物活体接种试验,进一步深入研究新疆野果林疫霉菌对野果林内树种的致病性影响,以期对野果林进行及时有效的保护。
新疆野果林疫霉菌对植物叶片致病性的测定与分析
Leaf Pathogenicity of Phytophthora in Wild Fruit Forest of Xinjiang
-
摘要:
目的 从病原菌和寄主两个维度,探究新疆野果林中疫霉菌对野果林内主要树种叶片的致病性,了解新疆野果林衰退的可能原因及疫霉对新疆野果林造成的潜在风险。 方法 利用菌丝块接种离体叶片的方法,将诱捕自新疆野果林的5种疫霉(Phytophthora plurivora、P.gonapodyides、P.lacustris、P.gregate、P.sp.)接种到苹果、杏、山楂和核桃的健康离体叶片上,一周后观察测量、计算病斑大小,并用R软件对数据进行双因素方差分析和作图。 结果 实验结果及数据分析表明:供试5种疫霉菌株接种到4种植物叶片上后,病斑大小存在显著性差异。其中,P.plurivora和P.lacustris产生的病斑较大,最大分别为18.68 cm2和14.14 cm2;寄主植物中,核桃和杏树叶片产生病斑较大,最大分别为18.68 cm2和9.55 cm2。 结论 供试树种和菌种对叶片病斑大小都有显著影响;供试5种疫霉对苹果、杏、山楂、核桃叶片的致病性依次为:P.plurivora > P.lacustris > P.gregate≈P.gonapodyides > P. sp.;苹果、杏、山楂和核桃这4种植物叶片对以上5种疫霉菌的感病性依次为:核桃>杏>山楂>苹果。 Abstract:Objective Pathogenicity investigation of the Phytophthora was carried out among the main fruit tree species in Xinjiang wild forest to understand the possible causes of the decline of wild fruit forest in Xinjiang and the potential risks caused by Phytophthora. Method By inoculating mycelium plugs artificially on the in vitro leaves, collections of Phytophthora:Phytophthora plurivora, P. gonapodyides, P. lacustris, P. gregate, P. sp. from Xinjiang were inoculated in the health leaves of apple, apricot, hawthorn, and walnut trees, respectively, and observing and recording the disease spot size one week later. Two-way analysis of variance (ANOVA) was implemented to find the differences among the treatments by software R. Result There were significant differences in the size of disease spots after inoculation of 5 Phytophthora strains on 4 host leaves. Among them, P. plurivora and P. lacustris had larger lesions after inoculation on leaves which were 18.68 cm2 and 14.14 cm2. Among the host plants, the leaves of walnut and apricot trees had larger disease spots which were 18.68 cm2 and 9.55 cm2. Conclusion The Phytophthora pathogenicity on the leaves of apple, apricot, hawthorn and walnut is in the order of P. plurivora > P. lacustris > P. gregate≈P. gonapodyides > P. sp. Among leaves of the 4 host trees (apple, apricot, hawthorn and walnut), the susceptibility to the 5 Phytophthora strains mentioned above is in the order of walnut > apricot > hawthorn > apple. -
Key words:
- pathogenicity
- / Phytophthora
- / leaf spots
- / wild fruit forest
-
图 2 不同疫霉菌接种叶片后病斑大小
Figure 2. Size of the leaves spots after different Phytophthora inoculated
图 3 不同寄主叶片接种疫霉菌后病斑大小
Figure 3. Size of the leaves spots after different leaves inoculated
表 1 供试菌株信息
Table 1. The information of tested strains
种名
Species菌株号
Strain no.采集地
Location采集方法
MethodP. gregate GL1710.6 巩留 溪流诱捕 GL1710.9 巩留 溪流诱捕 P. sp. GLW1717.8 巩留 溪流诱捕 XYW1722.2 新源 溪流诱捕 P. plurivora S040203 巩留 土壤诱捕 XYS85.1 新源 土壤诱捕 XYS77.1 新源 土壤诱捕 P. gonapodyides XYW1710.1 新源 溪流诱捕 XYW1723.3 新源 溪流诱捕 XYS42 新源 土壤诱捕 P. lacustris XYW1701.3 新源 溪流诱捕 XYW1711.1 新源 溪流诱捕 XYW1724.4 新源 溪流诱捕  下载: 导出CSV
下载: 导出CSV
表 2 菌株接种叶片病斑大小
Table 2. The size of leaf spots inoculated by the strain
cm2 种名Species 菌株号Strain no. 苹果Apple 杏Apricot 山楂Hawthorn 核桃Walnut P. gregate GL1710.6 0±0 2.56±0.74 2.39±1.4 1.58±0.35 GL1710.9 0.03±0.03 2.35±0.06 0±0 8.57±0.77 P. sp. GLW1717.8 0±0 1.91±0.68 0.01±0.01 2.07±0.54 XYW1722.2 0.3±0.28 1.94±0.66 0±0 0.87±0.64 P. plurivora S040203 0.67±0.44 3.56±1.71 4.46±2.14 11.88±3.99 XYS85.1 0.4±0.07 4.9±2.36 2.42±0.72 13.12±2.34 XYS77.1 3.19±1.69 3.06±2.62 1.32±1.02 10.08±4.76 P. gonapodyides XYW1710.1 0.02±0.02 0.76±0.34 0.01±0.01 4.25±2.39 XYW1723.3 0.17±0.17 3.72±1.28 0.95±0.94 2.73±1.25 XYS42 0.04±0.01 3.41±0.29 0.5±0.15 7.41±5.21 P. lacustris XYW1701.3 1.28±0.86 3.26±1.8 2.51±1.51 4.02±0.63 XYW1711.1 1.56±0.19 5.31±0.68 2.21±0.85 4.45±0.76 XYW1724.4 1.83±0.72 2.12±0.7 2.21±0.27 5.73±4.29 对照 0.12±0.12 0±0 0.55±0.54 0.01±0.01
下载: 导出CSV
表 3 双因素方差分析表
Table 3. ANOVAs of effects of trees and Phytophthora on spots size
因素
Effect自由度
DFF值
FTrees 3 27.631*** Phytophthora 4 11.052** Residuals 12 Sigif. codes: ***0.001. **0.01
下载: 导出CSV
-
[1] 林培钧, 崔乃然.天山野果林资源——伊犁野果林综合研究[M].北京, 中国林业出版社. 2008:10. [2] Li L P, Hai Y, Anwar Moham.mat, et al. Community structure and conservation of wild fruit forests in the Ili valley, Xinjiang[J]. Arid Zone Research, 2011, 28(1):60-66. doi: 10.3724/SP.J.1148.2011.00060 [3] 淮稳霞.中国西南地区杜鹃——栎树林中疫霉菌的分离鉴定及快速检测技术研究[D].北京: 中国林业科学研究院, 2013: 1-3 [4] Wen-wen L, Wen-xia Z, Wen-xia H. Phytophthora pseudopolonica sp. nov., a new species recovered from stream water in subtropical forests of China[J]. International Journal of Systematic and Evolutionary Microbiology, 2017, 67(9):3666-3675. doi: 10.1099/ijsem.0.002254 [5] Huai W X, Tian G, Hansen, E M, et al. Identification of Phytophthora species baited and isolated from forest soil and streams in northwestern Yunnan province, China[J]. Forest Pathology, 2013, 43(2):87-103. doi: 10.1111/efp.12015 [6] Denman S, Kirk S A, Brasier C M, et al. In vitro leaf inoculation studies as an indication of tree foliage susceptibility to Phytophthora ramorum in the UK[J]. Plant Pathology, 2005, 54(4):512-521. doi: 10.1111/j.1365-3059.2005.01243.x [7] Milenkovic I, Ke a N, Karadž i c D, et al. Isolation and pathogenicity of Phytophthora species from poplar plantations in Serbia[J]. Forests, 2018, 9(6):330. doi: 10.3390/f9060330 [8] Jung T, Burgess T I. Re-evaluation of Phytophthora citricola isolates from multiple woody hosts in Europe and North America reveals a new species, Phytophthora plurivora sp. nov. Persoonia[J]. Molecular Phylogeny and Evolution of Fungi, 2009, 22(1):95-110. doi: 10.3767/003158509X442612 [9] Rytkönen A, Lilja A, Vercauteren A, et al. Identity and potential pathogenicity of Phytophthora species found on symptomatic Rhododendron plants in a Finnish nursery[J]. Canadian Journal of Plant Pathology, 2012, 34(2):255-267. doi: 10.1080/07060661.2012.686455 [10] Weiland J E, Nelson A H, Hudler G W. Aggressiveness of Phytophthora cactorum, P. citricola I, and P. plurivora from European beech[J]. Plant Disease, 2010, 94:1009-1014. doi: 10.1094/PDIS-94-8-1009 [11] Jankowiak R, Stepniewska H, Bilański P, et al. Occurrence of Phytophthora plurivora and other Phytophthora species in oak forests of southern Poland and their association with site conditions and the health status of trees[J]. Folia Microbiologica, 2014, 59(6):531-542. doi: 10.1007/s12223-014-0331-5 [12] Orlikowski L B, Trzewik A, Ptaszek M, et al. Relationship between source of water, occurrence, and pathogenicity of Phytophthora plurivora[J]. Acta Mycologica, 2012, 47(1):3-9. [13] Nechwatal J, Bakonyi, J, Cacciola, S O, et al. The morphology, behaviour and molecular phylogeny of Phytophthora taxon Salixsoil and its redesignationas Phytophthora lacustris sp.Nov[J]. Plant Pathology, 2012, 62(2):355-369. [14] Jung T, Stukely M J C, Hardy G E S, et al. Multiple new Phytophthora species from ITS Clade 6 associated with natural ecosystems in Australia:evolutionary and ecological implications[J]. Persoonia-Molecular Phylogeny and Evolution of Fungi, 2011, 26(1):13-39. doi: 10.3767/003158511X557577 [15] Belhaj R, McComb J, Burgess T I, et al. Pathogenicity of 21 newly described Phytophthora species against seven Western Australian native plant species[J]. Plant Pathology, 2018, 67(5):1140-1149. doi: 10.1111/ppa.12827 [16] Corcobado T, Cubera E, Pérez-Sierra A, et al. First report of Phytophthora gonapodyides involved in the decline of Quercus ilex in xeric conditions in Spain[J]. New Disease Reports, 2010, 22:33. doi: 10.5197/j.2044-0588.2010.022.033 -

 点击查看大图
点击查看大图

图(4) / 表(3)
计量
- 文章访问数: 3842
- HTML全文浏览量: 1936
- PDF下载量: 32
- 被引次数: 0
