

 下载:
下载:

-
延迟开放(Serotiny)是木本植物在火灾环境下的一种常见特征和生活史策略,它表现为种子在冠层封闭的球果中保留了1个以上的繁殖周期[1]。研究发现,目前有1 000余种植物存在种子延迟开放现象,其中,普遍认为松属(Pinus Linn.)植物是延迟开放的代表[1],大量关于延迟开放的研究是基于松属植物开展的[2-5]。
具有延迟开放性状的树种林冠中储存了大量成熟和健康的多年宿存的种子,只有在火灾后才会迅速打开,以提高森林的火灾恢复能力,使森林在火灾后能够迅速恢复[6]。虽然大量研究表明,高温可以促进延迟开放的球果打开[7-10],但是球果延迟开放背后的机制尚不明确。
松树成熟的球果鳞片是由死细胞组成,不具有自主运动的能力[11]。在电子显微镜观察下,发现鳞片由2层组织组成,鳞片的内表面由纤维组织构成,成束排列,鳞片的外表面由厚壁细胞组成[12-13]。二者的拉伸刚度和吸湿膨胀系数都存在显著差异,水分的变化引起纤维运动,从而导致鳞片张开或闭合[11, 14]。延迟开放的球果是由于鳞片中有松脂,松脂作为一种粘合材料起到密封球果不让内部水分丢失,使球果成熟后保持长期闭合。有研究表明,松脂粘合力消失的温度大概在45~50℃[15-17],高温可能使松脂粘合力消失,从而导致球果打破延迟开放,但是松脂含量与延迟开放之间的关系尚不清楚。
云南松(P. yunnanensis Franch.)、高山松(P. densata Mast.)、思茅松(P.kesiya var. langbianensis Gaussen.)以及马尾松(P.massoniana Lamb.)是中国西南地区4种常见松属植物,也是常见的采脂树种[18],其中,云南松是我国西南地区的特有种,以云南高原为起源和分布中心,在四川、贵州、广西、西藏均有分布,在云南西北横断山区与高山松毗连,在云南西南部则与思茅松邻接[19]。云南松种群具有丰富的多态性、突出的生态地理变异以及明显的种群间多型性,其变种有生态小种地盘松(P. yunnanensis var. pygmaea Hsueh.)和地理小种细叶云南松(P. yunnanensis var. tenuifolia Cheng et Law.)[20]。已有研究表明,云南松是延迟开放树种[5],其球果在枝头宿存多年后内部种子仍有较高的萌发率,且高温烘烤会打破云南松球果的延迟开放[10]。
-
本研究以云南松的3个变种(云南松原种、地盘松、细叶云南松)和马尾松、思茅松、高山松为研究对象,每个物种选取4~6个地点,调查其延迟开放度并测定球果鳞片松脂含量,旨在探讨松属植物球果鳞片松脂含量对球果延迟开放的影响,进一步加深对延迟开放的认识,丰富松属植物延迟开放的研究。
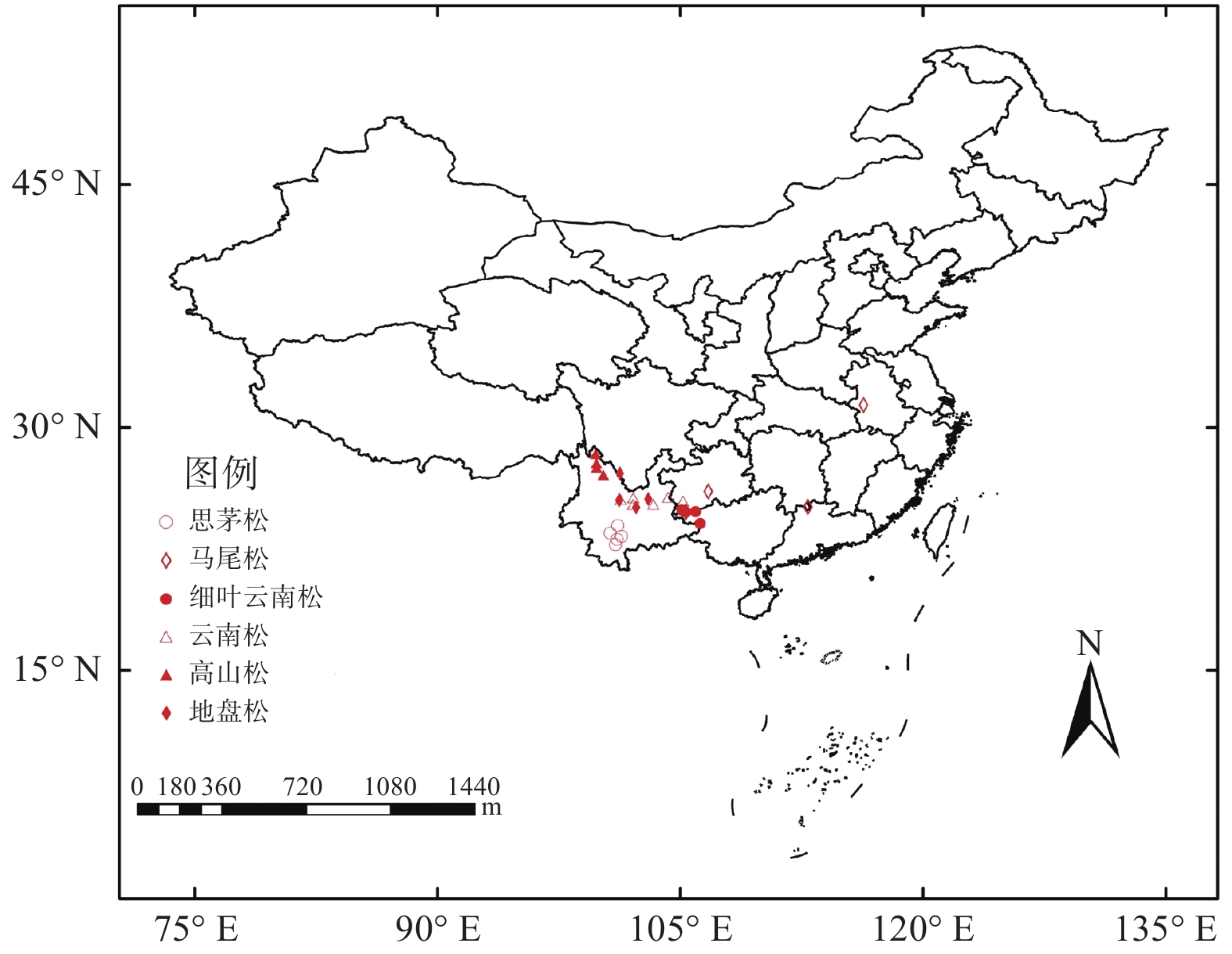
本研究选取了6个物种共28个地点进行松树延迟开放度的调查和当年闭合球果的采样(图1)。云南松选取了云南省内富源、旧县、兴仁、武定、姚安和禄丰6个地点;地盘松选取了勤丰、格萨拉、牟定新桥和寻甸潘所海4个地点;细叶云南松选取了兴义天生桥、册亨县弼佑乡、隆林革布乡、万峰湖和雅长5个地点;思茅松选取了墨江通关镇、宁洱、普洱、景谷和镇沅5个地点;高山松选取了小中甸、格咱乡、普达措和丽江4个地点;马尾松选取了广西隆林、贵州贵阳、湖南宜章和安徽霍山4个地点。
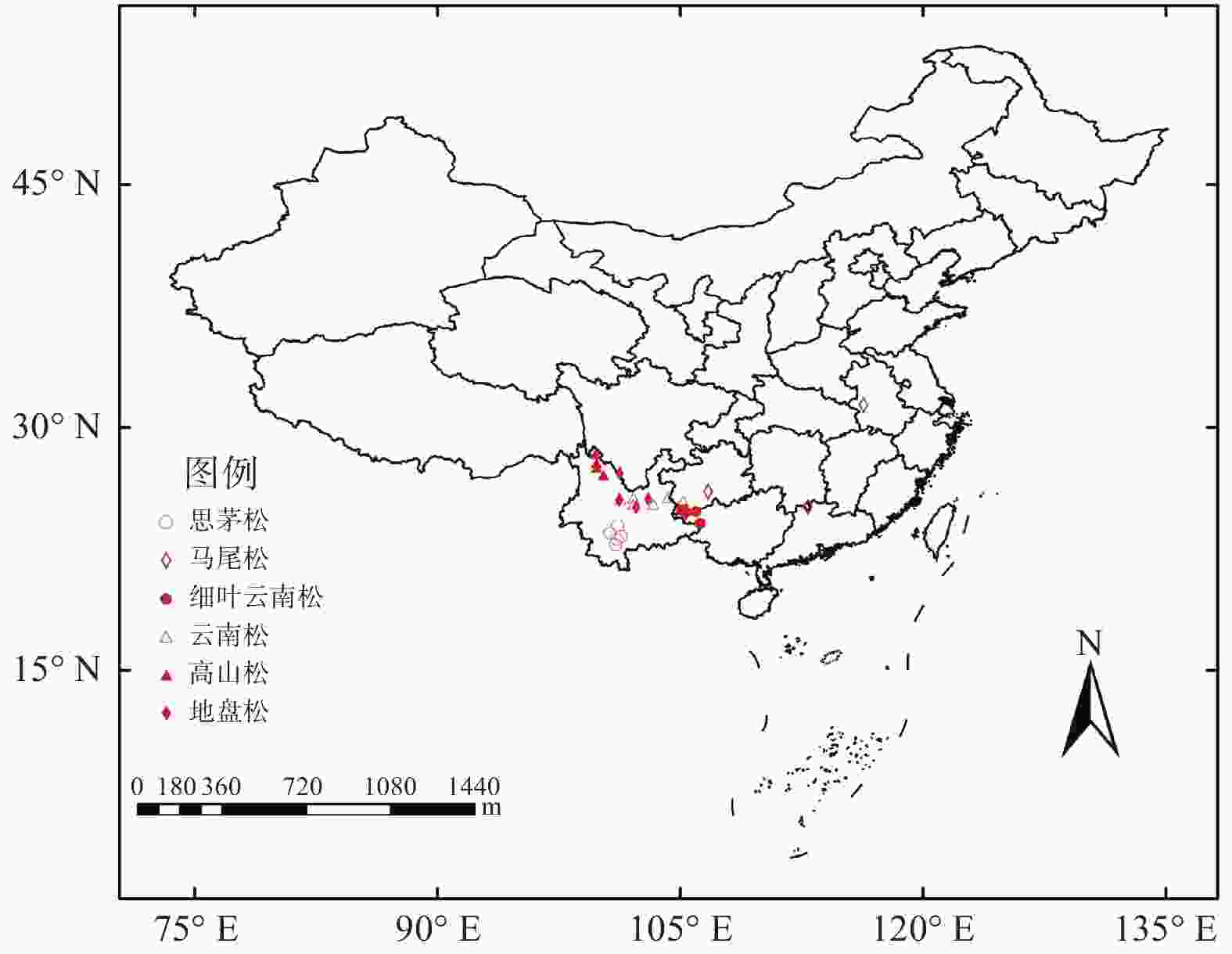
图 1 采样分布图
Figure 1. Distribution of sampling sites
-
云南松、地盘松、细叶云南松和高山松主干和侧枝的顶部每年只分化生长发出1段新枝,马尾松每年生长1段(广东2段),思茅松2 a分化出1段新枝,新枝和上年生长的老枝之间会形成一个较明显的环状节,球果着生在节上。因此,可依据球果着生节的位置推算出结实年份和宿存延迟开放时间。
-
球果成熟后继续留在树枝上的闭合球果的数量占这棵树产生的所有球果数量的比例就是延迟开放度[21]。于2017年10月,每个地点选取30棵树(每棵树至少间隔50 m),计数每棵树闭合球果(未成熟的球果不进行统计,因为无法确定其成熟后立即开放或是延迟开放)和开放球果的数量,估算球果的延迟开放度。
-
每个地点选取10颗当年成熟的球果,剥下约1~2 g鳞片,剪碎后放入质量为m0的茶袋中;再将样品放入−60℃超低温冰箱冷冻12 h,使鳞片内水分结晶,之后放入冷冻干燥机中冻干24 h后,称质量为m1;将样品放入乙醚-乙醇(3:1)混合液浸泡12 h后,再以乙醚-乙醇混合液进行索氏提取[22-23],30 min 1次循环,循环20次后,放入烘箱100℃烘至恒定质量,记为m2,利用残差法[24]测定松脂含量为: Cr=(m1−m2)/(m1−m0)×100%
-
试验数据先用Excel进行简单整理后作图,IBM SPSS Statistics在95%水平上进行单因素方差(ANOVA)分析,Duncan检验法进行差异显著性分析。
-
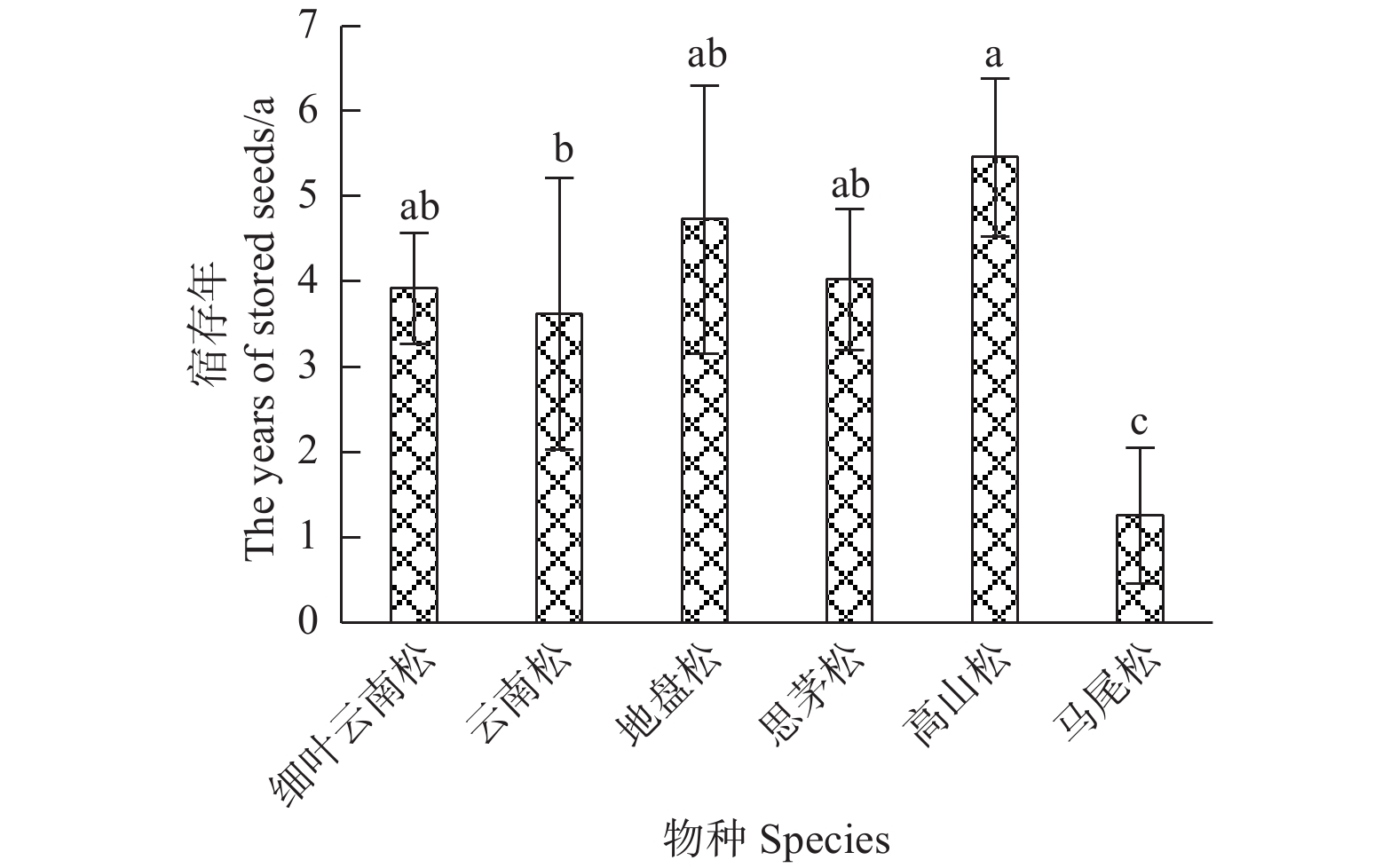
球果宿存是其延迟开放的基础。调查结果(图2)发现:几个物种的宿存年分别为高山松(5.46±0.93) a、地盘松(4.73±1.56) a、思茅松(4.03±0.83) a、细叶云南松(3.92±0.65) a、云南松(3.63±1.59) a、马尾松(1.26±0.80) a。可以看出:宿存年在物种间差异显著,其中,高山松的宿存年最高,马尾松的宿存年最低(F=6.29,P=0.001)。
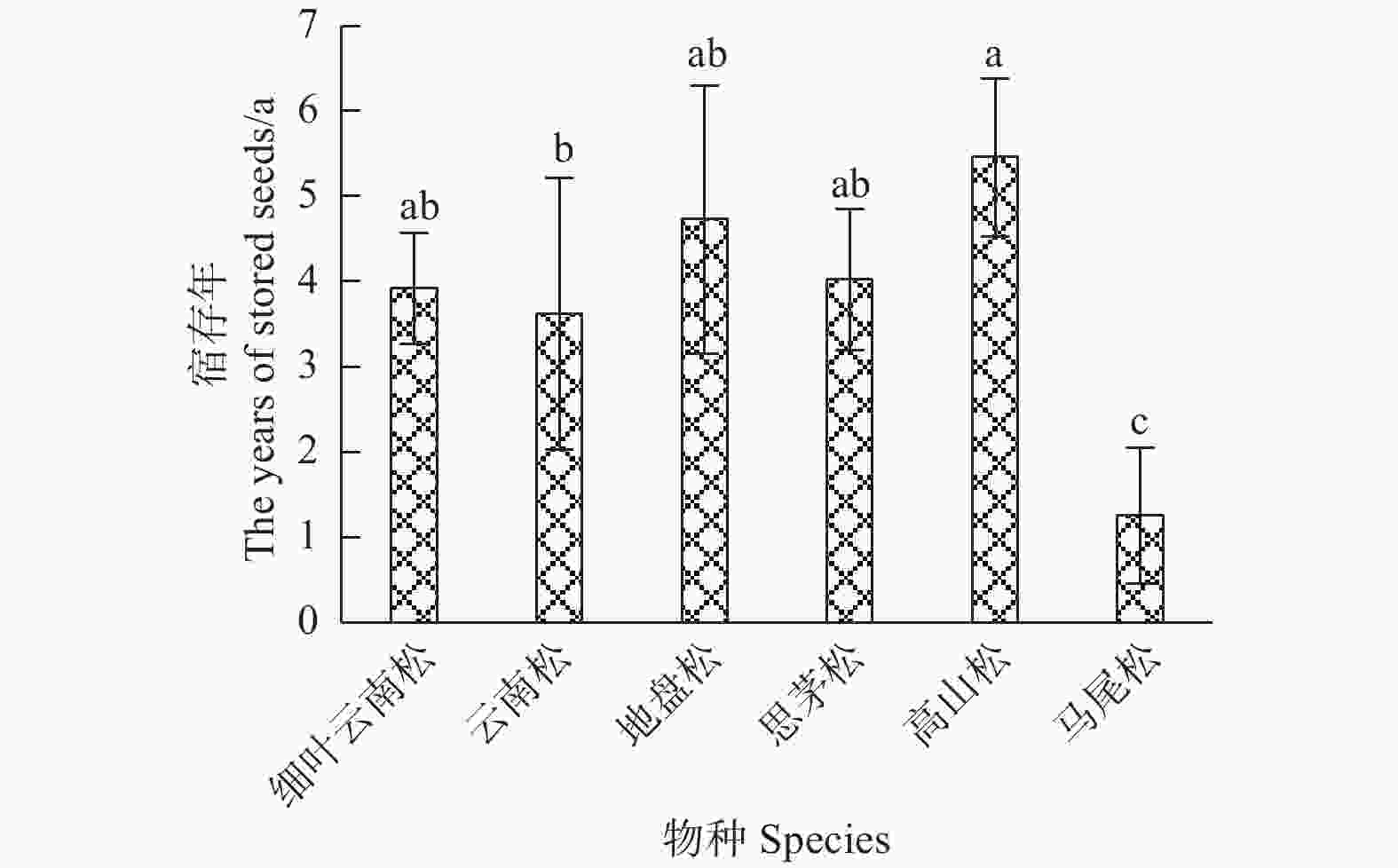
图 2 不同松树球果宿存年差异性比较
Figure 2. The dfference comparison of the years of stored seeds in pines
-
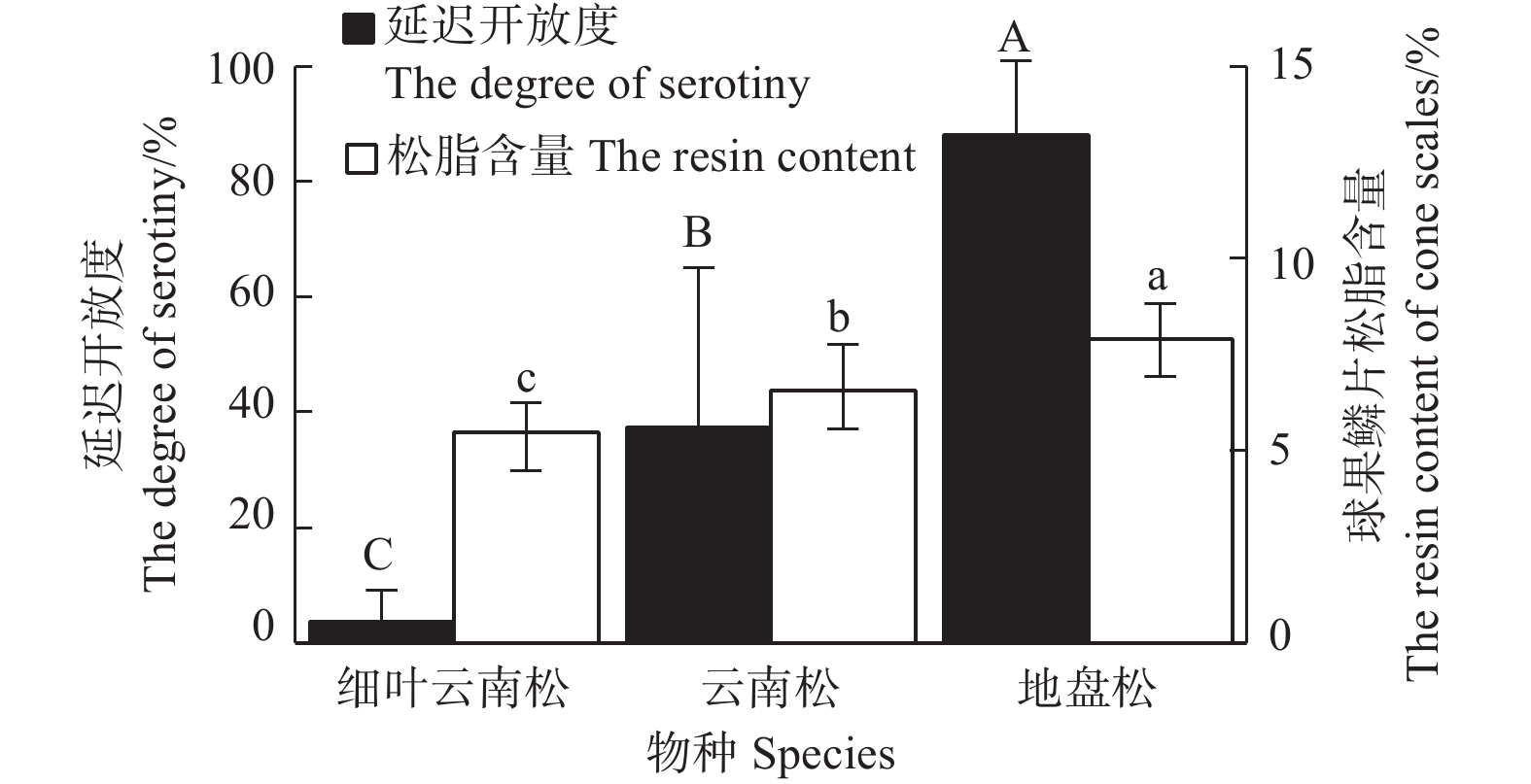
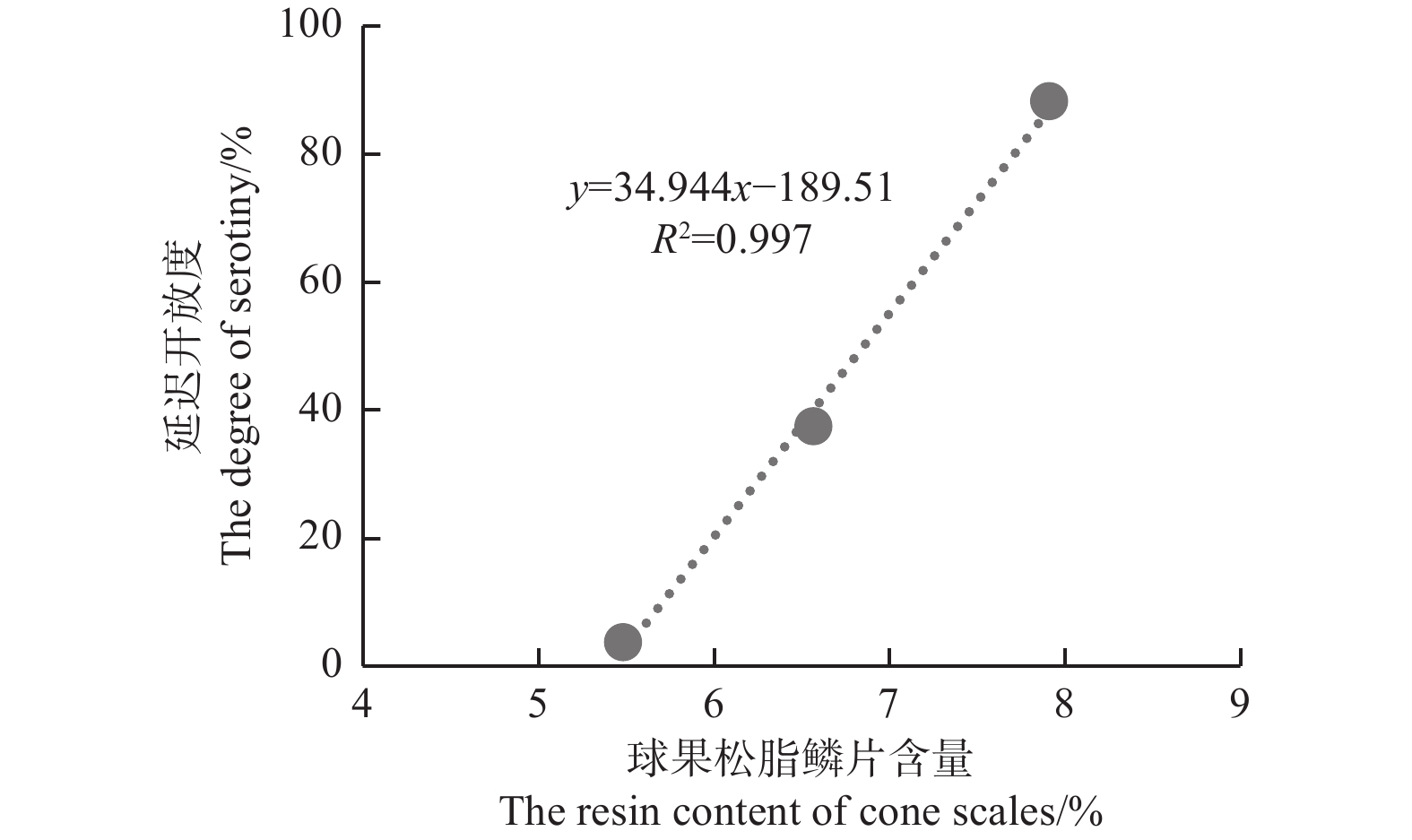
图3表明:云南松、地盘松、细叶云南松的球果延迟开放度差异极显著(F=21.58,P<0.001),地盘松(88.25±12.65)%>云南松(37.35±27.71)%>细叶云南松(3.64±5.43)%;云南松、地盘松、细叶云南松的球果鳞片松脂含量也差异显著(F=6.73,P=0.011),地盘松(7.91%±0.90%)%>云南松(6.57±1.18)%>细叶云南松(5.48±0.76)%。由图4可看出:球果延迟开放度总体上随着鳞片松脂含量的增加而增大,二者呈显著正相关(R2=0.997,P=0.036)。
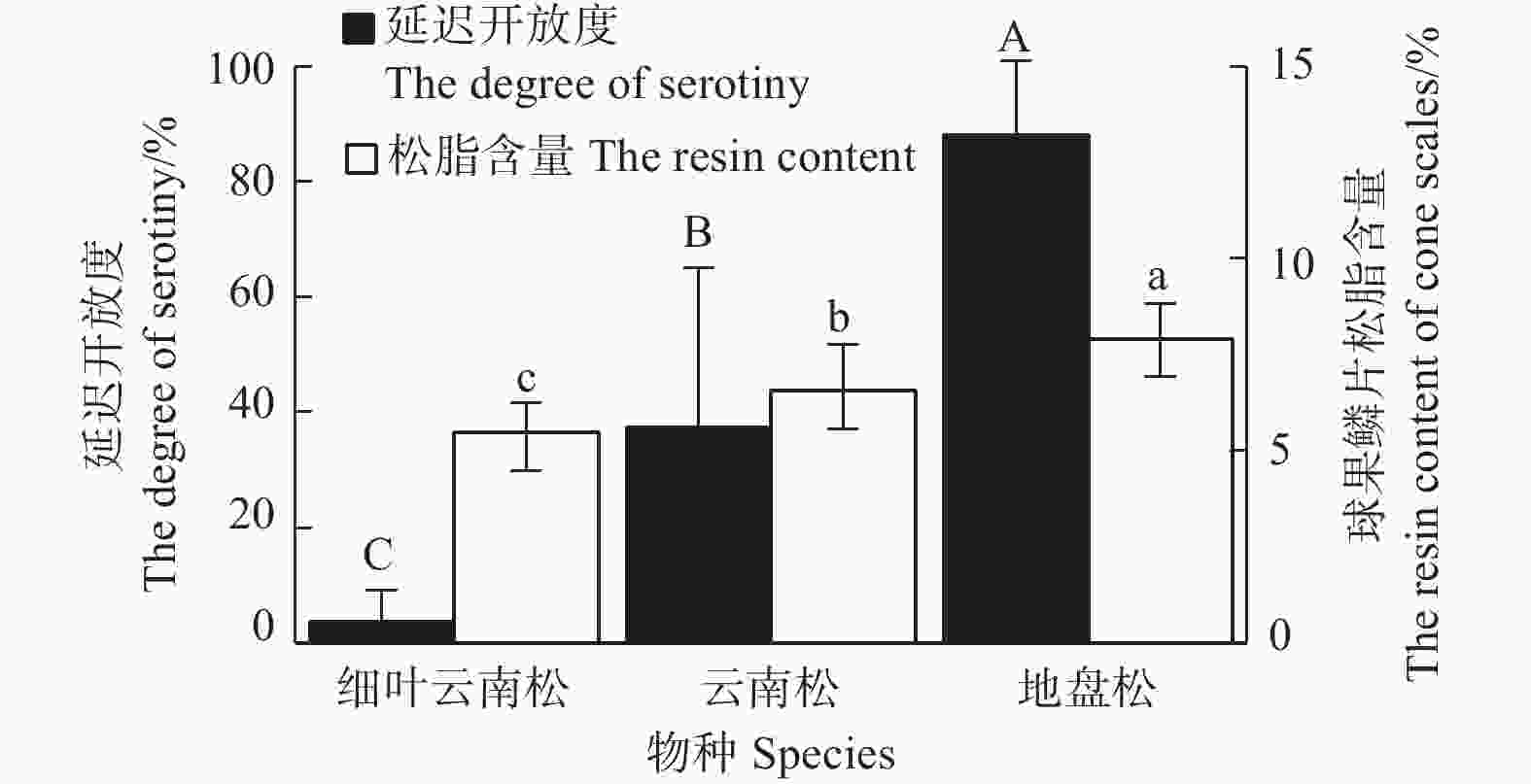
图 3 云南松变种内延迟开放度和球果鳞片松脂含量
Figure 3. The degree of serotiny and the resin content of cone scales in P. yunnanensis varieties

图 4 云南松3个变种间延迟开放度与球果鳞片松脂含量的关系
Figure 4. The relationship between the degree of serotiny and the resin content of cone scales in P. yunnanensis varieties
-
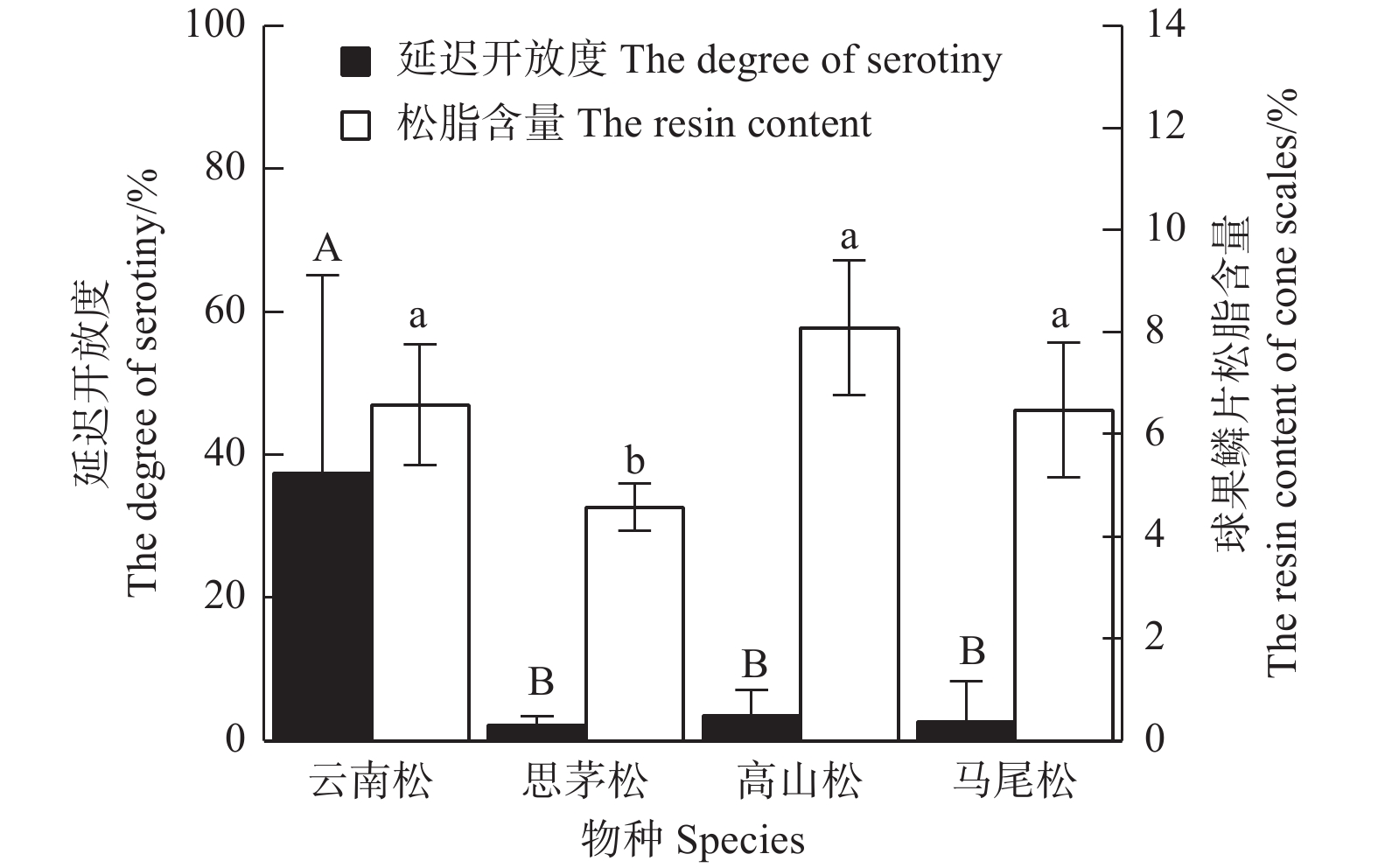
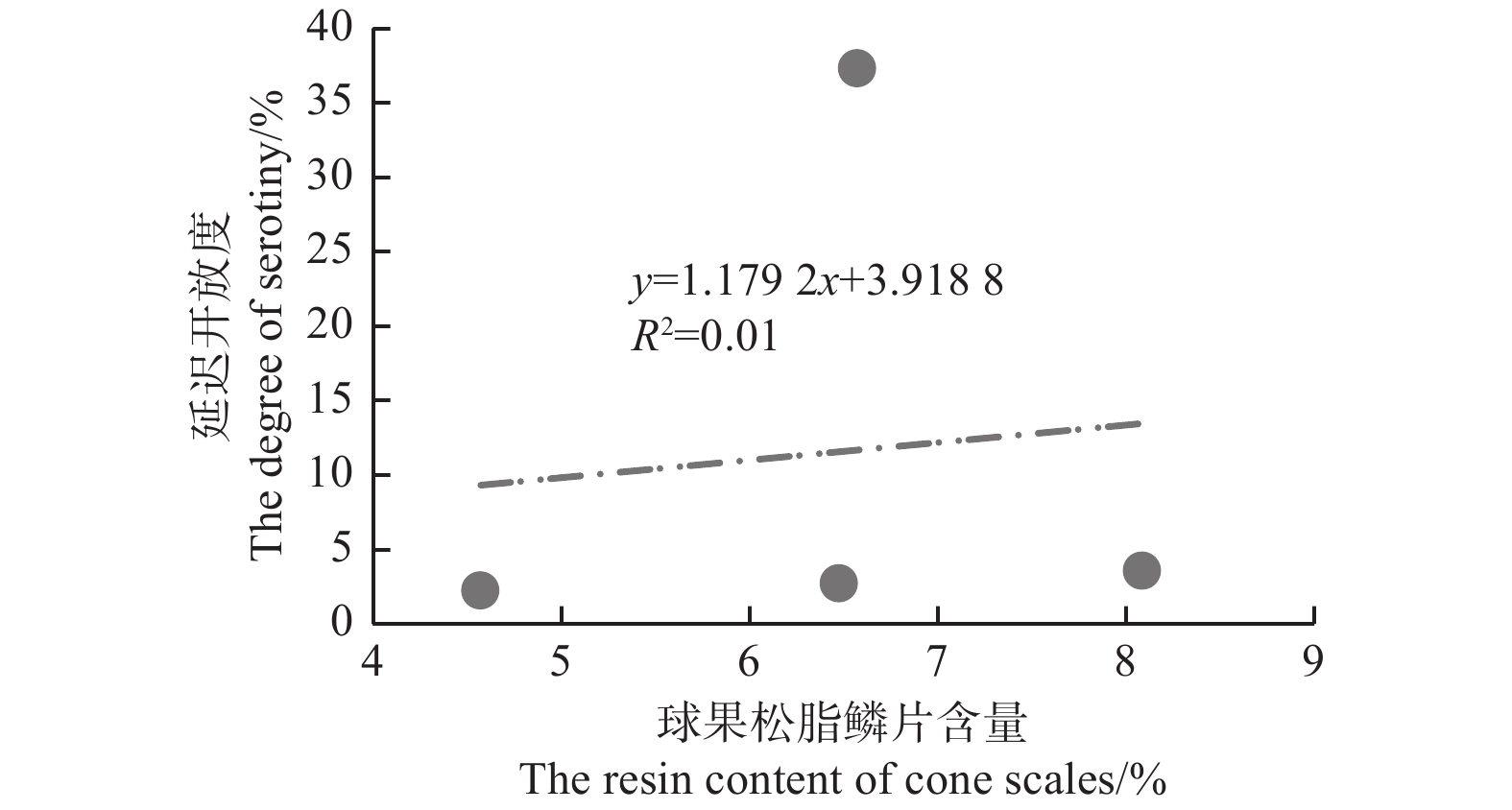
图5表明:在中国西南4种常见的松树中,云南松的延迟开放度最高,为(37.35±27.71)%,极显著 (F=6.162,P=0.006) 高于思茅松(2.26±1.16)%、马尾松(2.76±5.53)%和高山松(3.62±3.47)%,而思茅松、马尾松和高山松之间无差异;4种松树中,思茅松的球果鳞片松脂含量最低(4.57±0.46)%,显著 (F=7.721,P=0.002) 低于云南松(6.75±1.18)%、高山松(8.09±1.32)%和马尾松(6.48±1.32)%,而云南松、马尾松、高山松的球果鳞片松脂含量差异不显著。图6表明:4种松树间,延迟开放度与球果鳞片松脂含量并没有显著的正相关性(R2=0.01,P=0.902)。
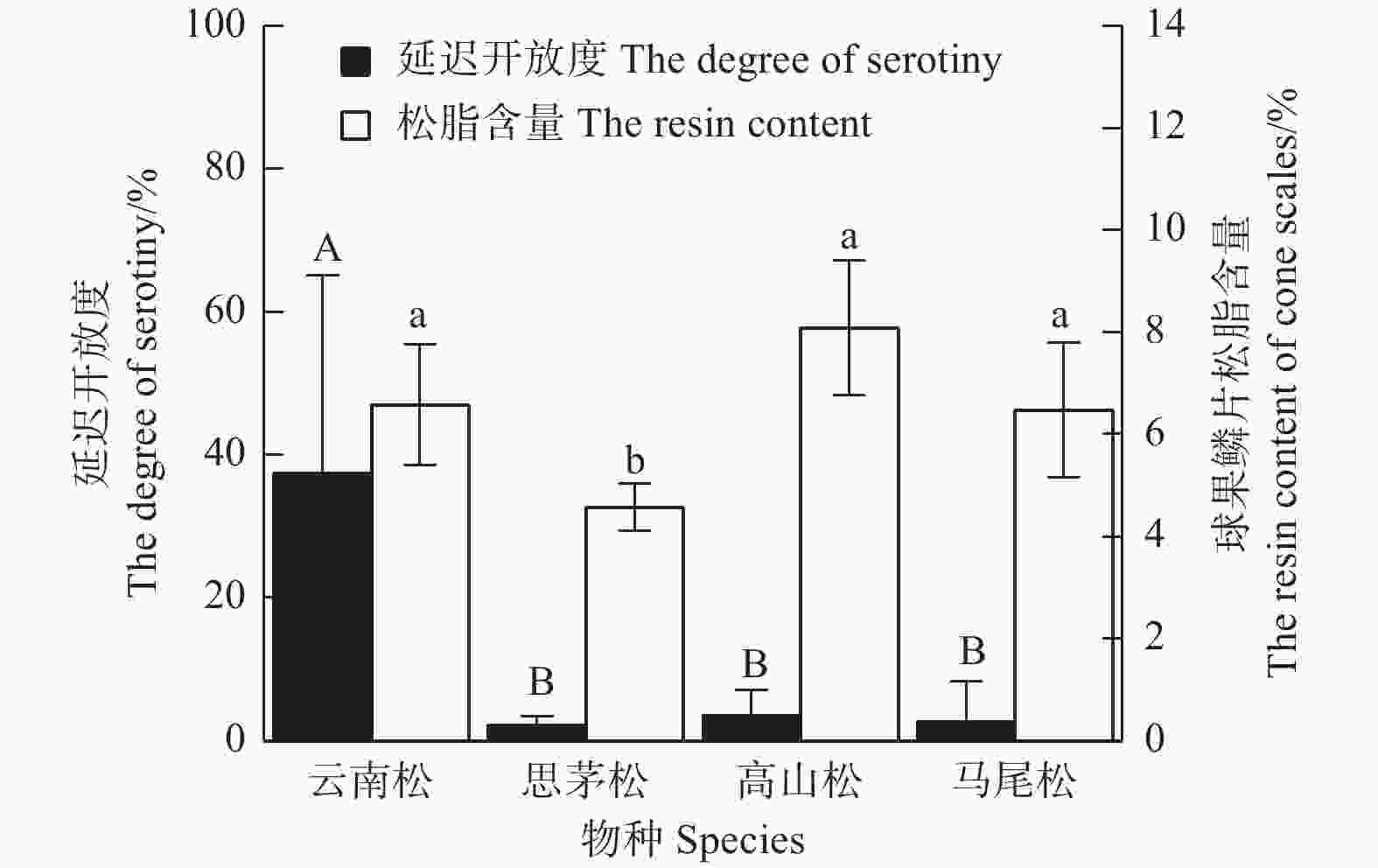
图 5 四种松树的延迟开放度和球果鳞片松脂含量
Figure 5. The degree of serotiny and the resin content of cone scales in 4 pines
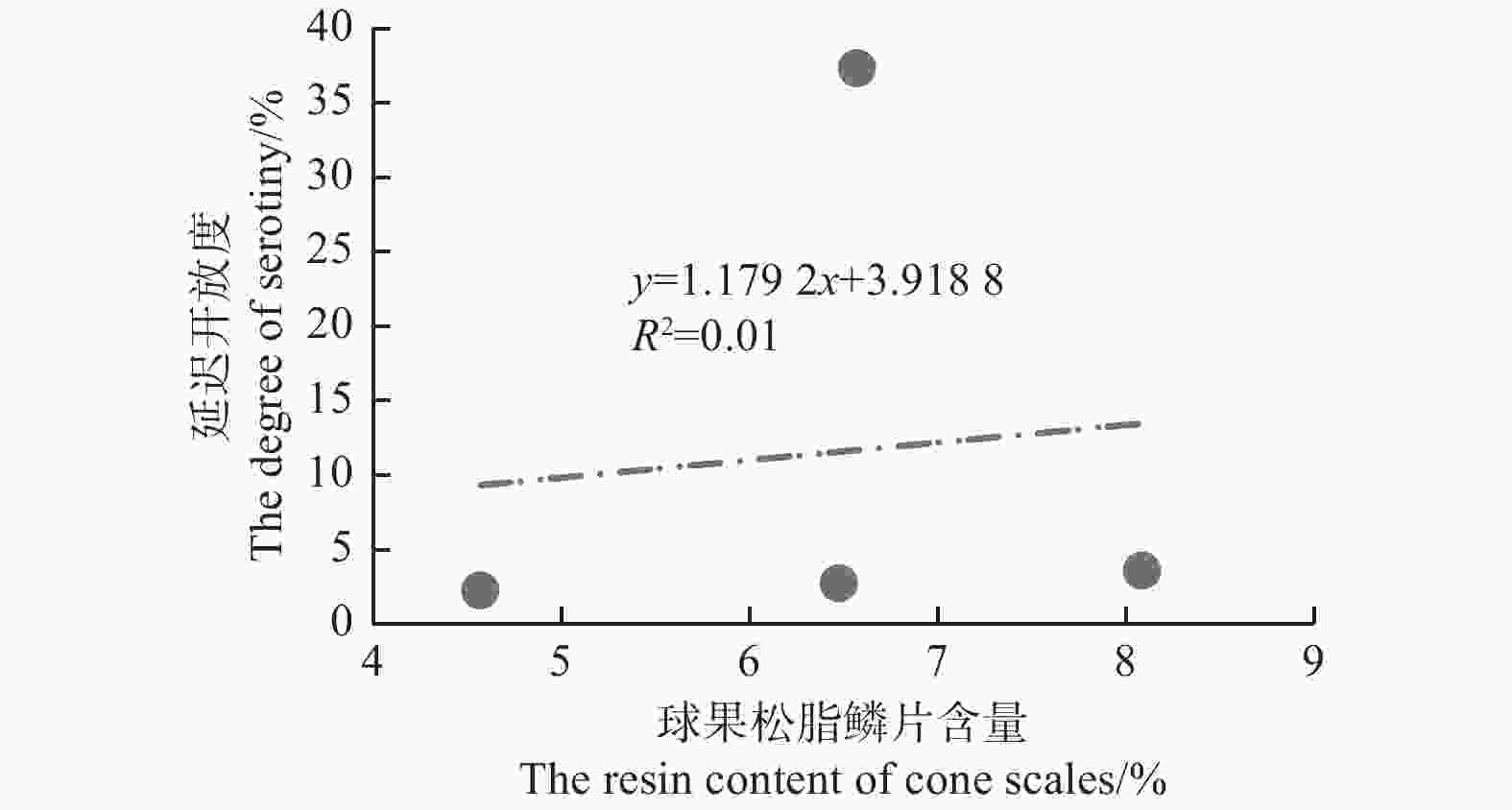
图 6 四种松树的延迟开放度和球果鳞片松脂含量的关系
Figure 6. The relationship between the degree of serotiny and the resin content of cone scales in 4 pines
-
不考虑种间及种内的影响,将28个松树种群的延迟开放度与球果鳞片松脂含量做散点图。由图7可知:球果鳞片松脂含量为3.9%~9.38%,可以观察到,松脂含量与球果延迟开放度不具有线性关系(R2=0.182,P=0.024);但当球果鳞片松脂含量小于6%时,延迟开放度极低(0%~6.05%),即无论何种松树物种、哪个种群,当松脂含量低于6%时,种群的延迟开放水平都极低,表现为非延迟开放种群;当松脂含量大于6%时,延迟开放度变化较大,为0%~100%。
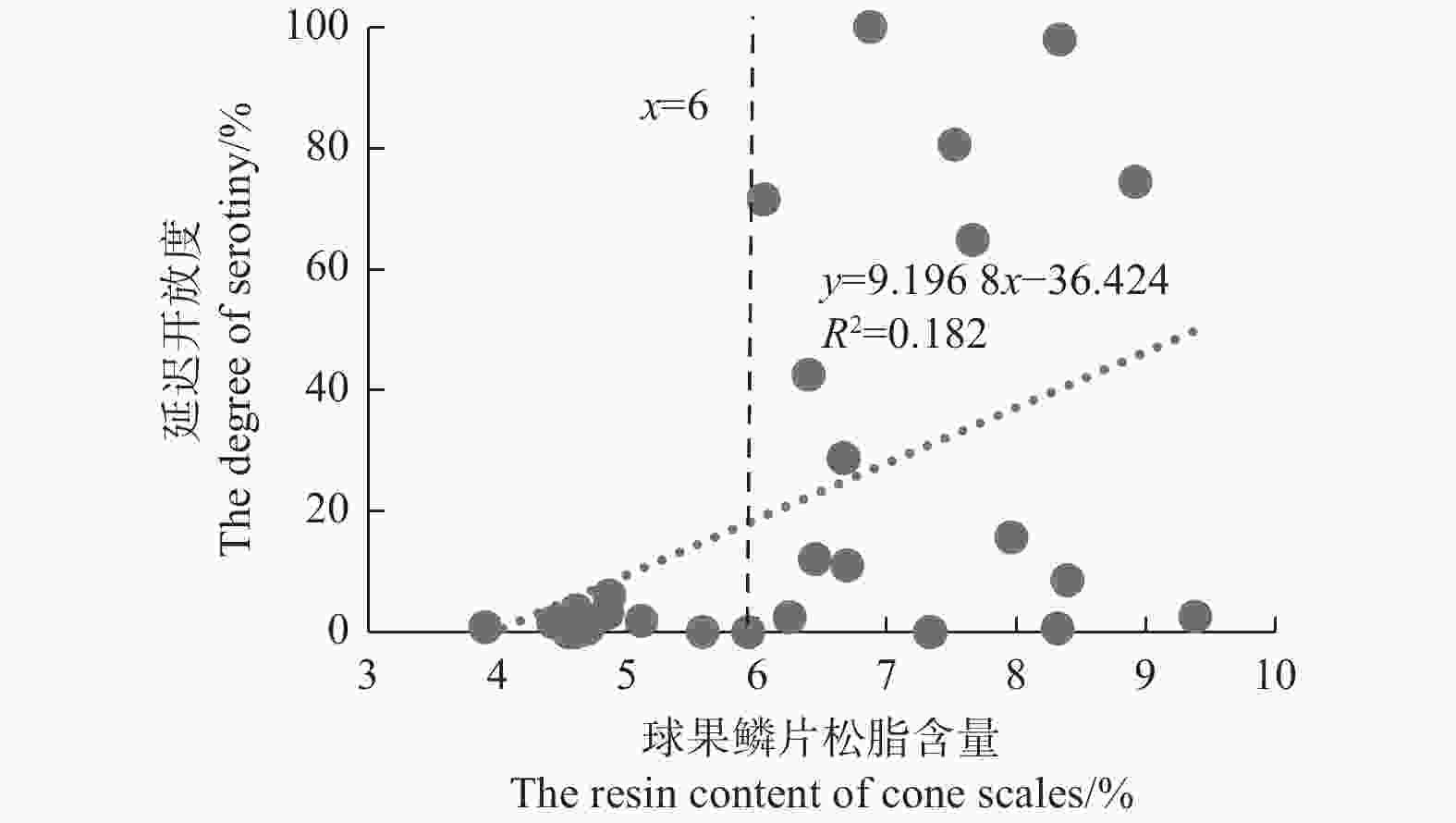
图 7 28个样地球果鳞片松脂含量和延迟开放度的散点图
Figure 7. Scatter plots of the degree of serotinyand the resin content of cone scales of 28 sites
-
延迟开放是木本植物适应火灾的一种生活史对策,而球果宿存是球果延迟开放的基础。调查发现,球果宿存年极低的马尾松(图2)其延迟开放度也表现的极低(2.76±5.53)%(图5),而其他几个种都具有3~6 a的宿存(图2)。云南松和地盘松有较高的延迟开放度,而细叶云南松、思茅松、高山松基本没有延迟开放(图3、图5),说明宿存在枝条上的球果,还受其他因素影响,选择开放或者闭合。
有研究报道,球果延迟开放已经独立进化了多次[25-26],且在基因层面由多基因控制[27-28]。松脂是松属植物分泌的树脂,具有黏性,常用于制作粘合剂[18]。许多研究认为,松脂作为粘合材料可以使球果鳞片密封、种子延迟开放[15-17]。Kossuth等[29]将球果鳞片的外层部分剥下,分别测定鳞片外层和内部的松脂含量,认为鳞片外层松脂含量的增加可能有助于保持球果在秋季脱水和开放前闭合。本研究直接测定整个鳞片的松脂含量,发现延迟开放度与球果鳞片松脂含量的关系在种内水平与种间水平具有不同的表现。
在种群水平上(图7),不同种群球果鳞片松脂含量与延迟开放度虽然不具有线性相关(R2=0.182,P=0.024),但松脂的含量却对延迟开放有着决定性作用。当球果鳞片松脂含量低于6%时,球果基本没有延迟开放现象;当球果鳞片松脂含量高于6%时,这时虽然松脂含量满足密封球果的条件,但可能受环境或者松脂理化性质如松脂熔点、松脂粘合力大小的影响,不同松种的延迟开放度表现极大的波动[16, 30]。高山松的球果鳞片松脂含量虽然高达(8.09±1.32)%,但它的延迟开放度仅有(3.62±3.47)%。其球果鳞片松脂含量高可能是因为高山松是云南松和油松的天然杂交种[31]。打破延迟开放需要高温,而高山松可能因为分布区海拔高、温度低,火灾相对很难发生,延迟开放对其繁殖反而是不利的[32],所以,高山松表现较低的延迟开放度。
云南松属于延迟开放物种,云南松3个变种的(图4)球果鳞片松脂含量与延迟开放度具有正相关关系(R2=0.997,P=0.036),除松脂含量,变种间松脂其他性状(松脂熔点、树脂道等)的基因差异很小,球果鳞片松脂含量影响着延迟开放度的大小[33];但在物种水平(图6)上,可能由于松脂各个性状的基因在物种间都存在较大差异,球果鳞片松脂含量与延迟开放度上不呈线性相关(R2=0.01,P=0.902)。此外,延迟开放还受环境的影响,过火[34]、气候[30]、林分结构[15]等都会使延迟开放度产生很高的差异性。此次种间采样分布区域跨度过大,气候差异大,故气候等环境因素的差异也可能弱化了球果鳞片松脂含量和延迟开放度的种间关系。
-
球果宿存是延迟开放的基础,球果鳞片松脂含量低于6%的松属植物基本没有延迟开放。在具有延迟开放的云南松种内的3个变种间,球果鳞片松脂含量与其延迟开放度呈正相关;但在物种水平,球果鳞片松脂含量与其延迟开放度并无线性关系。在以后的研究中,除球果鳞片松脂含量外,还应综合考虑球果形态、鳞片结构、鳞片树脂道、球果松脂熔点、球果松脂粘合力大小等对松属植物球果延迟开放的影响。
中国西南几种松树球果鳞片松脂含量对球果延迟开放的影响
Effect of Resin Content in Cone Scales on Cone Serotiny in Pines in Southwest China
-
摘要:
目的 探究松树球果延迟开放的机制,讨论松树球果鳞片松脂含量对延迟开放的影响。 方法 以云南松、地盘松、细叶云南松、高山松、马尾松和思茅松为研究对象,调查球果宿存年、延迟开放度。采集当年生球果并测定球果鳞片中松脂含量,分析松树球果鳞片松脂含量与延迟开放度的相关性。 结果 球果宿存是延迟开放的基础,当球果鳞片松脂含量低于6%时的物种,其球果没有延迟开放现象。在具有延迟开放的云南松种内的3个变种,其球果鳞片松脂的含量与其延迟开放度呈正相关(R2=0.997,P=0.036),但在种间水平,球果鳞片松脂含量与其延迟开放度并无线性关系(R2=0.01,P=0.902),推测是由于环境和基因的差异较大所致。 结论 球果鳞片松脂含量对球果延迟开放起重要作用,含量过低则没有延迟开放;在具有延迟开放的云南松变种内,球果鳞片松脂含量显著影响其延迟开放程度。 Abstract:Objective To investigate the effect of resin content of pine cone scales on serotiny. Method Cone persistent ages, the degree of serotiny and the resin content of current-year cone scales in six pine species (Pinus yunnanensis Franch., P. yunnanensis var. pygmaea Hsueh., P. yunnanensis var. tenuifolia Cheng et Law., P. densata Mast., P. massoniana Lamb., and P. kesiya var. langbianensis Gaussen.) were investigated, and the correlation between resin content of cone scales and the degree of serotiny were analyzed. Result For the species whose resin content of cone scales less than 6%, the persistence of cones, which is the indication of serotiny, was not observed. In P. yunnanensis varieties characterized by serotiny, the resin content of cone scales was positively related to the degree of serotiny (R2=0.997, P=0.036). However, the relationship between the cone scale resin content and the degree of serotiny was not significant (R2=0.01, P=0.902) across these taxa, which may be due to the large genetic and environmental differences. Conclusion The results indicate that the resin content of cone scales plays an important role in pine serotiny and there will be no serotiny when it is too low. In P. yunnanensis varieties with serotiny, the serotiny degree is effected by the resin content of cone scales. -
Key words:
- pine; persistent
- / serotiny
- / cone scales
- / resin content
-
图 2 不同松树球果宿存年差异性比较
Figure 2. The dfference comparison of the years of stored seeds in pines
图 3 云南松变种内延迟开放度和球果鳞片松脂含量
Figure 3. The degree of serotiny and the resin content of cone scales in P. yunnanensis varieties
图 4 云南松3个变种间延迟开放度与球果鳞片松脂含量的关系
Figure 4. The relationship between the degree of serotiny and the resin content of cone scales in P. yunnanensis varieties
图 5 四种松树的延迟开放度和球果鳞片松脂含量
Figure 5. The degree of serotiny and the resin content of cone scales in 4 pines
图 6 四种松树的延迟开放度和球果鳞片松脂含量的关系
Figure 6. The relationship between the degree of serotiny and the resin content of cone scales in 4 pines
-
[1] Lamont B B, Maitre D C L, Cowling R M, et al. Canopy seed storage in woody plants[J]. Botanical Review, 1991, 57(4): 277-317. doi: 10.1007/BF02858770 [2] Tapias R, Gil L, Fuentes-Utrilla P, et al. Canopy seed banks in Mediterranean pines of southeastern Spain: a comparison between Pinus halepensis Mill., P. pinaster Ait., P. nigra Arn. and P. pinea L.[J]. Journal of Ecology, 2001, 89(4): 629-638. doi: 10.1046/j.1365-2745.2001.00575.x [3] Schoennagel T, Turner M G, Romme W H. The influence of fire interval and serotiny on postfire lodgepole pine density in Yellowstone National Park[J]. Ecology, 2003, 84(11): 2967-2978. doi: 10.1890/02-0277 [4] Vincenzi S, Piotti A. Evolution of serotiny in maritime pine (Pinus pinaster) in the light of increasing frequency of fires[J]. Plant Ecology, 2014, 215(7): 689-701. [5] Su W, Yu J, Zhang G, et al. Comparison of the canopy and soil seed banks of Pinus yunnanensis in central Yunnan, China[J]. Forest Ecology and Management, 2019, 437: 41-48. doi: 10.1016/j. foreco. 2019.01.002. [6] Ne'eman G, Goubitz S, Ran N. Reproductive traits of Pinus halepensis in the light of fire – a critical review[J]. Plant Ecology, 2004, 171(1-2): 69-79. [7] Givnish T J. Serotiny, geography, and fire in the pine barrens of New Jersey[J]. Evolution, 1981, 35(1): 101-123. doi: 10.1111/j.1558-5646.1981.tb04862.x [8] Hellum A K, Pelchat M. Temperature and time affect the release and quality of seed from cones[J]. Canadian Journal of Forest Research, 1979, 9(2): 154-159. doi: 10.1139/x79-029 [9] Reyes O, Casal M. Effect of high temperatures on cone opening and on the release and viability of Pinus pinaster and P. radiata seeds in NW Spain[J]. Annals of Forest Science, 2002, 59(3): 327-334. doi: 10.1051/forest:2002028 [10] 苏文华, 崔凤涛, 赵元蛟, 等. 云南松球果延迟开放及其植冠种子库[J]. 生态学报, 2017, 37(2):541-548. [11] Dawson C, Vincent J F V, Rocca A M. How pine cones open[J]. Nature, 1997, 390: 668. doi: 10.1038/37745. [12] Harlow W M, Côté W A, Day A C. The opening mechanism of pine cone scales[J]. Journal of Forestry, 1964, 62(8): 538-540. doi: 10.1093/jof/62.8.538 [13] Song K, Chang S-S, Lee S J. How the pine seeds attach to/detach from the pine cone scale?[J]. Frontiers in life science, 2017, 10(1): 38-47. doi: 10.1080/21553769.2017.1287777 [14] Song K, Yeom E, Seo S-J, et al. Journey of water in pine cones[J]. Scientific Reports, 2015, 51: 9963. doi: 10.1038/srep09963. [15] Crossley D I. Effect of crown cover and slash density on the release of seed from slash-borne lodgepole pine cones[R]. Forest Research Division Technical Note 41. Ottawa: Canada Department of Northern Affairs and National Resources, Forestry Branch, 1956. [16] Huss J C, Spaeker O, Gierlinger N, et al. Temperature-induced self-sealing capability of Banksia follicles[J]. Journal of the Royal Society Interface, 2018, 15(143). doi: 10.1098/rsif. 2018.0190. [17] Clements F E. The life history of lodgepole burn forests[M]. US Dept. of Agriculture, Forest Service. Washington: Government Printing Office, 1910. [18] 宋湛谦. 中国松脂特征与松属分类[M]. 北京: 中国林业出版社, 1998. [19] 陈 飞, 王健敏, 孙宝刚, 等. 云南松的地理分布与气候关系[J]. 林业科学研究, 2012, 25(2):163-168. [20] 黄瑞复. 云南松的种群遗传与进化[J]. 云南大学学报: 自然科学版, 1993, 15(1):50-63. [21] Goubitz S, Nathan R, Roitemberg R, et al. Canopy seed bank structure in relation to: fire, tree size and density[J]. Plant Ecology, 2004, 173(2): 191-201. [22] 魏 红, 钟红舰, 汪 红. 索氏抽提法测定粗脂肪含量的改进[J]. 中国油脂, 2004, 29(6):52-54. [23] 谭真真, 王 雁, 李 亚. 芍药种子含油率与脂肪酸成分研究[J]. 林业科学研究, 2018, 31(3):45-50. [24] 罗 凡, 费学谦, 车运舒, 等. 香叶树挥发油、油脂等主要成分分析[J]. 林业科学研究, 2015, 28(2):284-288. [25] Eva G, Marcel R, Sanderson M J, et al. Evolution of genome size in pines (Pinus) and its life-history correlates: supertree analyses[J]. Evolution, 2004, 58(8): 1705-1729. doi: 10.1111/j.0014-3820.2004.tb00456.x [26] Tianhua H, Pausas J G, Belcher C M, et al. Fire-adapted traits of Pinus arose in the fiery cretaceous[J]. New Phytologist, 2012, 194(3): 751-759. doi: 10.1111/j.1469-8137.2012.04079.x [27] Budde K B, Heuertz M, Hernández-Serrano A, et al. Insitu genetic association for serotiny, a fire-related trait, in Mediterranean maritime pine (Pinus pinaster)[J]. New Phytologist, 2014, 201(1): 230-241. doi: 10.1111/nph.12483 [28] Parchman T L, Gompert Z, Mudge J, et al. Genome-wide association genetics of an adaptive trait in lodgepole pine[J]. Molecular Ecology, 2012, 21(12): 2991-3005. doi: 10.1111/j.1365-294X.2012.05513.x [29] Kossuth S V, Biggs R H. Role of apophysis and outer scale tissue in pine cone opening[J]. Forest Science, 1981, 27(4): 828-836. [30] Cowling R M, Lamont B B. Variation in serotiny of three Banksia species along a climatic gradient[J]. Australian Journal of Ecology, 1985, 10(3): 345-350. doi: 10.1111/j.1442-9993.1985.tb00895.x [31] 毛建丰, 刘玉军, 刘 灏, 等. 高山松种实性状与生殖适应性[J]. 植物生态学报, 2007, 31(2):291-299. [32] Johnson E A, Gutsell S L. Heat budget and fire behaviour associated with the opening of serotinous cones in two Pinus species[J]. Journal of Vegetation Science, 2010, 4(6): 745-750. [33] Tapias R, Climent J, Pardos J A, et al. Life histories of Mediterranean pines[J]. Plant Ecology, 2004, 171(1-2): 53-68. [34] Hernández-Serrano A, Verdú M, González-Martínez S C, et al. Fire structures pine serotiny at different scales[J]. American Journal of Botany, 2013, 100(12): 2349-2356. doi: 10.3732/ajb.1300182 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 5036
- HTML全文浏览量: 3206
- PDF下载量: 57
- 被引次数: 0
