


-
紫耳箭竹(Fargesia decurvata J. L. Lu)是禾本科箭竹属的木本克隆植物,在西南地区山地阔叶林内广泛分布[1],有极强的适应能力和扩张能力,为金佛山国家自然保护区林下灌木层优势种[2],对该地区的水土保持、小气候环境的调节、林分更新和群落演替等均有影响。紫耳箭竹属于多年生一次开花结实植物,其同时大量开花和集中死亡是一种大型的群落内干扰,会对森林群落的结构和功能造成严重影响。自然界中成片而定期开花竹类的营养生长阶段可达30~120 a[3],大面积开花现象极为罕见,其中,紫耳箭竹大面积开花事件也未见记载。目前对竹子开花相关的研究较少,综观多年研究成果,主要集中在开花原因[4]、开花生物学特性[5]以及开花分子生物学[6]、开花对森林群落产生的后果[7]等方面,而成片开花死亡紫耳箭竹的有性繁殖和克隆生长对比研究未见报道。
植物体对有限资源的分配始终存在权衡,而克隆植物既能进行有性繁殖,也能进行无性繁殖,因此,这种权衡不仅体现在营养生长与繁殖之间,也体现在有性繁殖与无性繁殖之间[8-9]。植物资源分配的研究大多集中在生物量配置上[10],生物量分配格局不仅体现了各器官间的差异性分配,也反映了植物体在不同生活史阶段的分配特征和对不同环境条件的响应[11-12]。目前,对克隆植物的生物量配置和资源权衡的研究,主要集中在1年生或多年生草本植物[13-17],竹类植物方面则相对较少。对有性繁殖与克隆生长权衡的研究也未深入。本试验的研究地点位于重庆市金佛山国家自然保护区,2019年该地区的紫耳箭竹大面积开花结实。选取林下灌木层优势种为紫耳箭竹的3个典型林型(落叶阔叶林、常绿落叶阔叶混交林、常绿阔叶林),对其生活史中的营养生长期和果期的形态特征和生物量配置、繁殖特征进行研究,以探究不同林型中营养生长期和有性繁殖期紫耳箭竹种群形态可塑性和生物量分配的差异及探讨长寿命一次性开花结实的竹类克隆生长和有性繁殖权衡的特性。
-
研究地为重庆市金佛山国家级自然保护区(28°46′~29°38′ N,106°54′~107°27′ E)的烂坝菁,位于重庆市南川区,地处大娄山北坡,为亚热带湿润季风气候,冬短、春早、夏长,雨热同季、气候垂直变化明显。山体主要由灰岩和石灰岩组成,属于典型的喀斯特地貌。年均气温8.2℃,年降水量1 434 mm,海拔约1 400~2 252 m,高程差大,植被类型丰富,山体中上部主要森林类型为山地阔叶林(落叶阔叶林、常绿落叶阔叶混交林、常绿阔叶林),林下广泛分布紫耳箭竹[2],为灌木层优势种。
-
选择立地条件基本一致的3种典型群落类型:落叶阔叶林(De)、常绿落叶阔叶混交林(De+Ev)和常绿阔叶林(Ev)作为研究样地(样地基本概况见表1),每个林型样地面积约1 hm2,间隔不小于50 m,从2016年11月紫耳箭竹种群处于营养生长期开始对上述样地进行连续观察。2019年01月,紫耳箭竹种群出现开花前兆(紫耳箭竹种群有少量植株的枝条顶端第一叶展开处开始膨大,将之取下进行解剖发现有穗状花序组织),到2019年3月,花序逐渐明显,从枝条顶端开始抽穗。2019年6月中旬,紫耳箭竹种群中绝大部分分株进入果期。
表 1 样地基本环境概况
Table 1. Overview of the basic environment of the sample plot
林型
Habitat坡度
Slope/(°)坡向
Aspect坡位
Slope position纬度(N)
Latitude经度(E)
Longitude海拔
Altitude/mDe 18.09 西北 中坡 29°01′18″ 107°09′01″ 1 520 Ev+De 13.44 西北 中坡 29°01′18″ 107°09′05″ 1 529 Ev 18.13 西北 中坡 29°01′18″ 107°09′09″ 1 527 2016年11月,在紫耳箭竹营养生长期选择3个不同林型样地(De、Ev+De、Ev),在每个林型样地中随机选取20个1 m2的小样方,用锄头和铁锹等工具将小样方内的紫耳箭竹分株全部挖出,随机选择其中一个完整分株作为研究材料。2019年6月,以相同的方式在上述3个林型样地中随机挖取处于果期的20株紫耳箭竹完整分株,并用自封袋将小穗包好。2次取样过后均将分株编号后测定株高,将植株带回实验室进行形态指标测定:单株叶片数、单叶面积、隔离者(连接2个分株的地下根状茎)长度、隔离者直径、隔离者节间数、隔离者节间均长(隔离者长度/隔离者节间数)、单株小穗数、单株种子数(计数小穗数后,脱粒计种子数和结实率),并采用沉水法(体积置换法)测量植株根、秆、隔离者的体积。使用根系扫描仪(Win RHIZO Pro v. 2004c Root Analysis System, Canada)计算单叶面积。用游标卡尺和直尺等对各分株隔离者指标进行测量。将各分株地下根系、隔离者分离并编号(同一分株不同部位编号相同),于室外用流水冲洗干净并擦干,分别装入信封,同时将小穗(笋芽)、叶、秆、枝剪下,连同种子分别置于信封中并进行编号,放入烘箱于70℃条件烘干至恒质量取出,用 1 /10 000 天平分别称取分株各构件的生物量,并计算秆、根和隔离者的组织密度。
-
2017年5月,研究地区的紫耳箭竹进入竹笋萌发盛期,在3个不同林型样地(De、Ev+De、Ev)分别随机选择27个1 m × 1 m的小样方,记录小样方里面的紫耳箭竹竹笋萌发数;2020年8月,研究地的林下紫耳箭竹幼苗进入萌发盛期,在3个不同林型样地分别随机选取45个1 m × 1 m的小样方,记录小样方里面的紫耳箭竹幼苗数。
-
用分株各组分的生物量占分株总生物量的百分比作为生物量分配的数量指标,生殖分配指小穗生物量占总生物量的比值,结实率为单个分株种子数与总粒数的比值,组织密度为部分构件的干质量与该部分构件烘干前体积的比值[18]。用SPSS 20.0对营养生长期和果期紫耳箭竹各构件在3个林型的形态特征、生物量积累特征以及生物量分配特征、繁殖特征进行单因素方差(p < 0.05)分析,比较3个林型间的差异性,并作 LSD多重比较;对3个林型下的紫耳箭竹在营养生长期和果期的形态特征、生物量积累特征和生物量分配特征、繁殖特征进行独立样本T检验,采用Origin 2018作图。
-
不同时期、不同林型对紫耳箭竹分株的部分形态指标有显著影响(表2)。在营养生长期,紫耳箭竹分株在Ev的单株叶片数、株高、隔离者长度、隔离者节间数、隔离者直径、隔离者节间均长均显著低于De和De+Ev(p < 0.05),单叶面积也是Ev的最小。果期,De的紫耳箭竹分株小穗数显著高于De+Ev和Ev(p < 0.05),其余各指标在不同林型间无显著差异。
表 2 营养生长期和果期不同林型下紫耳箭竹分株的形态特征(平均值 ± 标准差)
Table 2. The morphological traits of F.decurvata ramets under different forest types and different stage(Means ± SD)
形态指标
Morphological traits落叶阔叶林
De常绿落叶阔叶混交林
De+Ev常绿阔叶林
Ev营养生长期
Vegetative stage叶片数Leaf numbers/(片·株−1) 32.55 ± 34.53 a 27.50 ± 18.47 a 10.05 ± 8.19 b 株高 Plant Height/cm 70.25 ± 20.56 a 85.00 ± 15.15 a 42.55 ± 12.33 b 隔离者长度 Spliter length/cm 25.37 ± 7.94 a 20.18 ± 9.50 a 8.60 ± 7.16 b 隔离者节间数 Internode number of spliter 33.20 ± 6.41 a 27.3 ± 7.87 a 18.95 ± 10.41 b 隔离者直径 Mean diameter of spliter/cm 0.33 ± 0.08 a 0.31 ± 0.06 a 0.22 ± 0.04 b 隔离者节间均长 Mean internode length of spliter/cm 0.75 ± 0.14 a 0.71 ± 0.21 a 0.40 ± 0.18 b 单叶面积 Leaf area/(cm × cm) 8.95 ± 2.54 a 9.76 ± 3.51 a 7.85 ± 3.57 a 小穗数Spikelet numbers/(个·株−1) — — — 种子数Seed numbers/(个·株−1) — — — 果期
Ripening stage叶片数Leaf numbers/(片·株−1) 4.35 ± 4.44 a** 4.40 ± 2.80 a** 10.60 ± 13.02 ans 株高Plant Height/cm 112.51 ± 19.61 a** 111.24 ± 31.26 a** 112.75 ± 26.66 a** 隔离者长度 Spliter length/cm 19.06 ± 6.94 a* 17.72 ± 4.33 ans 18.50 ± 8.56 a** 隔离者节间数 Internode number of spliter 27.10 ± 7.83 a* 24.95 ± 4.62 ans 25.70 ± 10.17 a* 隔离者直径 Mean diameter of spliter/cm 0.22 ± 0.04 a** 0.20 ± 0.03 a** 0.22 ± 0.04 ans 隔离者节间均长 Mean internode length of spliter/cm 0.69 ± 0.12 ans 0.71 ± 0.14 ans 0.71 ± 0.15 a** 单叶面积 Leaf area/(cm × cm) 8.91 ± 2.24 ans 9.69 ± 1.91 ans 9.65 ± 1.87 a* 小穗数Spikelet numbers/(个·株−1) 22.35 ± 12.80 a 9.05 ± 4.75 b 11.4 ± 7.36 b 种子数Seed numbers/(个·株−1) 19.75 ± 15.75 a 33.7 ± 21.56 a 24.00 ± 20.24 a 注:不同的小写字母表示紫耳箭竹形态特征在不同林型间的差异;*号表示形态特征在营养生长期和果期的差异(*: p < 0.05, **: p< 0.01)。
Notes: Different lowercase letters indicate the differences in the morphological characteristics of F.decurvata among different forest types; * indicates the difference between morphological characteristics in the vegetative stage and ripening stage(*: p < 0.05, **: p < 0.01).不同林型、不同生长阶段的紫耳箭竹种群分株的部分形态特征之间存在差异。在De条件下,营养生长期的紫耳箭竹单株叶片数、隔离者长度、隔离者节间数和隔离者直径显著大于果期,隔离者节间均长和单叶面积与果期无显著差异(p > 0.05);在De+Ev条件下,营养生长期紫耳箭竹的单株叶片数和隔离者直径极显著大于果期(p < 0.01);在Ev条件下,营养生长期紫耳箭竹的隔离者长度、隔离者节间数、隔离者节间均长和单叶面积显著或极显著低于果期,单株叶片数和隔离者直径与果期无显著差异(p > 0.05)。从营养生长期到果期,随着小穗伸长生长,3 个林型中紫耳箭竹株高均有极显著增加( p < 0.01)。
-
不同时期、不同林型对紫耳箭竹分株部分构件的生物量积累也有显著影响(图1)。在营养生长期,Ev中紫耳箭竹每分株笋芽生物量最低,De中最高,但在3个林型间无显著差异(p > 0.05);Ev中的隔离者生物量、枝生物量、叶生物量、秆生物量均显著低于其余2种林型(p < 0.05),根生物量也在Ev中最低,与De+Ev差异显著 (p < 0.05)。果期,紫耳箭竹分株的根生物量、隔离者生物量、枝生物量、种子生物量、小穗生物量在3个林型间差异不显著(p > 0.05),其中,De中紫耳箭竹的枝生物量最高,根生物量、隔离者生物量在Ev中最高,但Ev中紫耳箭竹的小穗生物量和种子生物量却最低;叶生物量也是Ev中最高,且与De中差异显著(p < 0.05);De+Ev中紫耳箭竹的秆生物量显著低于Ev和De中紫耳箭竹的秆生物量(p < 0.05)。
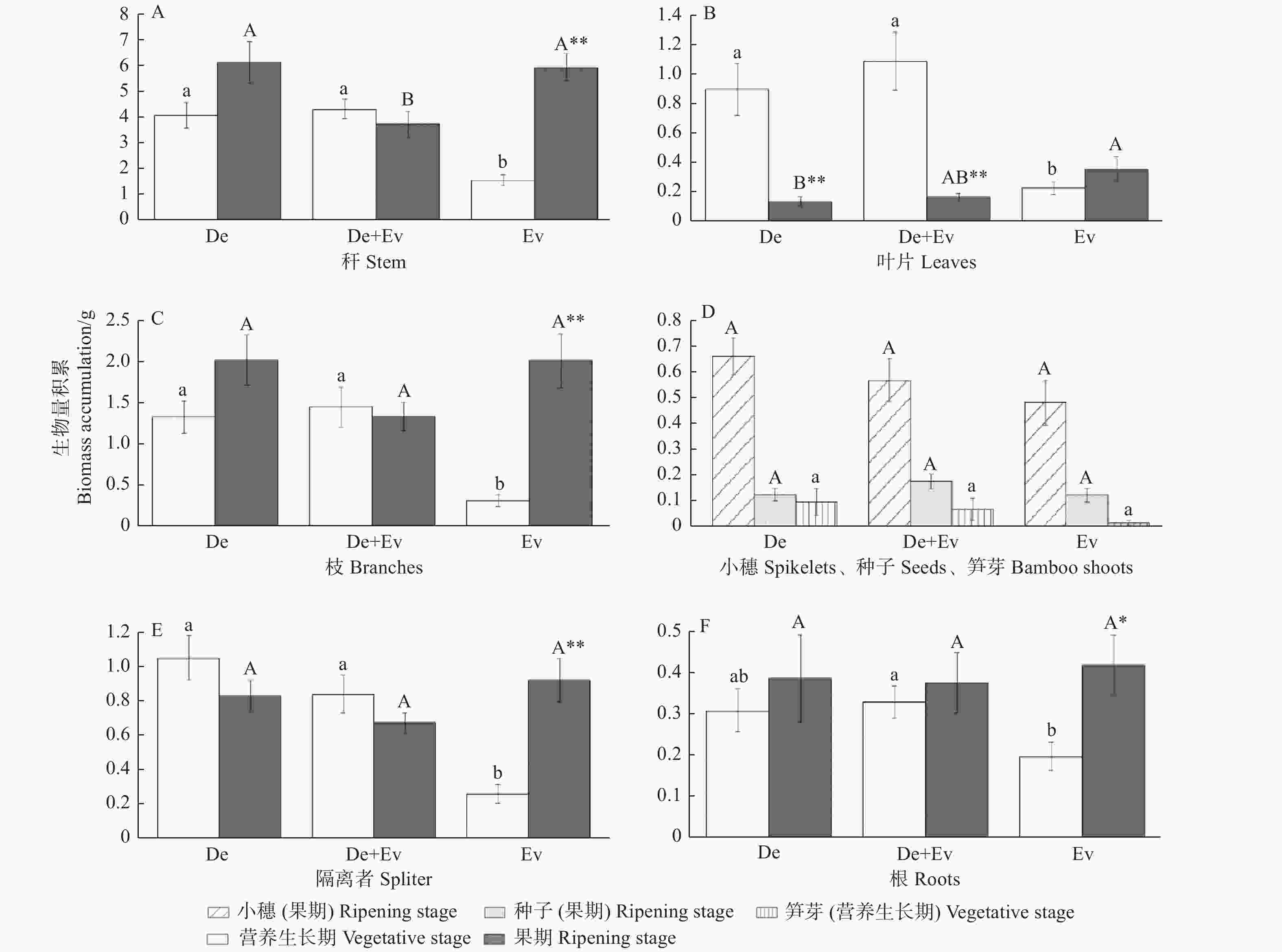
图 1 不同时期各林型下紫耳箭竹分株的各构件生物量积累特征
Figure 1. The biomass accumulation traits of F.decurvata ramets under different forest types and different stage
不同林型、不同生长阶段的紫耳箭竹种群分株的部分构件生物量积累量之间存在差异(图1);在De和De+Ev中,营养生长期仅叶片生物量显著大于果期,其余各构件生物量与果期无显著差异(p > 0.05);在Ev中,营养生长期的叶生物量与果期无显著差异,其余各构件生物量均显著低于果期(p < 0.05)。
-
不同生活史阶段紫耳箭竹分株的各构件生物量分配对不同林型响应方式不同(图2),3个林型下紫耳箭竹秆生物量占的比例均最大,超过50%;在营养生长期,Ev中紫耳箭竹分配给根的生物量显著高于De+Ev和De,而减少了对枝生物量的分配(p < 0.05);3个林型间的紫耳箭竹分株的秆生物量分配、隔离者生物量分配、叶生物量分配和芽生物量分配无显著差异(p > 0.05)。果期,Ev条件下更侧重于秆生物量的分配,显著高于De+Ev (p < 0.05);Ev的小穗生物量分配则有所减少,显著低于De (p < 0.05);隔离者生物量分配、根生物量分配、叶生物量分配、枝生物量分配在3个林型间无显著性差异(p > 0.05)。
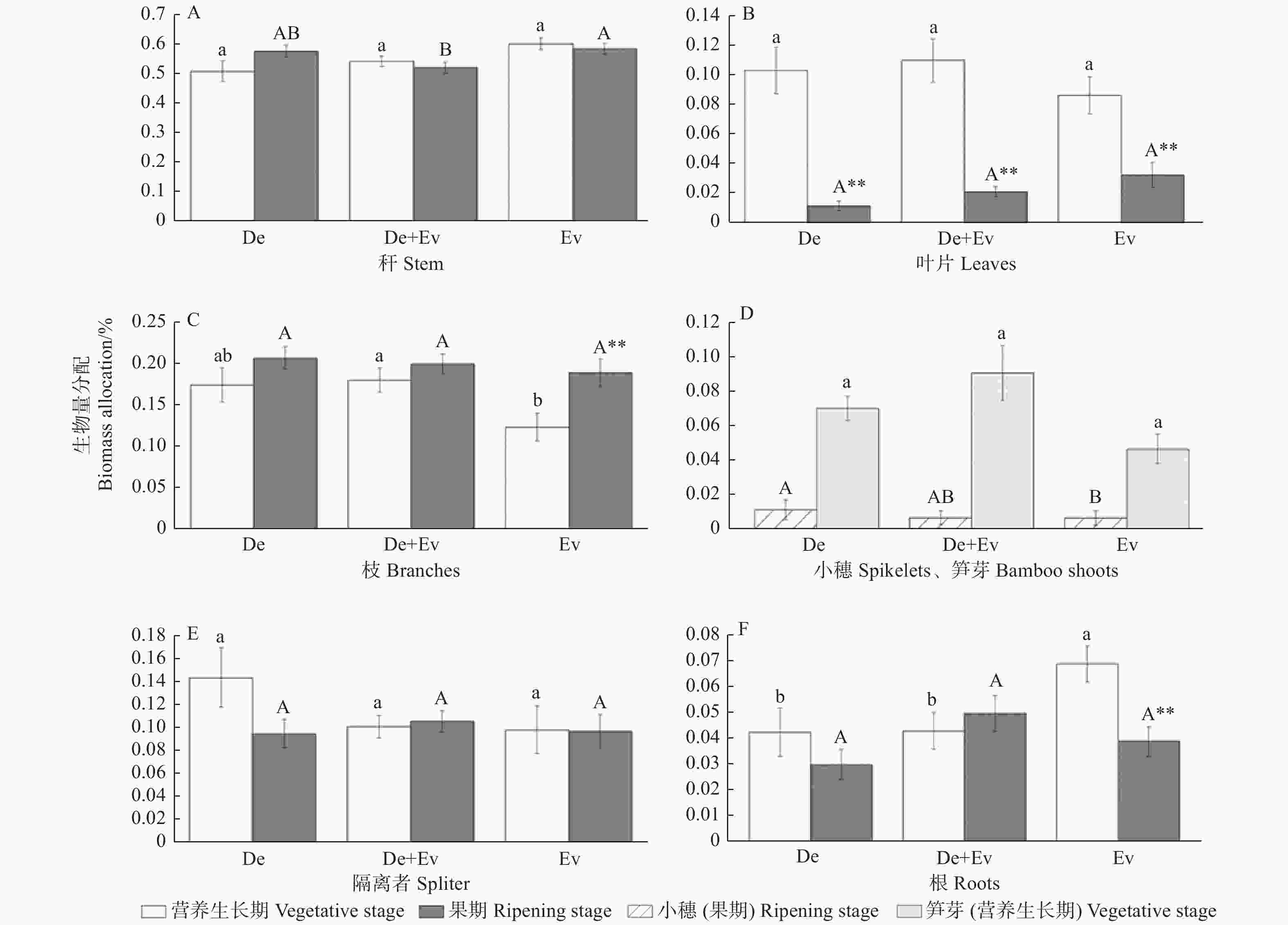
图 2 不同林型各时期紫耳箭竹分株各构件生物量分配特征
Figure 2. The biomass allocation traits of F.decurvata ramets under different forest types and different stage
不同林型下,构件生物量分配在不同生长发育阶段也不同(图2)。在De和De+Ev中,营养生长期的紫耳箭竹仅叶片生物量分配显著大于果期,其余各构件生物量分配与果期无显著差异(p > 0.05);在Ev条件下,营养生长期紫耳箭竹的秆生物量分配、隔离者生物量分配与果期无显著差异,根生物量分配和叶生物量分配显著高于果期,枝生物量分配则显著低于果期(p < 0.05)。
-
在营养生长期,紫耳箭竹种群分株在3个林型中均产生竹笋进行克隆繁殖,未见花序,混交林中的竹笋数显著高于常绿阔叶林( p < 0.05)。在果期结束后的竹苗萌发盛期对紫耳箭竹幼苗萌发情况进行调查,结果表明:紫耳箭竹种群在果期结束后均以种子发芽的方式进行有性繁殖,未见竹笋,其中,Ev的单位面积竹苗数显著低于其余2种林型(p < 0.05)(表3)。果期紫耳箭竹种群的结实率都很低,最高为13.4%(De),De+Ev和Ev中紫耳箭竹的结实率仅为5.5%和4.5%,显著低于De(p < 0.05),同时,每分株种子的生物量积累量也很低,为0.1~0.2 g(图1)。植物组织密度能够反映植物体的生长速率[18],对植物体的秆、隔离者和根组织密度的测量结果(表3)表明,3个林型的构件组织密度仅在处于果期的隔离者间存在显著差异(p < 0.05),营养生长期的秆组织密度极显著低于果期(p < 0.01)。
表 3 不同时期各林型下紫耳箭竹的繁殖特征(平均值 ± 标准差)
Table 3. The reproduction traits of F.decurvata ramets under different forest types and different stage(Means ± SD)
时期
Stage林型
Habitat竹苗密度/(株·m−2)
Bamboo seedlings竹笋数/(株·m−2)
Bamboo shoots结实率/%
Settingrate秆组织密度
Tissue density of stem /
(g·cm−3)隔离者组织密度
Tissue density of spliter /
(g·cm−3)根组织密度
Tissue density of roots /
(g·cm−3)营养生长期
Vegetative stageDe 6.98 ± 5.27 a 1.08 ± 1.14 ab — 0.68 ± 0.14 a** 0.53 ± 0.19 a 0.41 ± 0.08 a De+Ev 13.73 ± 12.00 a 2.79 ± 1.87 a — 0.64 ± 0.12 a** 0.48 ± 0.09 a 0.35 ± 0.09 a Ev 5.33 ± 6.08 b 0.10 ± 0.24 b — 0.55 ± 0.08 a** 0.52 ± 0.18 a 0.32 ± 0.11 a 果期
Ripening stageDe — 13.4 ± 14.4 a 0.75 ± 0.18 a 0.38 ± 0.19 b 0.24 ± 0.09 a De+Ev — 5.5 ± 4.6 b 0.81 ± 0.16 a 0.54 ± 0.27 ab 0.72 ± 0.82 a Ev — 4.5 ± 3.7 b 0.80 ± 0.16 a 0.73 ± 0.16 a 0.32 ± 0.14 a 注:不同的小写字母分别表示紫耳箭竹繁殖特征在不同林型间的差异;**表示繁殖特征在营养生长期和果期的差异(p < 0.01)。
Note: Different lowercase letters indicate the differences in the reproduction characteristics of F.decurvata among different forest types; ** indicates the difference between reproduction characteristics in the vegetative stage and ripening stage(p < 0.01). -
叶片是植物体进行光合作用获取养分资源的重要器官,大部分竹子开花后竹叶全部脱落[19]。随着De向Ev的过渡,在营养生长期,紫耳箭竹种群的叶片数呈逐渐降低的趋势,且Ev显著低于其余2个林型,但在果期,De和De+Ev的叶片迅速凋落,叶片数显著减少(表2),Ev中紫耳箭竹的叶片数则未发生明显变化,说明生活史阶段变化对紫耳箭竹的影响未体现在叶片数上。
根茎克隆植物是一种重要的克隆植物类型,其隔离者的高可塑性能有效提高克隆植物适应生境的能力[20],是衡量环境适应度的重要指标。紫耳箭竹的隔离者在不同生活史阶段和不同林型中有一定的差异,营养生长期,紫耳箭竹分株在冠层郁闭度相对较低的De和De+Ev下,其隔离者长度、隔离者节间数、隔离者直径、隔离者节间均长均显著高于Ev条件下的值(表2),这与华西箭竹(Fargesia nitida (Mitford) Keng f. ex Yi)和草本植物绢毛委陵菜(PotentillasericeaLinn.)等[21-22]的研究结果一致;但在对蛇莓(Duchesnea indica (Andr.) Focke)[23]的研究中发现,隔离者节间均长和隔离者长度在中等光照和资源水平下能达到最大值。果期,其变化趋势与营养生长期不同,3个林型中紫耳箭竹分株的隔离者长度、隔离者节间数、隔离者直径、隔离者节间均长均无显著性差异。从营养生长期到果期,De和De+Ev下紫耳箭竹分株的隔离者长度、隔离者节间数、隔离者直径、隔离者节间均长均呈降低的趋势,而在Ev中呈增加的趋势,并最终导致在3个林型下没有显著差异。紫耳箭竹作为多年生木本克隆植物,其隔离者对于不同林型条件的响应与竹类植物、一些1年生或多年生草本植物表现出了相同的趋势,但也与一些草本植物存在差异,说明不同物种对资源环境等的响应方式不同。由于Ev中光合有效辐射较低,紫耳箭竹在营养生长期所同化的资源无法满足自身生长需要,故隔离者投入也相对于在另外2种林型下小,果期紫耳箭竹的隔离者各指标在3个林型下没有显著差异。由于隔离者可以通过形态可塑性、觅食行为、生理整合作用等来适应环境[24],因此,进入有性繁殖期的紫耳箭竹分株,由于自身繁殖对资源等的需求加大,在营养生长期发育较差的隔离者可能会发生相应变化以达到最大适合度,最终导致其隔离者在3个林型间不具备显著差异。
果期,De条件下紫耳箭竹单株小穗数显著高于De+Ev和Ev条件下的单株小穗数目,结实率和每分株小穗生物量也最高,但每分株种子数目和种子生物量在3个林型间没有显著差异(表2、3,图2),说明在紫耳箭竹种群的有性繁殖过程中可能存在某种调节机制,即在环境条件和植株自身条件最佳时,种群分株对种子的资源投入量和种子产生量也是一定的,存在阈值限制[25]。
-
克隆性(Clonality)是区别克隆植物和非克隆植物的基本特性,克隆性包括克隆生长和克隆生殖,紫耳箭竹作为根源性木本克隆植物,既能通过克隆生长产生实际分株,也能通过有性繁殖产生种子进而发芽产生新植株,这就导致各构件在竞争有限资源时,植物体本身会进行权衡,包括有性繁殖和无性繁殖的权衡。秆和枝作为支撑器官在营养生长期和果期均占据很大的生物量分配(图1、2),从营养生长期到果期,紫耳箭竹增加小穗和种子生物量积累及分配的同时,De 中隔离者和叶片的生物量积累及分配在减少,De+Ev中秆和叶片的生物量积累及分配也在减少,Ev中则是根和叶片的生物量分配减少。说明植物体在利用有限资源的过程中存在权衡,增加繁殖构件资源分配的同时也在减少其他构件的资源分配,且在不同林型间这种权衡存在差异[8]。从营养生长向果期过渡过程中,除Ev外,其余2个林型下紫耳箭竹分株叶片生物量和生物量分配急剧下降,是由于资源间的权衡导致在繁殖过程中相对于其它构件,植株选择大量减少对叶片的资源投入,所损失的叶片光合作用产物可通过花穗的光合作用进行弥补[26]。而Ev中光合有效辐射低,红光/远红光的比例也较低,会影响植物体中的光敏色素分子活性、光合色素对光的吸收以及光信号的传导,在此条件下,紫耳箭竹为获取更多的光照会相对增加叶片的生物量和单位面积叶光合能力来适应弱光环境[27]。
不同林型中,随光资源的减少,紫耳箭竹分株在营养生长期各构件生物量均随之减少,生物量分配则表现为在营养生长期,枝的生物量分配从De向Ev减少,而根生物量分配增加,这与沙棘(Hippophae rhamnoides Linn.)的研究结果一致[28],其原因是在光照充足的情况下,与克隆生长相关的无性系构件获得的生物量比有性繁殖构件获得的生物量更少,枝作为同化器官可以间接影响有性繁殖[29]。
在营养生长期,3个林型中的紫耳箭竹均靠产生笋芽进行克隆生长(表3),这种方式成功率高并且有利于快速占据有限空间和资源[30]。果期,紫耳箭竹种群大量开花结实,加大了对小穗和种子的资源分配,取代了克隆生长,这种方式有利于种群间进行基因交流,产生新的基因型,以扩张新的环境[31],说明在某些情况下克隆生长和有性繁殖会根据环境资源的变化表现出相互替代的现象[8]。
-
无性繁殖和有性繁殖的权衡在不同植物以及生境间存在很大的差异,这种权衡会影响整个种群的适合度[8],紫耳箭竹作为多年生一次开花结实的克隆植物,在果期,将大量资源分配给繁殖器官导致分配给克隆生长的资源减少,使克隆生长几近消失(表3)。在一些草本克隆植物中,进行有性繁殖也会减少植物对克隆生长的资源分配[8-9, 13, 28],但在对荩草(Arthraxon hispidus (Thunb.) Makino)的研究中[14],这种权衡关系并不明显,因此,克隆生长和有性繁殖之间的权衡要综合物种和资源环境等差异进行考虑。
在金佛山自然保护区烂坝菁地区,紫耳箭竹主要分布在海拔1 300~1 500 m的林下,1 300 m以下的区域仅有少量分布,其开花时间稍晚于高海拔地区。研究地的紫耳箭竹种群在营养生长期产生竹笋进行克隆生长,而在果期克隆生长能力变弱甚至消失,种群靠开花结实产生的种子延续,并且在完成有性繁殖后,作为灌木层优势种的紫耳箭竹种群大面积干枯死亡,可以为种子萌发提供条件[7]。紫耳箭竹属于一年定型植物,具有极强的适应能力和扩张能力,在营养生长期通过克隆生长进行扩张,在短期内可以实现对适宜生境的有效利用,这也是紫耳箭竹在灌木层占据绝对优势地位的原因。但长年进行克隆繁殖会使种群中的基因型减少,一旦环境发生变化,种群将面临极大的风险,因此,种群存活的长期持久性在一定程度上还是依赖于有性繁殖对克隆多样性水平的补充[32],而成片开花的特性可以增加种群间基因交流的机会,使其能够产生更适应环境的基因型。紫耳箭竹的这种权衡特性使得其种群能够始终适应该地区的环境条件,但有性繁殖的风险较高,一是紫耳箭竹种群产生的种子数很少(表2),二是紫耳箭竹花后死亡的特性不仅为竹子的种子萌发提供了机会,也为其它物种的种子萌发创造了条件,加大了竹苗萌发和定居难度,因此,紫耳箭竹在该地区林下灌木层是否始终占优势并不确定。竹子的存在经常对树木的幼苗萌发、生存和生长产生不利影响,竹子开花枯死则会解除这种负面影响,为种子库中大量种子的萌发和生长提供机会,该地区的未来物种组成可能会由此发生巨大变化。
-
生活史阶段和林冠类型均会影响紫耳箭竹种群的形态、生物量积累与分配特征。从营养生长期到果期,落叶阔叶林和常绿落叶阔叶混交林中紫耳箭竹分株的叶片数和隔离者各指标均在降低,而常绿阔叶林中叶片数无明显变化,隔离者各指标呈现增高趋势;落叶阔叶林和常绿落叶阔叶混交林中的紫耳箭竹分株叶片生物量积累和分配急剧下降,其余构件的生物量积累和分配无明显变化,常绿阔叶林中各构件生物量除叶片外均显著增加,分配比例则在枝中显著增加,在叶片和根中显著降低。同时,紫耳箭竹克隆生长与有性繁殖间存在明显的权衡,随着有性繁殖的进行,克隆生长能力随之变弱并逐渐消失。本研究能为深入探讨竹类植物物质积累和资源分配策略以及竹类开花结实后林下灌木层物种更新和竹类自身的种群恢复提供参考。
金佛山不同群落紫耳箭竹生长和繁殖权衡的形态及生物量特征
Morphological and Biomass Characteristics of Fargesia decurvata in Different Forest Types in Jinfo Mountain
-
摘要:
目的 探究紫耳箭竹种群(Fargesia decurvata J. L. Lu)在不同林型下的生长和繁殖特征,探讨长寿命一次性开花结实的竹类克隆繁殖和有性繁殖的权衡特性。 方法 以金佛山国家级自然保护区3个典型林型:落叶阔叶林(De)、常绿落叶阔叶混交林(De+Ev)和常绿阔叶林(Ev)下的灌木层优势种紫耳箭竹为试验材料,研究不同林型下紫耳箭竹种群在营养生长期和果期的形态可塑性、生物量积累与分配特性以及繁殖特征。 结果 表明:(1)相较于营养生长期,果期紫耳箭竹单株叶片数在De和De+Ev中显著减少,在Ev中没有显著变化;隔离者长度、隔离者节间数、隔离者直径、隔离者节间均长在De和De+Ev中均降低,在Ev中均增加;果期De中的紫耳箭竹单株小穗数高于De+Ev和Ev。(2)从营养生长期向果期过渡过程中,De和De+Ev中的紫耳箭竹分株叶片生物量和分配比例急剧下降,其余构件的生物量和分配比例无明显变化;Ev中各构件生物量除叶片外均显著增加,分配比例则在枝中显著增加,在叶片和根中显著降低;秆和枝作为支撑器官在3个林型中均占据很大的生物量分配。(3)在营养生长期,3个林型中的紫耳箭竹均产生竹笋进行克隆生长,且De+Ev中的竹笋数显著高于Ev,Ev的单位面积竹苗数显著低于其余2种林型,秆组织密度显著低于果期。在果期均不产生竹笋只产生种子,且单株结实率和生殖分配都很低。 结论 紫耳箭竹种群的形态特征、生物量积累与分配特性在不同生活史阶段不同,且在不同林型间也存在差异;克隆生长与有性繁殖间存在明显的权衡,随着有性繁殖的进行,克隆生长能力随之变弱并逐渐消失。 Abstract:Objective To study the growth and reproduction characteristics of Fargesia decurvata J. L. Lu under different forest types, and the trade-off characteristics of clonal reproduction and sexual reproduction of long-lived, one-time flowering bamboos. Method F. decurvata, the dominant species in shrub layer under three forest types, i.e. deciduous broad-leaved forest (De), evergreen deciduous broad-leaved mixed forest (De+Ev) and evergreen broad-leaved forest (Ev) was selected as experimental materials, to study the morphological plasticity, biomass accumulation and biomass distribution characteristics and reproductive characteristics in vegetative stage and ripening stage. Result (1) From vegetative stage to ripening stage, the amount of leaves decreased significantly in De and De+Ev, and there was no significant change in Ev. The length of the splitter, the amount of internode of the splitter, the mean diameter of the splitter, and the internode length of the splitter all decreased in De and De+Ev, and increased in Ev. During the ripening stage, the amount of spikelets in De was higher than that in De+Ev and Ev. (2) During the transition from vegetative growth to ripening stage, the leaf biomass accumulation and leaf biomass allocation of F. decurvata ramets in De and De+Ev decreased sharply, there was no significant change in the biomass of other components in this two types of forest; the biomass accumulation of each component in Ev increased significantly except for the leaves, and the biomass allocation increased significantly in the branches, and decreased in the leaves and roots; the stems and branches as supporting organs accounted for a large amount of biomass allocation in the three forest types. (3) During the vegetative growth stage, F. decurvata in the three forest types produced shoots for clonal growth, and the amount of bamboo shoots per unit area of De+Ev was significantly higher than Ev. However, no shoots were produced during the ripening stage, only seeds were produced, and the amount of bamboo seedlings per unit area of Ev was significantly lower than that of the other two forest types. The tissue density was significantly higher than that in vegetative growth stage. The seed setting rate and reproductive distribution of F. decurvata were very low. Conclusion The morphological characteristics, biomass accumulation and biomass allocation characteristics of F. decurvata population are different in various life history stages, and there are also differences among these forest types. There is an obvious trade-off between clonal growth and sexual reproduction, as sexual reproduction progresses, the clonal growth capacity will weaken and disappear. -
图 1 不同时期各林型下紫耳箭竹分株的各构件生物量积累特征
Figure 1. The biomass accumulation traits of F.decurvata ramets under different forest types and different stage
图 2 不同林型各时期紫耳箭竹分株各构件生物量分配特征
Figure 2. The biomass allocation traits of F.decurvata ramets under different forest types and different stage
表 1 样地基本环境概况
Table 1. Overview of the basic environment of the sample plot
林型
Habitat坡度
Slope/(°)坡向
Aspect坡位
Slope position纬度(N)
Latitude经度(E)
Longitude海拔
Altitude/mDe 18.09 西北 中坡 29°01′18″ 107°09′01″ 1 520 Ev+De 13.44 西北 中坡 29°01′18″ 107°09′05″ 1 529 Ev 18.13 西北 中坡 29°01′18″ 107°09′09″ 1 527  下载: 导出CSV
下载: 导出CSV
表 2 营养生长期和果期不同林型下紫耳箭竹分株的形态特征(平均值 ± 标准差)
Table 2. The morphological traits of F.decurvata ramets under different forest types and different stage(Means ± SD)
形态指标
Morphological traits落叶阔叶林
De常绿落叶阔叶混交林
De+Ev常绿阔叶林
Ev营养生长期
Vegetative stage叶片数Leaf numbers/(片·株−1) 32.55 ± 34.53 a 27.50 ± 18.47 a 10.05 ± 8.19 b 株高 Plant Height/cm 70.25 ± 20.56 a 85.00 ± 15.15 a 42.55 ± 12.33 b 隔离者长度 Spliter length/cm 25.37 ± 7.94 a 20.18 ± 9.50 a 8.60 ± 7.16 b 隔离者节间数 Internode number of spliter 33.20 ± 6.41 a 27.3 ± 7.87 a 18.95 ± 10.41 b 隔离者直径 Mean diameter of spliter/cm 0.33 ± 0.08 a 0.31 ± 0.06 a 0.22 ± 0.04 b 隔离者节间均长 Mean internode length of spliter/cm 0.75 ± 0.14 a 0.71 ± 0.21 a 0.40 ± 0.18 b 单叶面积 Leaf area/(cm × cm) 8.95 ± 2.54 a 9.76 ± 3.51 a 7.85 ± 3.57 a 小穗数Spikelet numbers/(个·株−1) — — — 种子数Seed numbers/(个·株−1) — — — 果期
Ripening stage叶片数Leaf numbers/(片·株−1) 4.35 ± 4.44 a** 4.40 ± 2.80 a** 10.60 ± 13.02 ans 株高Plant Height/cm 112.51 ± 19.61 a** 111.24 ± 31.26 a** 112.75 ± 26.66 a** 隔离者长度 Spliter length/cm 19.06 ± 6.94 a* 17.72 ± 4.33 ans 18.50 ± 8.56 a** 隔离者节间数 Internode number of spliter 27.10 ± 7.83 a* 24.95 ± 4.62 ans 25.70 ± 10.17 a* 隔离者直径 Mean diameter of spliter/cm 0.22 ± 0.04 a** 0.20 ± 0.03 a** 0.22 ± 0.04 ans 隔离者节间均长 Mean internode length of spliter/cm 0.69 ± 0.12 ans 0.71 ± 0.14 ans 0.71 ± 0.15 a** 单叶面积 Leaf area/(cm × cm) 8.91 ± 2.24 ans 9.69 ± 1.91 ans 9.65 ± 1.87 a* 小穗数Spikelet numbers/(个·株−1) 22.35 ± 12.80 a 9.05 ± 4.75 b 11.4 ± 7.36 b 种子数Seed numbers/(个·株−1) 19.75 ± 15.75 a 33.7 ± 21.56 a 24.00 ± 20.24 a 注:不同的小写字母表示紫耳箭竹形态特征在不同林型间的差异;*号表示形态特征在营养生长期和果期的差异(*: p < 0.05, **: p< 0.01)。
Notes: Different lowercase letters indicate the differences in the morphological characteristics of F.decurvata among different forest types; * indicates the difference between morphological characteristics in the vegetative stage and ripening stage(*: p < 0.05, **: p < 0.01).
下载: 导出CSV
表 3 不同时期各林型下紫耳箭竹的繁殖特征(平均值 ± 标准差)
Table 3. The reproduction traits of F.decurvata ramets under different forest types and different stage(Means ± SD)
时期
Stage林型
Habitat竹苗密度/(株·m−2)
Bamboo seedlings竹笋数/(株·m−2)
Bamboo shoots结实率/%
Settingrate秆组织密度
Tissue density of stem /
(g·cm−3)隔离者组织密度
Tissue density of spliter /
(g·cm−3)根组织密度
Tissue density of roots /
(g·cm−3)营养生长期
Vegetative stageDe 6.98 ± 5.27 a 1.08 ± 1.14 ab — 0.68 ± 0.14 a** 0.53 ± 0.19 a 0.41 ± 0.08 a De+Ev 13.73 ± 12.00 a 2.79 ± 1.87 a — 0.64 ± 0.12 a** 0.48 ± 0.09 a 0.35 ± 0.09 a Ev 5.33 ± 6.08 b 0.10 ± 0.24 b — 0.55 ± 0.08 a** 0.52 ± 0.18 a 0.32 ± 0.11 a 果期
Ripening stageDe — 13.4 ± 14.4 a 0.75 ± 0.18 a 0.38 ± 0.19 b 0.24 ± 0.09 a De+Ev — 5.5 ± 4.6 b 0.81 ± 0.16 a 0.54 ± 0.27 ab 0.72 ± 0.82 a Ev — 4.5 ± 3.7 b 0.80 ± 0.16 a 0.73 ± 0.16 a 0.32 ± 0.14 a 注:不同的小写字母分别表示紫耳箭竹繁殖特征在不同林型间的差异;**表示繁殖特征在营养生长期和果期的差异(p < 0.01)。
Note: Different lowercase letters indicate the differences in the reproduction characteristics of F.decurvata among different forest types; ** indicates the difference between reproduction characteristics in the vegetative stage and ripening stage(p < 0.01).
下载: 导出CSV
-
[1] 黄慧敏, 董 蓉, 向运蓉, 等. 不同群落冠层环境下紫耳箭竹笋期生长发育研究[J]. 植物科学学报, 2018, 36 (5):696-704. doi: 10.11913/PSJ.2095-0837.2018.50696 [2] Qian F, Zhang T, Guo Q, et al. Dense understory dwarf bamboo alters the retention of canopy tree seeds[J]. Acta Oecologica, 2016, 73: 38-44. doi: 10.1016/j.actao.2016.02.004 [3] 林树燕, 毛高喜. 竹子开花习性和开花竹林的更新[J]. 林业科技, 2007, 32(5):23-25. doi: 10.3969/j.issn.1001-9499.2007.05.008 [4] 徐振国, 黄大勇, 郭起荣, 等. 麻竹开花前后主要营养元素变化研究[J]. 世界竹藤通讯, 2017, 15(1):1-5. [5] 乔士义, 廖光庐. 毛竹开花生物学特性的观察[J]. 竹类研究, 1984(2):20-25. [6] 田 波, 陈永燕, 严远鑫, 等. 一个竹类植物MADS盒基因的克隆及其在拟南芥中的表达[J]. 科学通报, 2005, 50(2):145-151. doi: 10.3321/j.issn:0023-074X.2005.02.008 [7] Marchesini V A, Sala O E, Austin A T. Ecological consequences of a massive flowering event of bamboo (Chusquea culeou) in a temperate forest of Patagonia, Argentina[J]. Journal of Vegetation ence, 2010, 20(3): 424-432. [8] Ronsheim M L, Bever J D. Genetic variation and evolutionary trade-offs for sexual and asexual reproductive modes in Allium vineale (Lillaceae)[J]. American Journal of Botany, 2000, 87: 1769-1777. doi: 10.2307/2656827 [9] Wu G, Hu T, Liu Z. Trade-off of sexual and asexual recruitment in a dominant weedLigularia virgaurea (Maxim.) in alpine grasslands (China)[J]. Journal of Ecology, 2010, 58(1): 81-86. [10] Ivo Müller, Bernhard S, Jacob W. The effect of nutrient availability on biomass allocation patterns in 27 species of herbaceous plants[J]. Perspectives in Plant Ecology, Evolution & Systematics, 2000, 3(2): 115-127. [11] Ge J, He J, Sun X, et al. Ecological responses of the invasive alien plant Solidago canadensis to changes of soil water content[J]. Acta Botanica Boreali-Occidentalia Sinica, 2010, 30(3): 575-585. [12] 洪 岚, 刘梦佼, 沈浩, 等. 两种生境中薇甘菊种群的生物量生殖分配[J]. 武汉植物学研究, 2010, 28(5):598-605. [13] 侯真珍, 何 爽, 王风雷, 等. 珍稀克隆植物北极花有性繁殖动态及影响因素[J]. 生态学杂志, 2013, 32(12):3167-3172. [14] 徐晓霞, 刘金平, 游明鸿, 等. 遮阴和干旱对荩草克隆生长和有性繁殖及权衡关系的影响[J]. 草业学报, 2019, 28(2):121-132. [15] 初丽爽, 李海燕, 杨允菲. 松嫩平原异质生境羊草种群营养繁殖特征[J]. 应用生态学报, 2020, 31(1):83-88. [16] Dorken M E, Eckert C G. Severely reduced sexual reproduction in northern populations of a clonal plant, Decodon verticillatus (Lythraceae)[J]. Jornal of Ecology, 2001, 89(3): 339-350. doi: 10.1046/j.1365-2745.2001.00558.x [17] Mizuki I, Ishida K, Kikuzawa K. Sexual and vegetative reproduction in the aboveground part of a dioecious clonal plant, Dioscorea japonica(Dioscoreaceae)[J]. Ecological research, 2005, 20(4): 387-393. doi: 10.1007/s11284-005-0036-y [18] Pérez-Harguindeguy N, Diaz S, Gamier E, et al. New handbook for standardised measurement of plant functional traits worldwide[J]. Australian Journal of Botany, 2013, 61: 167-234. doi: 10.1071/BT12225 [19] 柴振林, 秦玉川, 华锡奇, 等. 竹子开花原因研究进展[J]. 浙江林业科技, 2006, 26(2):53-57. doi: 10.3969/j.issn.1001-3776.2006.02.015 [20] Evans J P. The effect of local resource availability and clonal integration on ramet functional morphology in hydrocotyle bonariensis[J]. Oecologia, 1992, 89: 265-276. doi: 10.1007/BF00317227 [21] 陶建平, 宋利霞. 亚高山暗针叶林不同林冠环境下华西箭竹的克隆可塑性[J]. 生态学报, 2006, 26(12):4019-4026. doi: 10.3321/j.issn:1000-0933.2006.12.013 [22] 张淑敏, 陈玉福, 董 鸣. 匍匐茎草本绢毛匍匐委陵菜对局部遮荫的克隆可塑性[J]. 植物学报(英文版), 2000, 42 (1):89-94. [23] 董 鸣, 张淑敏, 陈玉福. 匍匐茎草本蛇莓对基质养分条件的克隆可塑性[J]. 植物学报(英文版), 2000, 42(5):518-522. [24] Dong M. Morphological plasticity of the clonal herb Lamiastrum galeobdolon (L.) Ehrend & Polatschek in response to partial shading[J]. New Phytologist, 2010, 124(2): 291-300. [25] Cheplick, Gregory P. Life history trade-offs in Amphibromus scabrivalvis (Poaceae): Allocation to clonal growth, storage, and cleistogamous reproduction[J]. American Journal of Botany, 1995, 82(5): 621-629. doi: 10.1002/j.1537-2197.1995.tb11506.x [26] Mcdowell S C L, Mcdowell N G, Hultine M K. Carbon and nitrogen allocation to male and female reproduction in Rocky Mountain Douglas-fir (Pseudotsuga menziesii var. glauca, Pinaceae)[J]. American Journal of Botany, 2000, 87(4): 539-546. doi: 10.2307/2656598 [27] 高荣孚, 张鸿明. 植物光调控的研究进展[J]. 北京林业大学学报, 2002, 24(5):235-243. doi: 10.3321/j.issn:1000-1522.2002.05.046 [28] 姜 准, 刘丹一, 陈贝贝, 等. 中国沙棘克隆生长对造林密度的早期响应及其生物量分配调节机制[J]. 林业科学, 2017, 53(10):29-39. doi: 10.11707/j.1001-7488.20171004 [29] 翟雅芯, 王 玥, 魏仙仙, 等. 珍稀克隆植物北极花有性繁殖及其对资源的分配和利用[J]. 新疆农业科学, 2016, 53(6):1122-1128. [30] Honnay O, Bossuyt B. Prolonged clonal growth: escape route or route to extinction?[J]. Oikos, 2005, 108(2): 427-432. doi: 10.1111/j.0030-1299.2005.13569.x [31] Jacques T, Jean N R, Philippe C. Reproductive versus vegetative recruitment of the invasive tree Schinus terebenthifolius: Implications for restoration on Reunion Island[J]. Restoration Ecology, 2007, 15(3): 412-419. doi: 10.1111/j.1526-100X.2007.00237.x [32] Santiago A, Ezcurra E. Propagation mechanisms inAgave macroacantha (Agavaceae), a tropical arid-land succulent rosette[J]. American Journal of Botany, 2002, 89(4): 632-641. doi: 10.3732/ajb.89.4.632 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 4825
- HTML全文浏览量: 2892
- PDF下载量: 56
- 被引次数: 0
