





-
杨树是我国重要的经济和能源物种,在工业、农业、林业等方面发挥着重要作用,具有广泛的应用价值和较高经济价值。随着杨树种植面积的进一步扩大,受环境、气候等因素影响导致病害频发,造成了巨大的经济损失[1]。目前,防治杨树病害主要靠物理防治或化学防治,不仅耗费人力、物力和财力,长期使用农药也造成了严重的环境污染。因此,培育杨树抗病品种是防治杨树病害最为经济、安全、有效的措施。与传统育种技术相比,分子育种技术能够定向、高效地培育出新品种,在植物抗病育种领域,尤其是周期较长的林木抗病育种领域具有较好的应用前景。研究杨树抗病的分子机理,可为分子育种提供理论支撑和基因资源,是开展杨树抗病分子育种的基础。
植物在长期应对生物胁迫过程中形成了PTI(PAMP-triggered immunity)和ETI(Effector triggered immunity)双层级免疫系统以防御病原菌的入侵。植物通过自身细胞表面模式识别受体(Pattern recognition receptors,PRRs)感知病原菌特征分子(PAMPs,pathogen-associated molecular patterns)可激活PTI反应,通过体内核苷酸结合和富亮氨酸重复受体(Nucleotide-binding domain and leucine-rich repeat containing receptor,NLRs)识别病原菌释放的毒性效应子(Effectors)可启动ETI反应,PTI和ETI协同作用可激活一系列抗病反应的发生,包括病程相关基因的表达、活性氧(ROS)爆发、细胞壁胼胝质沉积等[2-4]。对杨树-胶孢炭疽菌(Colletotrichun gloeosporioides)互作的研究表明,胶孢炭疽菌的侵染可诱导NB-LRR类蛋白的上调表达,继而引起细胞中活性氧爆发和超敏反应(HR)发生,以防御该病菌的进一步入侵[5]。在落叶松杨栅锈菌(Melampsora larici-populina Kleb)和杨树非亲和互作中,锈菌吸器母细胞可诱导寄主细胞壁增厚,并产生乳状突,从而阻止其侵入[6]。水杨酸、乙烯、茉莉酸等激素是植物防御反应重要的信号分子。外施水杨酸或其类似物可诱导杨树对锈菌(M. larici-populina)的防御反应[7]。WRKY、TGA等转录因子通过调控激素合成酶或激素受体的积累,抑制或激活激素信号转导通路中重要组分的表达,在植物防御反应过程中起关键调控作用[8-9]。对胡杨PeTGA1的研究表明,该转录因子不仅可以结合水杨酸合成酶PeSARD1的启动子从而诱导SA的积累,还可以直接调控病程相关基因PR1的表达。过表达PeTAG1转基因植株显著提高了胡杨对胶孢炭疽菌的抗病能力[10]。杨树PtoWRKY60可调控PR5.1、PR5.2、PR5.4、PR5.5和CPR5等基因的表达,在杨树中过表达这些基因显著增强了对溃疡病菌(Dothiorella gregaria Sacc.)的抗性[11]。
腐皮镰刀菌(Fusarium solani (Mart.) Sacc)属于半知菌亚门(Deuteromycotina),瘤座孢目(Tuberculariales),镰刀菌属(Fusarium)[12]。腐皮镰刀菌寄主范围十分广泛,可入侵杨树(Populus trichocarpa Torr. & Gray)[13]、桉树(Eucalyptus camaldulensis Dehn.)[14]、橄榄(Olea europaea L.)[15]、苏铁(Cycas L.)[16]等多种林木植物。该病菌主要从根部侵入,由维管束疏导组织向上扩展,造成根部腐烂、树干或枝条干枯、叶片枯萎脱落,最终导致植株死亡,具有传染速度快、致死率高等特点。由于杨树的广泛种植,由腐皮镰刀菌引起的病害造成了该树种的大面积死亡,经济损失惨重。但目前,该病害防治手段主要依赖于日常栽培管护,例如造林时尽量选择木质化程度较高的粗壮苗木以减少病害的发生[17]。有关该病害的研究主要集中于病害发生的描述和防治措施的探讨,针对杨树响应腐皮镰刀菌侵染的分子机理的研究仍十分有限。
随着测序手段和生物信息学技术的发展,高通量、快速、低成本的转录组(RNA-seq)技术已广泛应用于植物与病原菌分子互作研究,成为研究林木抗病机制、挖掘抗病基因的重要手段。本研究以银腺杨84K为试验材料,在转录水平上分析腐皮镰刀菌(F. solani (Mart.) Sacc)侵染杨树根部后不同时间内差异表达基因(Differentially Expression Genes)及相关信号通路,以期为进一步揭示杨树抗腐皮镰刀菌侵染的分子机理奠定基础。
-
供试杨树材料为中国林业科学研究院保存的84K杨(Populus alba × P. glandulosa)。将培养4周的组培苗移栽至上口径6.5 cm,下口径5 cm,高6 cm的小营养钵中,栽种基质为草炭土与珍珠岩(比例为3∶1),每盆装土量约为营养钵的90%,毎盆栽种1株。待幼苗在小营养钵中生长2周后移栽至大营养钵(内口径 × 底径 × 盆高 = 20 cm × 14.5 cm × 15 cm),生长一个月后进行接菌实验。培养条件为:温度22~25 ℃、相对湿度70%、光照16 h/黑暗8 h,生长期间适时浇水,保证植株正常生长。供试病原菌为腐皮镰刀菌(F. solani (Mart.) Sacc),由中国林业微生物菌种保藏管理中心提供,菌种保存号为CFCC 51703,培养条件为:温度25~28 ℃、黑暗,采用PDA培养基扩繁。
实验试剂及仪器:植物组织RNA提取试剂盒(天根,北京);反转录试剂盒 PrimeScript™ RT reagent Kit (Perfect Real Time)(Takara,日本);荧光定量试剂KAPA SYBR FAST qPCR Master Mix (KAPA Biosystems,美国);改良型霍格兰营养液NSP1020(酷莱博,北京);Calcofluor White stain solution(酷莱博,北京)。NanoDrop One 微量紫外可见分光光度计(Thermo Fisher Scientific,美国)、The Roche Light Cycler 480(Roche,瑞士);奥林巴斯荧光显微镜(Olympus BX 51)。
-
孢子液制备:将在PDA培养基上生长5 d的腐皮镰刀菌用灭菌水冲洗并收集其孢子制成孢子液。现用现制,以保证孢子的最佳活性。取100 μL 收集到的孢子液用血球计数板在显微镜下计数,调整孢子液浓度至1 × 107 个·mL−1。
-
针对茎干危害症状观察,选择正常生长2个月的84K植株,使用排针在茎干上制造伤口,利用直径5 mm的打孔器获取腐皮镰刀菌菌饼,敷于伤口处。将脱脂棉球用无菌水湿润后包裹菌饼,并用封口膜缠绕,爪夹固定。接种两天后,拆卸夹子、封口膜、棉球,以观察伤口处病斑扩散情况。针对根部危害症状的观察,用1 × 107 个·mL−1浓度的孢子液浸泡生长20 d的组培小苗根部3 h后移栽至土中培养,12 d后观察腐皮镰刀菌对植株生长的影响。
-
在茎干接腐皮镰刀菌菌饼5 d后,撕下伤口处树皮,用清水清洗,使用2~3滴Calcofluor white(CFW)荧光染料和2~3滴10% KOH对树皮进行染色,1 min后用清水清洗1~2次,将样本置于载玻片上于奥林巴斯荧光显微镜下观察,激发波长约在355 nm。
-
将在营养土中生长45 d的84K杨幼苗根部洗净,清洗过程中尽可能保证根部的完整。试验设置对照组(0 h)和2个接菌组(48 h和72 h),每组3棵幼苗,即3次生物学重复。利用添加了改良型霍格兰营养液的水调整孢子液浓度为1 × 107 个·mL−1,以保证植株正常生长所需的营养成分。霍格兰营养液主要营养成分浓度:Ca(NO3)2·4H2O为945 mg·L−1,KNO3为506 mg·L−1,NH4NO3为80 mg·L−1,KH2PO4为136 mg·L−1,MgSO4·7H2O为241 mg·L−1,FeNaEDTA为36.7 mg·L−1。将清洗干净的84K杨幼苗的根部置于提前配好的孢子液中,在浸泡孢子液的过程中,根部完全浸泡,并且保证苗子正常生长所需的适宜温度和光照强度。之后分别在侵染后0、48、72 h采集根部组织,并立即将其根组织置于液氮中冷冻,然后移至-80 ℃超低温冰箱中保存,以待后续实验使用。
-
样品由基迪奥生物科技(广州)有限公司进行高通量转录组测序。通过Trizol总RNA提取试剂盒对样品进行RNA的提取,采用安捷伦2100生物分析仪和琼脂糖凝胶电泳检验所提取RNA的质量。样品质量检测合格后,采用Illumina Novaseq6000进行双端测序分析。
-
通过fastq 0.18.0软件对序列进行质量控制,过滤去除只含adapter、含N比例大于10%、全部都是A碱基和低质量的 reads (质量值 Q≤20 的碱基数占整条 reads 的 50%以上)后得到Clean Reads。通过碱基错误率为1%(Q20)和1‰(Q30)来评估过滤后reads的质量,通常Q20在95%以上,Q30达到90%以上,说明测序结果质量合格。
-
采用HISAT 2.2.4软件将获得的Clean Reads与毛果杨4.1参考基因组(
https://phytozome-next.jgi.doe.gov/info/Ptrichocarpa_v4_1 )进行比对。根据比对结果,利用Stringtie重构转录本,并使用RSEM软件计算每个样本中所有基因的表达量,即FPKM。通过计算FPKM值比较不同样本中基因表达水平的差异。 -
利用DESeq2 软件对原始reads count进行标准化,根据模型计算假设检验概率(p value)和多重假设检验校正,得到FDR值(错误发现率)。以FDR<0.05、|log2FC|≥2为标准筛选显著差异表达基因。利用GO数据库(
http://www.geneontology.org/ )对差异基因进行功能注释分析,利用KEGG数据库(https://www.genome.jp/kegg/ )进行富集通路分析,使用基迪奥生物信息云平台(https://www.omicshare.com/ )对GO功能注释和KEGG富集分析并进行图形可视化。 -
选取6个差异表达基因进行荧光定量PCR分析,以验证转录组测序分析结果的准确性和可靠性。利用SnapGene 6.0.2和DNAMAN V6对所选基因和内参基因PagActin设计特异性引物,引物序列见表1。引物由生工生物工程(上海)股份有限公司合成。荧光定量PCR反应程序为:95 ℃ 5 min;95 ℃ 10 s,60 ℃ 10 s,72 ℃ 10 s,45个循环。采用2−△△CT法计算基因相对表达水平。每个处理设置3次生物学重复,每个生物学重复设置 4 次技术重复。采用GraphPad Prism 8.0.1 和Excel 2010对实验数据进行显著性分析并作图。
表 1 qRT-PCR 引物序列信息
Table 1. Primer sequences used for qRT-PCR analysis
序号 No. 基因号 Gene ID 引物序列 Primer sequence 1 Potri.005G187300 F:GTAGAGACCCTTTCGTTGCC R:CTTTTCCGGGACCAATCTCTAG 2 Potri.012G031400 F:CATGATCACCACCATCGCAA R:CGCGAGAAAGAGATCATGGC 3 Potri.015G101500 F:GTCTGGTCTGTCTTGCTCCT R:TGGTGATGAGAAGGATGGCA 4 Potri.003G166800 F:TCTTCCGAACATCTTGGGCT R:ACGTCAGGTTTTGCATCGAC 5 Potri.013G013800 F:GAGTGCCGAATGTTGTTGGT R:TCAGATTGCCAGCTTCCTCA 6 Potri.012G001400 F:CTGTCGCAGCATCAGGAAAT R:CTGCAGCGCTATCTTGTGAG 7 PagActin F:ACCCTCCAATCCAGACACTG R:TTGCTGACCGTATGAGCAAG -
腐皮镰刀菌(F. solani (Mart.) Sacc)在PDA培养基上生长时菌落较致密,正面形态呈雪白色,边缘呈黄色(图1A)。将该病菌菌饼接种于84K茎段,接种处利用CFW荧光染料染色后在显微镜下观察,发现病原菌孢子定殖在树皮上(图1B)。用孢子液侵染小苗根部,观察发现病菌从根部入侵,沿维管束向上蔓延,茎干变黑溃烂,叶片变黄枯萎(图1C),最终导致整个植株死亡。用腐皮镰刀菌菌饼接种84K茎段,2 d后开始出现病斑,12 d时茎段黑斑扩散明显(图1D)。

图 1 腐皮镰刀菌(F. solani (Mart.) Sacc)侵染84K杨幼苗情况
Figure 1. Colonies of F. solani (Mart.) Sacc and the inoculated 84K seedlings
-
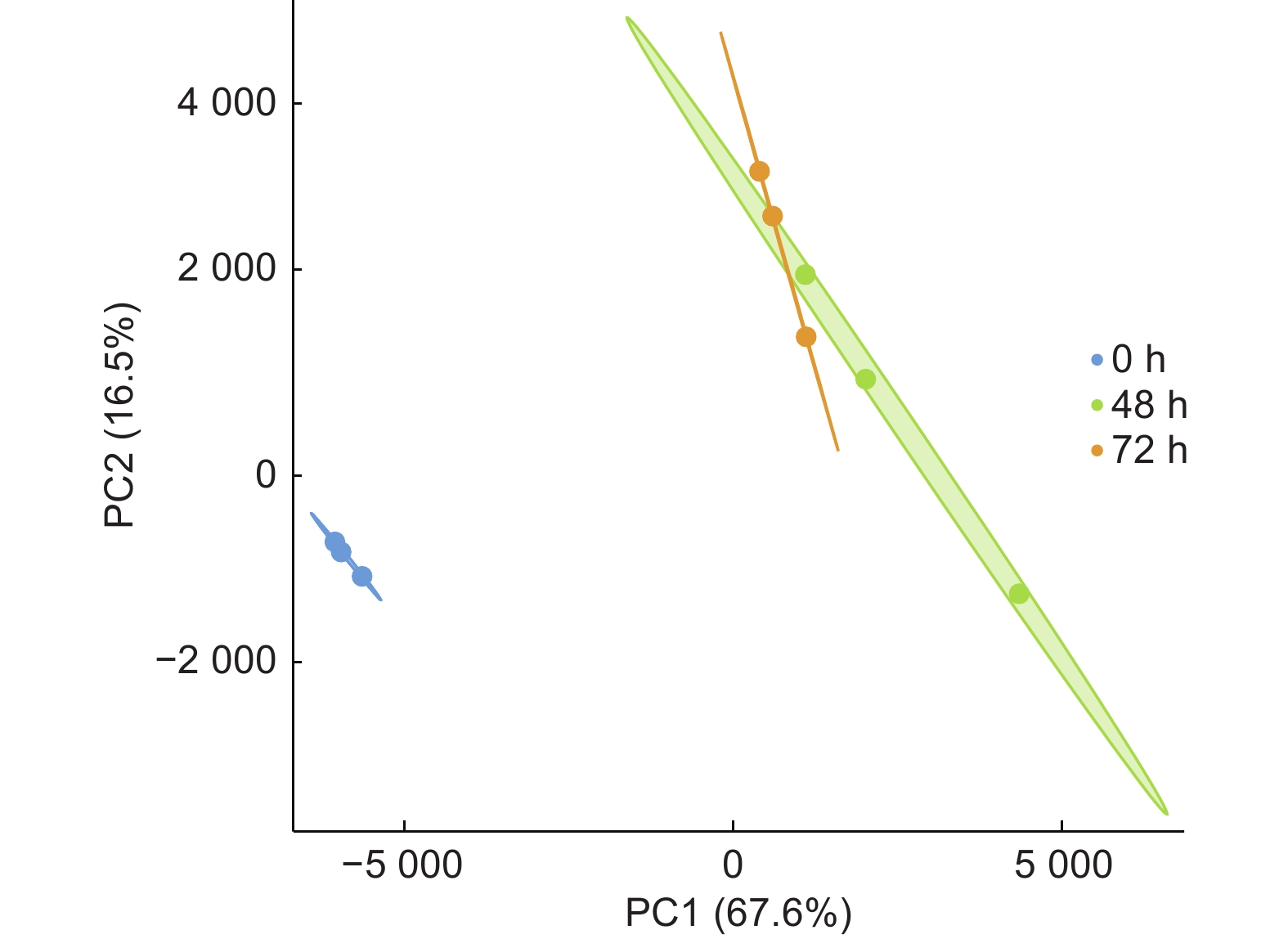
对侵染不同时期的84K杨根部组织进行RNA提取,通过质量检测后,经转录组测序获得原始数据。过滤低质量数据得到了37 366 426~469 727 56个Clean reads。对不同样本的质量检测表明,各样本Q20均高于97.61%,Q30均高于93.05%,GC含量为44.05%~45.79%(表2)。样品主成分分析(图2)显示,接菌组(48 h和72 h)与对照组(0 h)各聚为一类,样品间生物学重复较好。
表 2 转录组测序数据质控分析
Table 2. Quality test of transcriptome sequencing data
样品
Sample原始数据量
Raw reads/bp高质量数据量
Clean reads/bp准确率
Clean ratio/%Q20
Content of Q20/%Q30
Content of Q30/%GC含量
Content of GC/%0 h-1 40 228 010 40 032 220 99.51 97.89 93.63 44.19 0 h-2 41 478 100 41 262 044 99.48 97.76 93.37 44.14 0 h-3 42 237 814 42 014 800 99.47 97.76 93.39 44.05 48 h-1 40 187 478 40 029 556 99.61 97.94 93.75 45.24 48 h-2 37 900 752 37 782 074 99.69 98.11 94.18 45.38 48 h-3 41 773 292 41 533 374 99.43 97.61 93.05 45.43 72 h-1 37 472 500 37 366 426 99.72 98.08 94.19 45.71 72 h-2 38 707 252 38 547 366 99.59 97.74 93.41 45.79 72 h-3 47 140 310 46 972 756 99.64 97.86 93.67 45.47 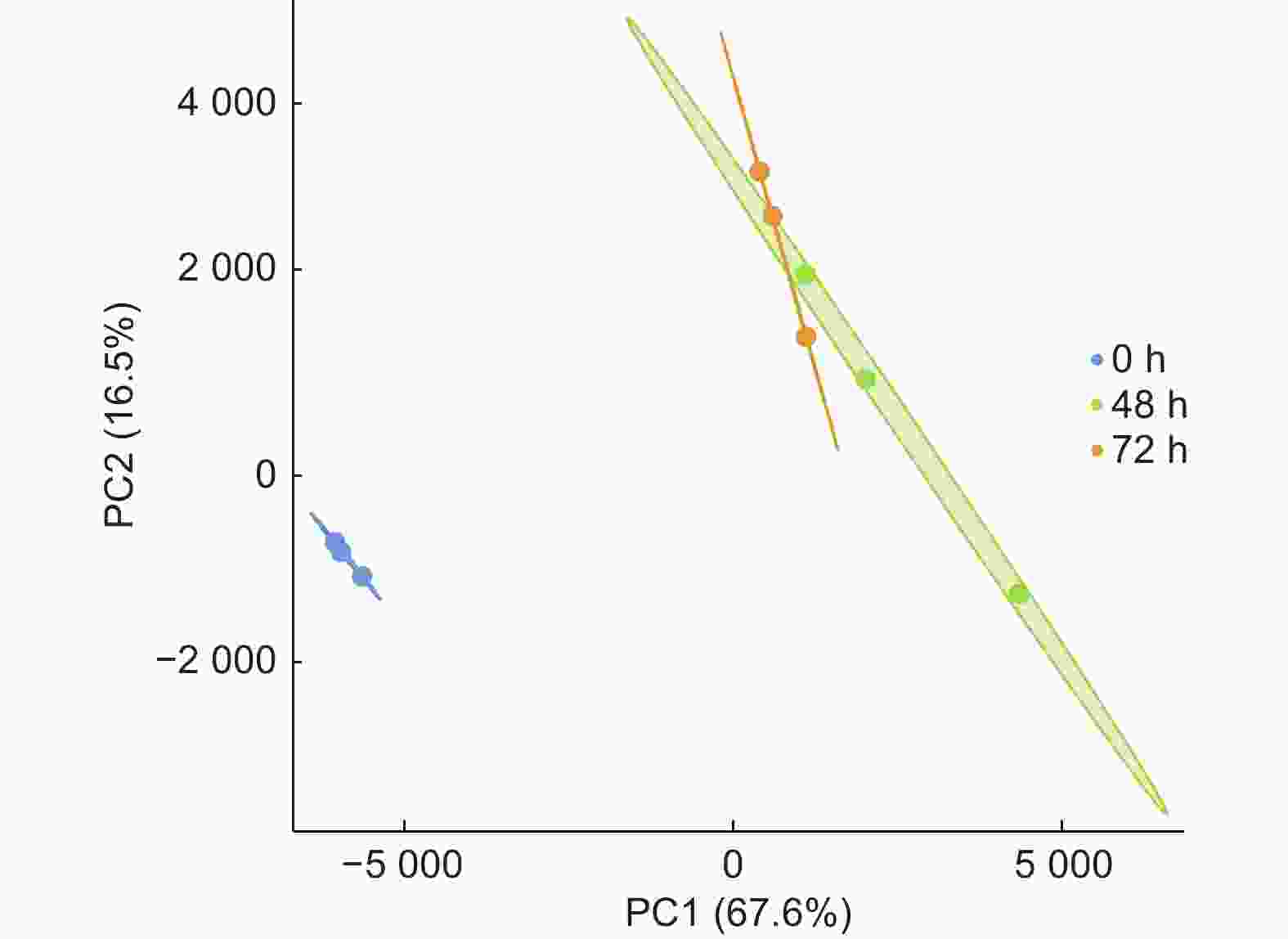
图 2 样品主成分分析
Figure 2. PCA analysis of the samples
-
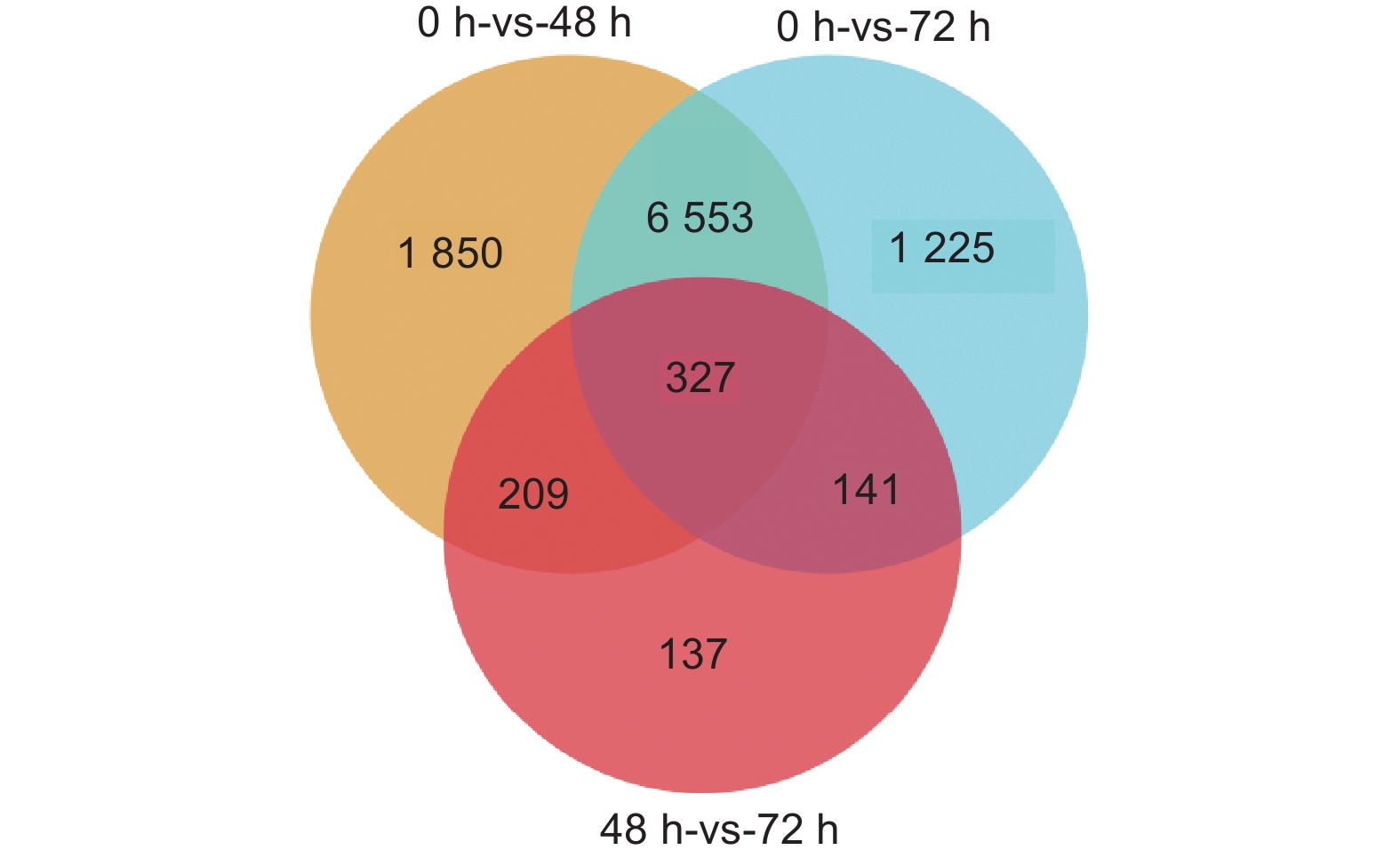
通过转录组测序共检测到35 235个表达基因,以FDR<0.05、差异倍数FC≥2或者FC≤0.5为筛选条件,对DEGs数量进行统计学分析。结果显示,接菌后48 h,对照组和接菌组之间共检测到8 939个差异表达基因,其中上调基因3 020个,下调基因5 919个(图3A)。接菌后72 h,共检测到差异表达基因8 246个,其中上调基因2 910个,下调基因5 336个(图3B)。进一步统计发现,接菌48 h和72 h后均被检测到的差异表达基因共有327个(图4)。
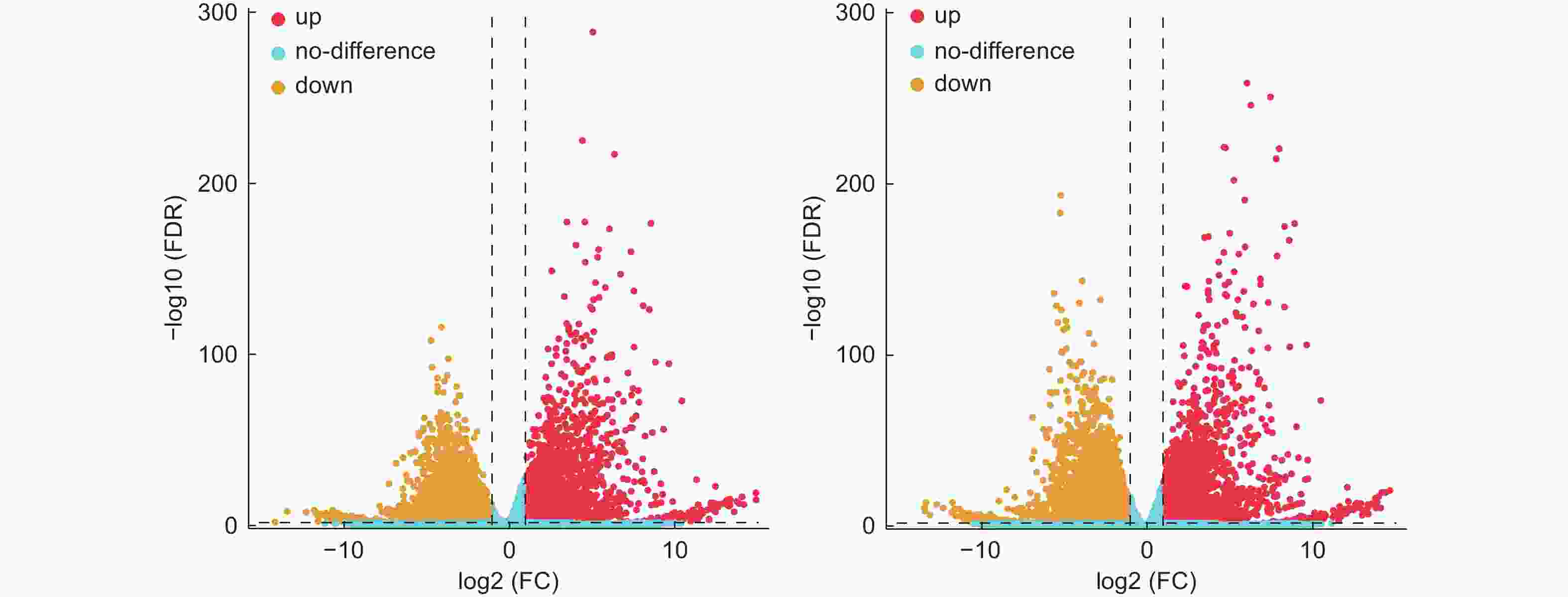
图 3 杨树接种腐皮镰刀菌后差异表达基因火山图
Figure 3. Volcano map of DEGs in poplar root inoculated with F. solani (Mart.) Sacc
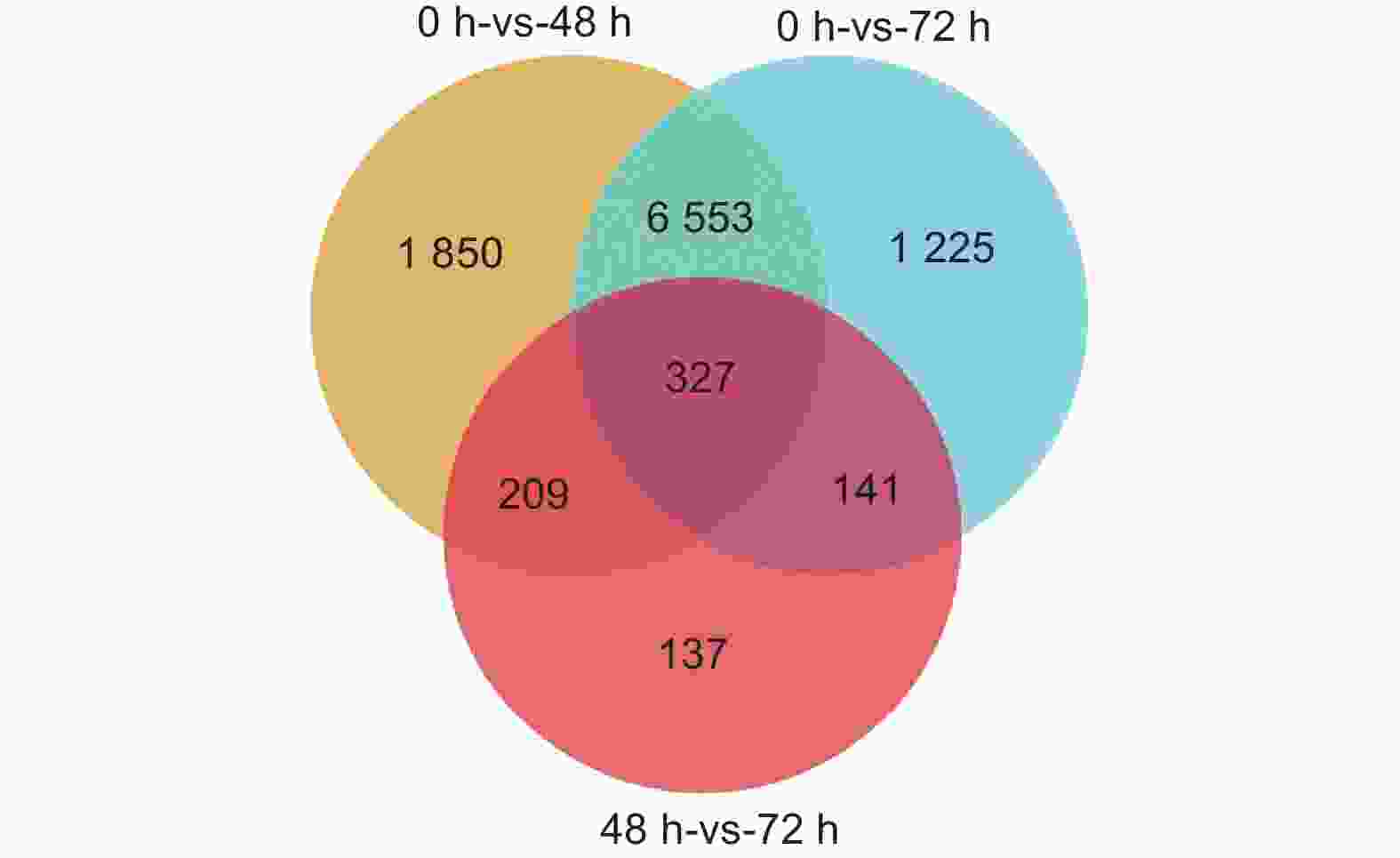
图 4 差异表达基因的维恩图
Figure 4. Venn diagram of differentially expressed genes
-
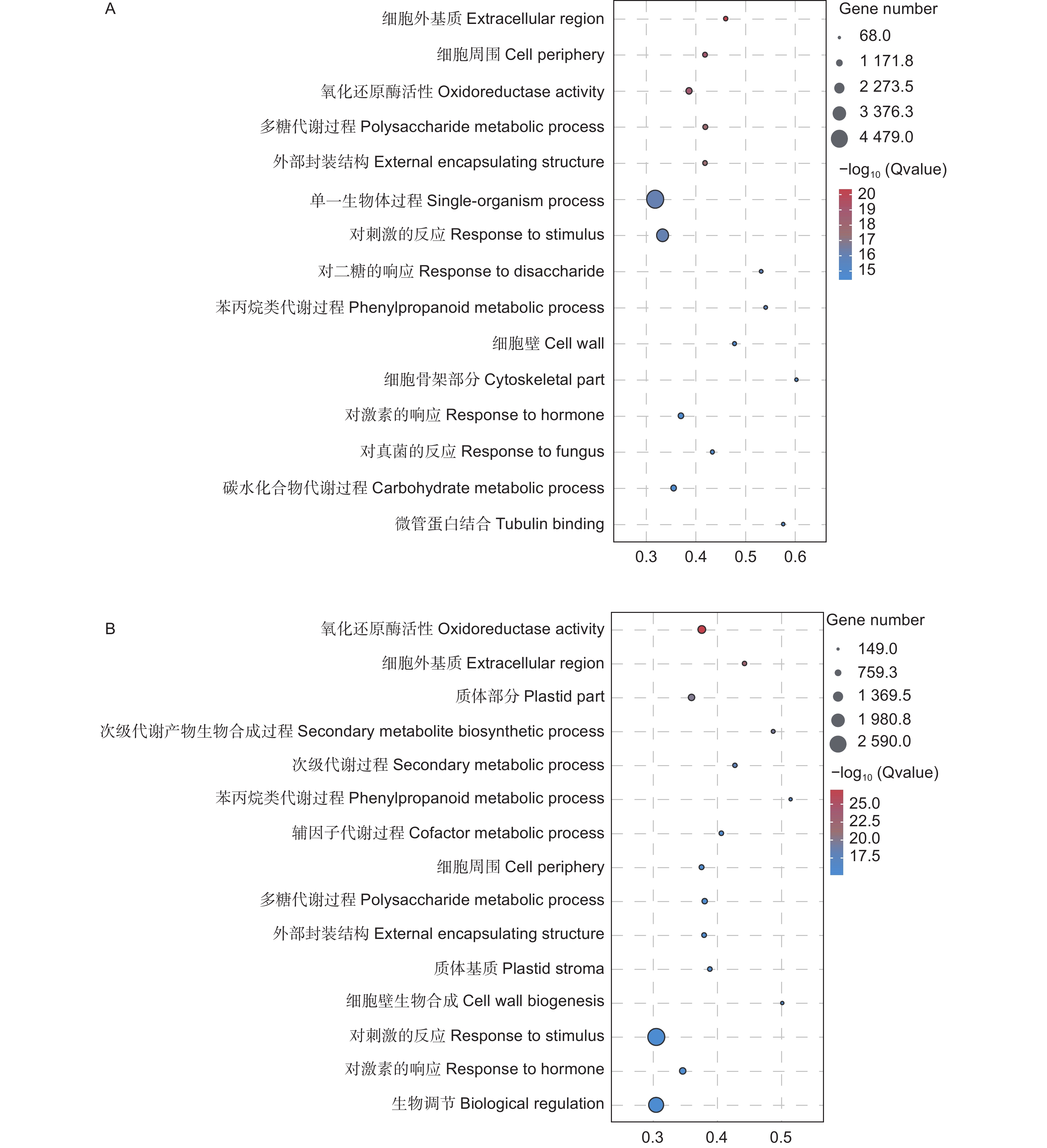
对差异基因进行GO功能富集分析,结果(图5)显示,在生物学过程、分子功能和细胞组分方面均存在显著富集。接菌48 h后,差异表达基因主要富集在单一生物体过程(Single-organism process)、对刺激反应(Response to stimulus)和碳水化合物代谢过程(Carbohydrate metabolic process)等过程。接菌72 h后,则主要富集在对刺激的反应(Response to stimulus)、生物调节(Biological regulation)和对激素的响应(Response to hormone)等通路。
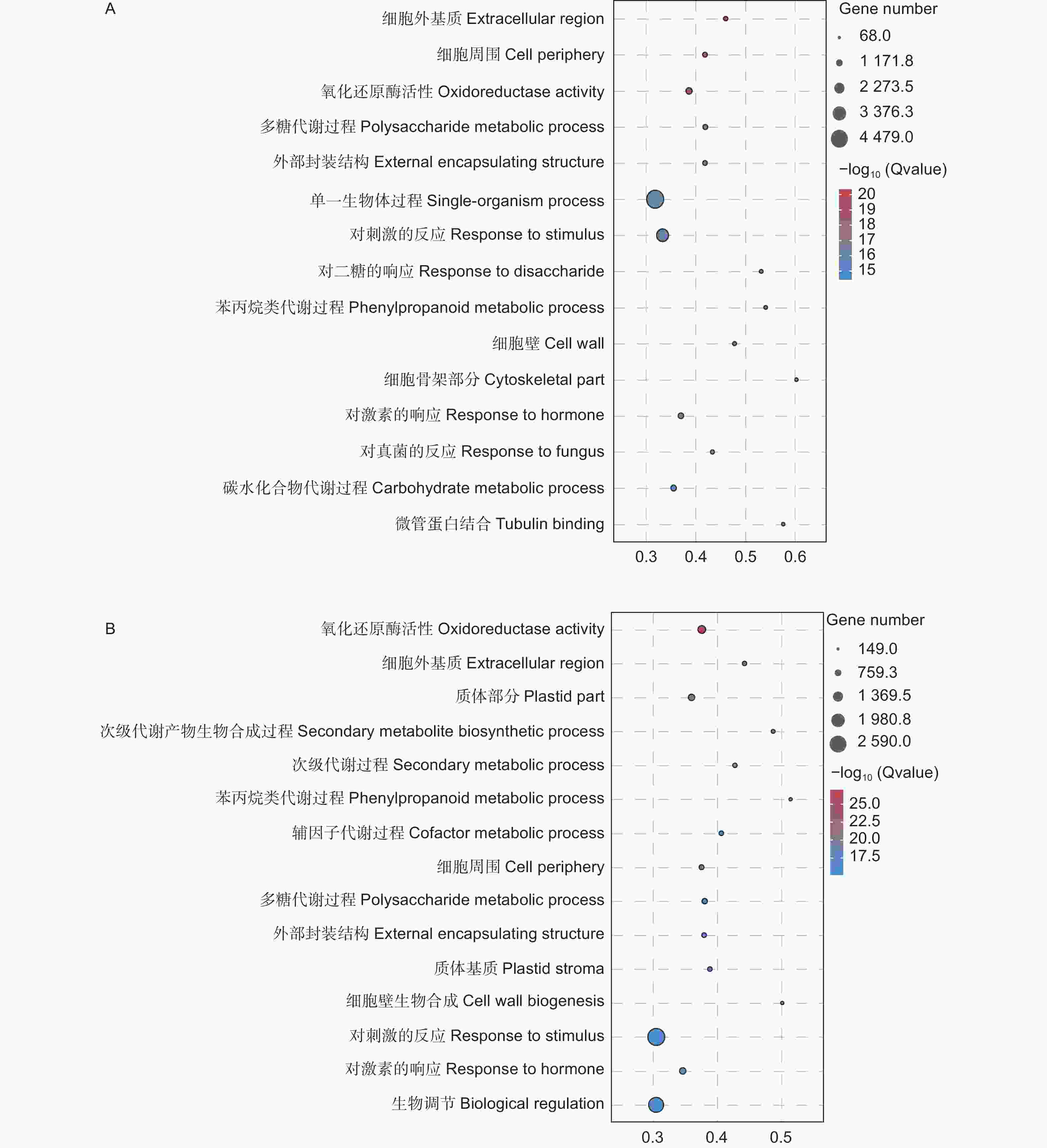
图 5 差异表达基因的GO富集分析
Figure 5. GO enrichment analysis of differentially expressed genes
进一步对差异基因进行KEGG富集分析,结果显示,接菌48 h后,92个差异基因富集到糖酵解/糖异生(Glycolysis / Gluconeogenesis)途径,127个差异基因富集到苯丙烷生物合成途径(Phenylpropanoid biosynthesis),147个差异基因富集到碳代谢途径(Carbon metabolism)、40个差异基因富集到果糖和甘露糖代谢途径(Fructose and mannose metabolism),144个差异基因富集到植物激素信号传递途径(Plant hormone signal transduction)。接种病原菌72 h后,124个差异基因富集到苯丙烷生物合成途径,76个差异基因富集到糖酵解/糖异生途径,128个差异基因富集到植物激素信号传递通路,122个差异基因富集到碳代谢通路、69个差异基因富集到淀粉和蔗糖代谢(Starch and sucrose metabolism)通路(图6)。
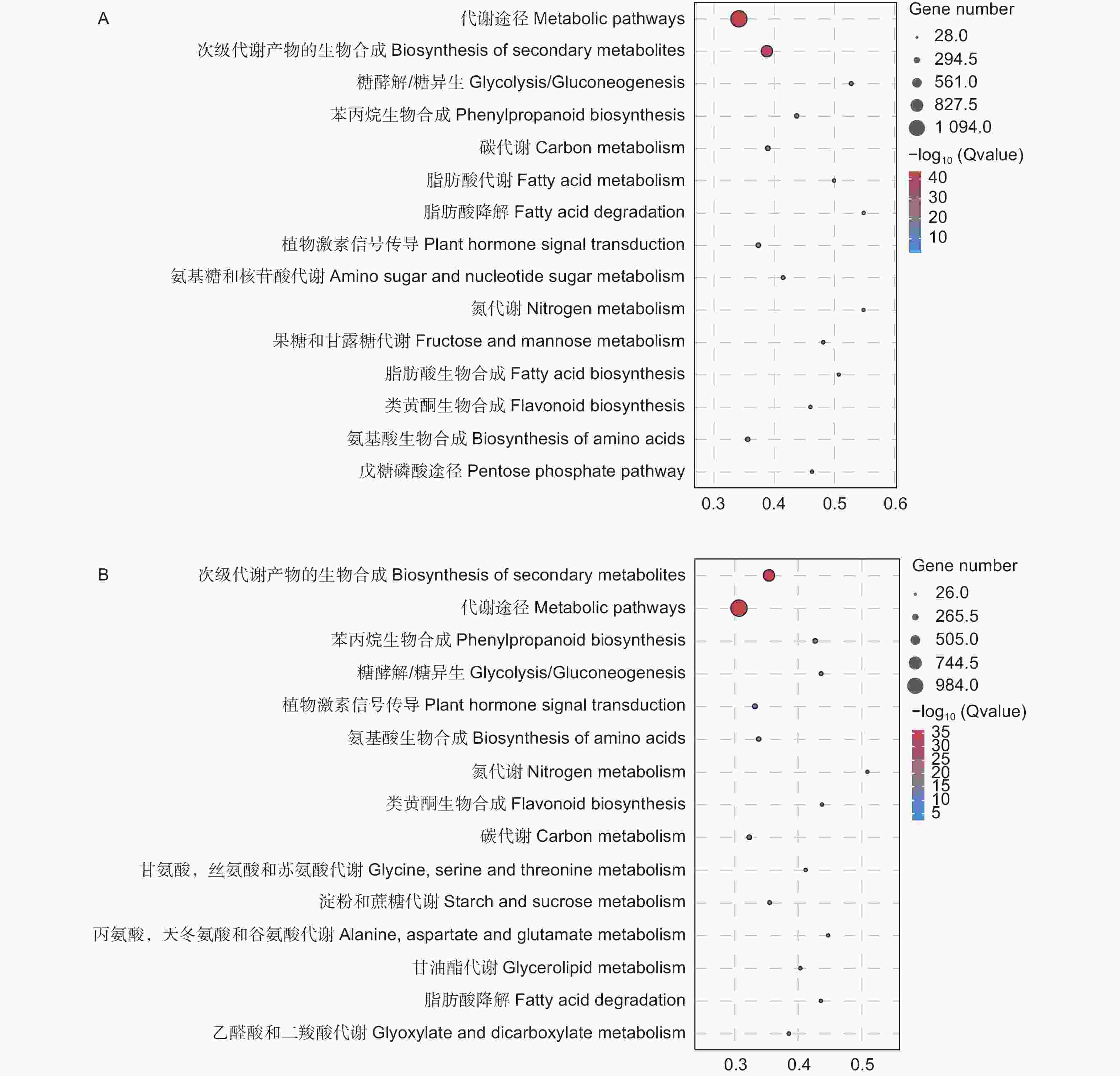
图 6 差异表达基因KEGG通路分析
Figure 6. KEGG pathway analysis of DEGs
-
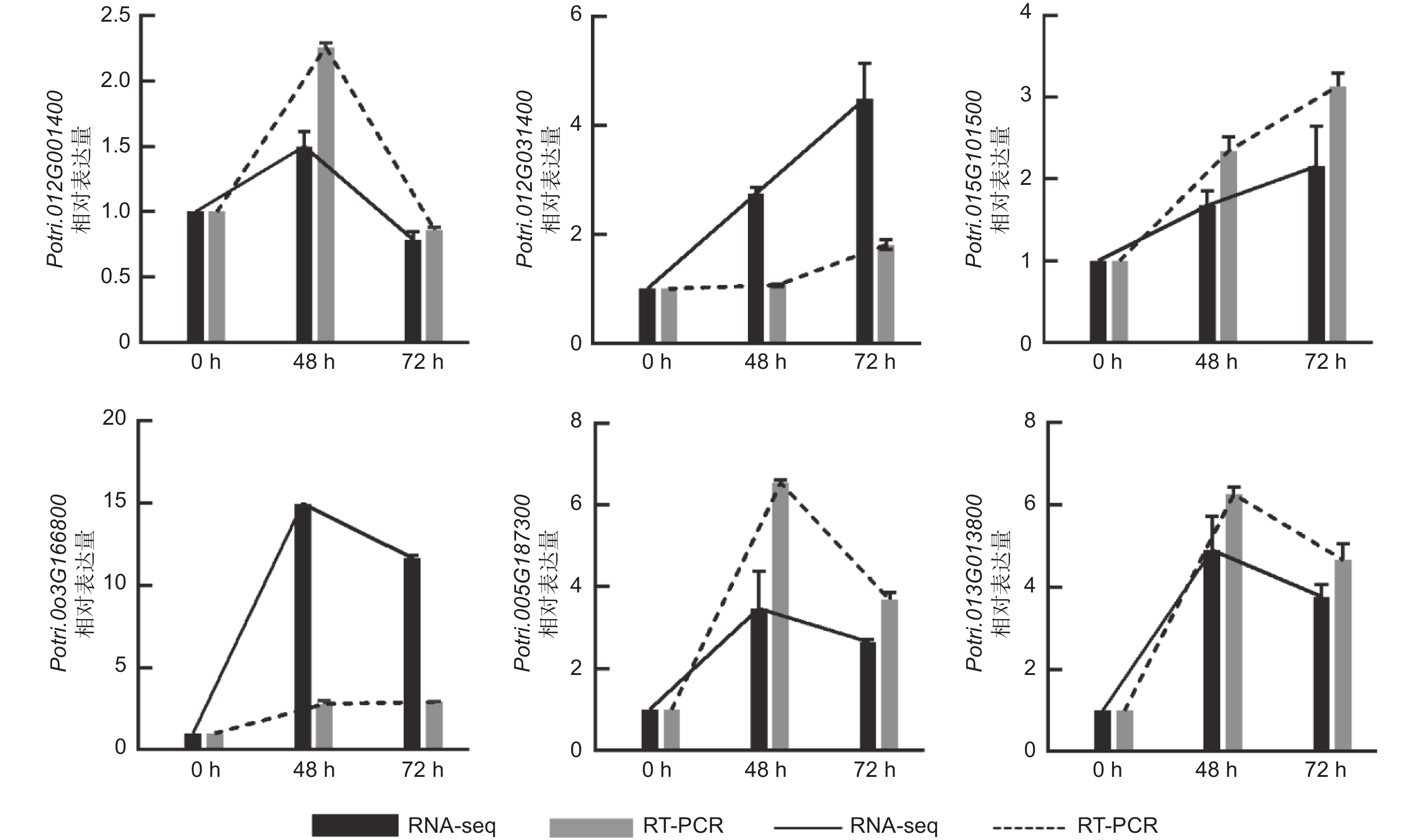
为了进一步验证转录组测序结果的准确性,本研究选取6个基因进行荧光定量PCR验证,即Potri.005G187300、Potri.012G031400、Potri.015G101500、Potri.003G166800、Potri.013G013800、Potri.012G001400,选取PagActin为内参基因。结果显示,6个基因的表达趋势与转录组测序结果基本一致(图7),表明本研究中分析获得的差异表达基因可信度较高,可用于后续实验。
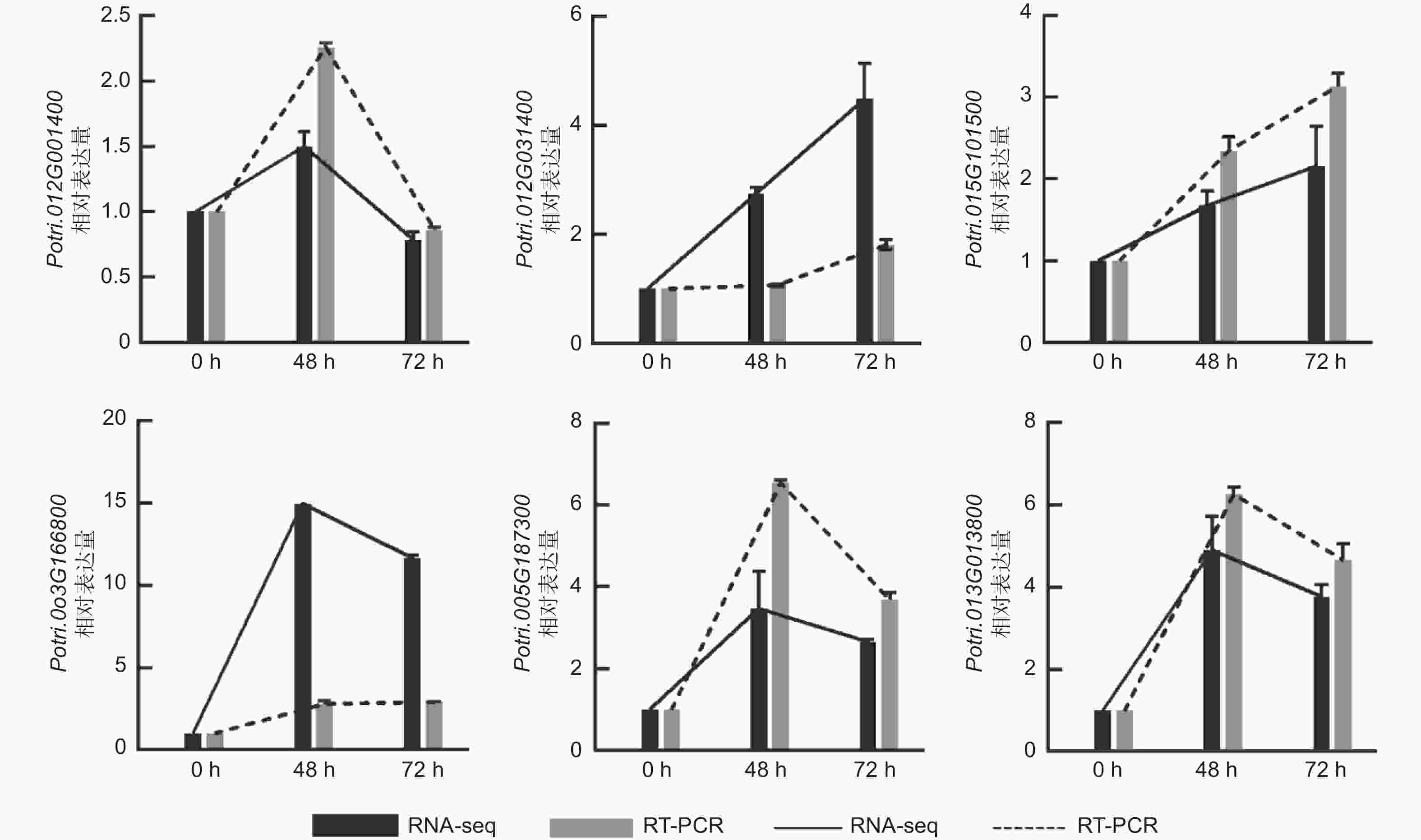
图 7 qRT-PCR验证差异表达基因
Figure 7. Validation of DEGs using qRT-PCR
-
KEGG富集通路结果显示,接菌48 h和72 h后差异基因显著富集到多个与糖合成与代谢相关的途径。植物SWEETs家族是一类重要的糖转运蛋白,在植物和病原菌互作过程中,不仅可以作为病原菌效应子的识别受体,还可以通过负责糖的运输影响植物本身的免疫反应[18]。之前的研究表明,杨树SWEET家族共有27个家族成员[19]。本研究共检测到26个杨树SWEET基因,其中48 h后10个上调、10个下调,72 h后11个上调、10个下调。9个SWEET基因在病原菌侵染48 h和72 h后均上调表达(表3)。因此,推测SWEETs可能在杨树与腐皮镰刀菌的互作过程中发挥重要作用。
表 3 杨树SWEETs基因表达情况
Table 3. Expression changes of poplar SWEET genes
基因名
Gene name基因号
Gene ID48 h表达变化
expression changes at 48 h72 h表达变化
expression changes at 72 hSWEET1a Potri.002G072600 down down SWEET1b Potri.002G072800 down down SWEET1c Potri.002G072700 — — SWEET1d Potri.005G187300 up up SWEET2a Potri.001G383000 up down SWEET2b Potri.011G103600 down down SWEET2c Potri.001G355500 down down SWEET3b Potri.015G021900. down down SWEET3c Potri.012G031400 up up SWEET4 Potri.001G344300 up up SWEET5 Potri.015G074300 — up SWEET7 Potri.003G143100 up up SWEET9 Potri.019G030500 — — SWEET10a Potri.015G101400 — — SWEET10b Potri.015G101500 up up SWEET10c Potri.015G101600 — — SWEET10d Potri.012G103200 — — SWEET11 Potri.015G101700 down up SWEET15a Potri.001G060900 up up SWEET15b Potri.003G166800 up up SWEET16a Potri.013G013800 up up SWEET16b Potri.013G013900 up up SWEET17a Potri.013G014500 down down SWEET17b Potri.013G014400 down down SWEET17c Potri.005G023900 down down SWEET17d Potri.008G220600 down down 注:“-” 表示与对照组的结果相比,基因表达未发生显著变化
Notes: “-” means that the gene expression in the inoculated grouped did not changed significantly compared with that of control group -
进一步对KEGG富集结果分析发现,植物激素信号转导途径中乙烯、水杨酸和茉莉酸信号通路中的基因受腐皮镰刀菌不同程度诱导。接菌后,乙烯合成限速酶基因ACS6表达量上调2.1~2.6倍,乙烯受体ETR 3个成员Potri.002G201500、Potri.008G164400和Potri.010G07300上调表达1.8~3.0倍。乙烯信号转导途径关键成员EIN3和ERF1/2的表达水平被腐皮镰刀菌诱导,前者在接菌48 h显著升高,后者在48 h和72 h均显著上调。乙烯信号通路重要调控因子MPK6的编码基因表达量显著下降。水杨酸信号通路中,水杨酸受体NPR1的编码基因(Potri.005G206100)表达量和关键转录因子TGA的编码基因(Potri.002G090700)在接菌48 h显著上调。该通路标志基因PR1的表达水平在接菌后48 h和72 h均显著上调。茉莉酸酰氨基酸结合物合成酶JAR1编码基因(Potri.002G168200)受腐皮镰刀菌诱导后表达量下调,茉莉酸信号通路关键调节因子JAZ、MYC2在接菌后48 h和72 h均显著下调表达(图8)。
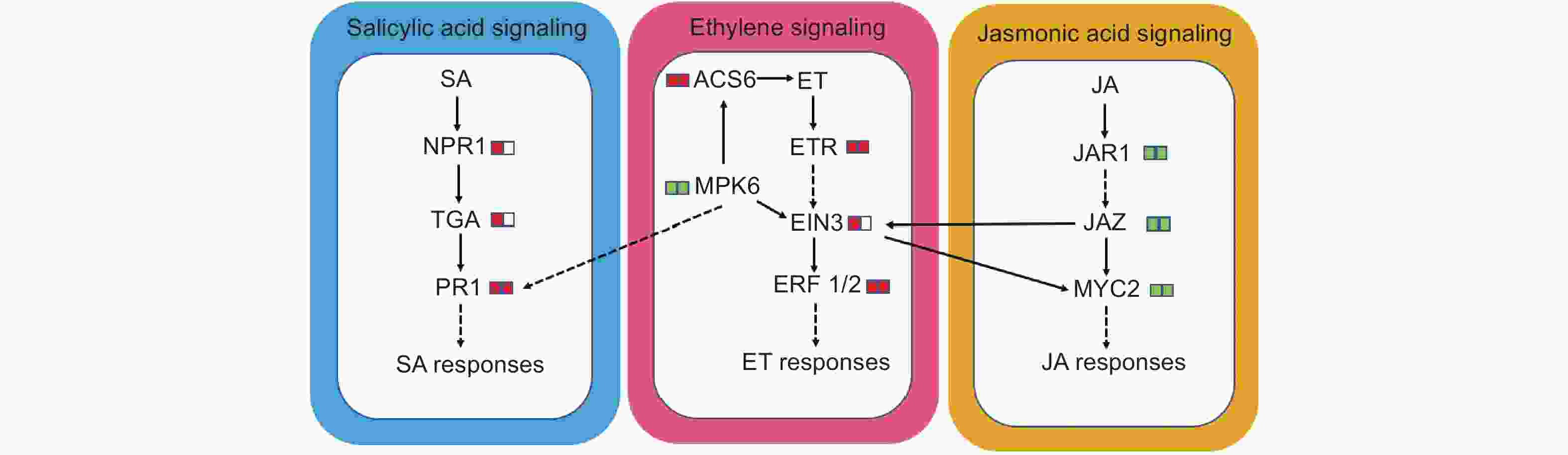
图 8 乙烯、茉莉酸和水杨酸途径中的差异表达基因
Figure 8. DEGs in ethylene, jasmonic acid and salicylic acid signaling pathway
-
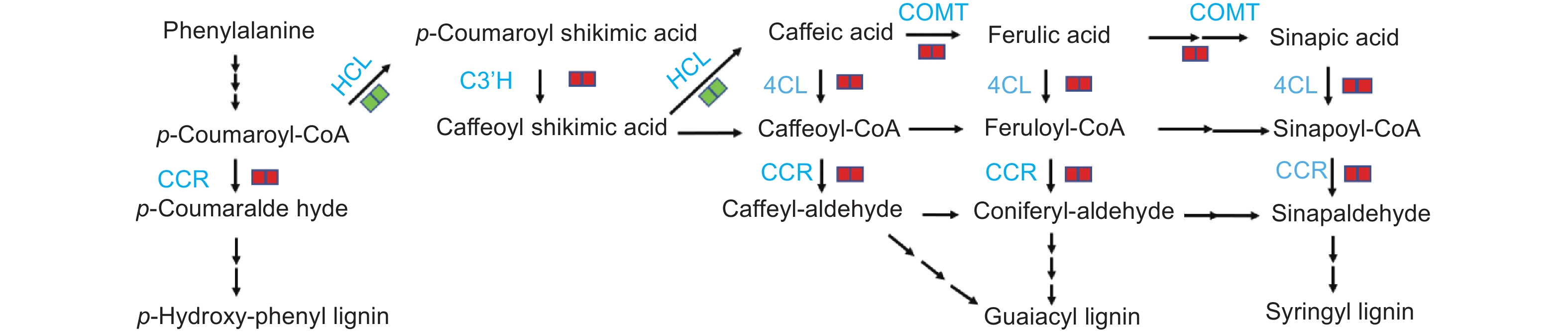
KEGG富集结果分析显示,杨树在响应腐皮镰刀菌入侵的过程中,苯丙烷类生物合成途径中关键酶基因的表达量发生显著变化。肉桂酰辅酶 A 还原酶CCR 为该途径第一个限速酶,同时催化3种羟基肉桂酸CoA酯的还原反应,生成相应的肉桂醛。接菌后48 h和72 h,CCR 基因表达量分别为0 h的1.4~10.5倍和1.7~10.5倍。咖啡酸-O-甲基转移酶 COMT 催化苯丙素类化合物羟基上氧原子的甲基化生成阿魏酸、芥子酸,受腐皮镰刀菌诱导后表达量上调4.0~4.5倍。4-香豆酸辅酶A连接酶 4CL 催化肉桂酸及其衍生物生成相应的辅酶A酯,在接菌后48 h和72 h,表达量分别上调1.3倍和1.7倍。香豆酸-3-羟基化酶 C3’H 催化对香豆酰莽草酸/奎宁酸到咖啡酰莽草酸/奎宁酸C3位置的羟基化反应,在接菌后表达量显著升高。莽草酸O-羟基肉桂酰转移酶 HCT 催化对香豆酰CoA生成对香豆酰莽草酸,在接菌后48 h和72 h分别下调1.5~8.2倍和1.3~8.2倍(图9)。
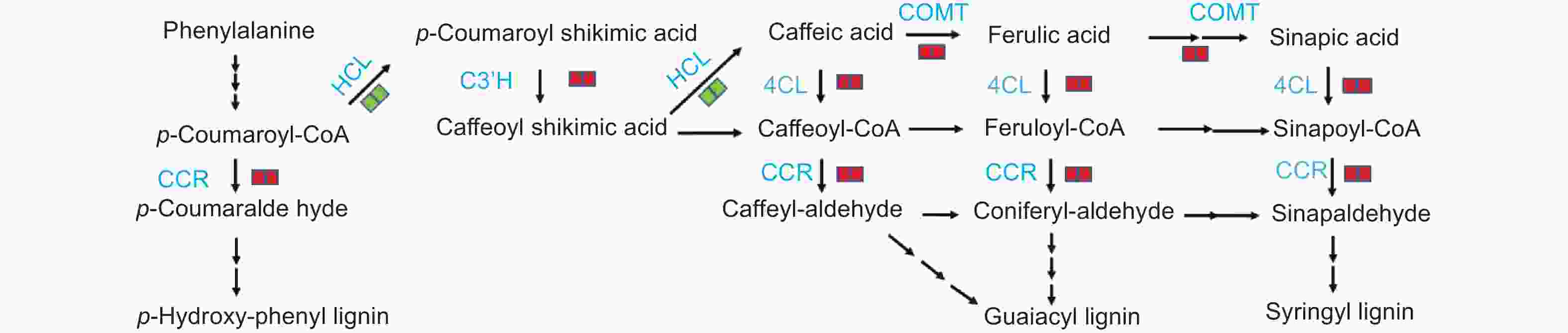
图 9 苯丙烷生物合成途径中的差异表达基因
Figure 9. DEGs in phenylpropanoid biosynthesis pathway
-
由腐皮镰刀菌引起的杨树病害是近几年江苏、安徽等南方地区发生较为严重的杨树病害,主要发病于新生幼苗,一旦发病,造成的经济损失严重[20]。从分子方面研究其发病机理,可为杨树抗病新品种的培育提供理论基础,但是目前相关研究仍十分有限。本研究利用RNA-seq技术对该病菌侵染杨树根部后不同时间的差异表达基因进行了分析,鉴定出杨树-腐皮镰刀菌互作过程关键基因、相关信号通路及代谢途径,为进一步揭示杨树对该致病菌的抗性分子机理奠定了基础。
对腐皮镰刀菌侵染杨树后差异表达基因的GO和KEGG代谢途径富集分析发现,糖酵解、果糖和甘露糖代谢等糖代谢途径发生显著变化。糖代谢是植物和微生物能量获取的主要方式,也可为二者细胞内生物合成提供物质基础,产生的部分糖还可作为信号分子调控寄主防御反应[21]。糖转运是植物-病原菌互作过程中伴随糖代谢的重要事件。研究表明,植物遭受病原菌侵染后,二者会发生营养争夺,竞争调控糖在细胞内外的转运方向,负责糖运输的糖转运蛋白在此过程中发挥重要作用[22]。SWEET是植物中广泛存在的一类糖转运蛋白。对拟南芥-腐霉病(Pythium irregulare)的研究发现,拟南芥(Arabidopsis thaliana (L.) Heynh.)AtSWEET2在根中高度表达,参与其与腐霉菌的互作过程,敲除该基因后促进了腐霉菌的进一步扩展,导致敏感性明显增强[23]。芜菁SWEET家族成员被根瘤菌(Plasmodiophora brassicae)接种诱导,可操纵寄主体内糖从合成部位到根瘤菌定植部位,为病菌的生长和扩繁提供营养,敲除SWEET11增强了寄主对根肿病的抗病性[24]。棉花(Gossypium hirsutum L.)GhSWEET42过表达植株在感染大丽轮枝菌(Verticillium dahliae)后具有高感病性,该基因的突变体则表现为抗病性[25]。由此可见,SWEET家族成员在不同的寄主-病原菌互作过程中所表现的作用不同。本研究分析表明,杨树27个SWEETs家族成员中有26个成员参与了对腐皮镰刀菌侵染的响应,其中SWEET1b等9个成员在接菌48 h和72 h后表达了均上调,SWEET1a等9个成员在接菌48 h和72 h后均下调,推测该基因家族在二者互作过程中发挥关键作用。后续,将通过进一步研究明确杨树SWEET不同成员在互作过程中的具体作用。
水杨酸、乙烯和茉莉酸信号通路不仅参与植物自身的生长发育,还调控植物的抗病反应,三者通过相互协同或拮抗的方式,使植物以最少的能量代价维持自身生长和防御各种病原微生物入侵[26]。在植物免疫反应中,通常认为水杨酸参与植物对活体营养型病原菌的抗性,乙烯与茉莉酸协同作用诱导植物对死体营养型病原的抗性[27]。对作物类土传真菌类病害的研究表明,乙烯信号通路参与棉花-枯萎病菌(Verticillium dahliae)、水稻-纹枯病菌(Rhizoctonia solani)的互作过程。棉花枯萎病菌的侵染可诱导乙烯信号通路下游关键调控因子GbERF1-like的表达,从而激活PR3、PR4等病程相关基因,引发抗病反应[28-29]。在水稻(Oryza sativa L.)中过表达乙烯合成酶基因OsACS2,可促进内源乙烯水平的积累,激活乙烯信号通路,提高了对纹枯病的抗性[30]。番茄枯萎病菌(Fusarium oxysporum)的侵染可诱导番茄水杨酸和乙烯信号通路标志基因显著上调,推测两条通路共同参与番茄-枯萎病菌的互作[31]。对杨树叶锈病的研究发现,水杨酸信号通路可通过提高叶片中儿茶素、原花青素等次生代谢物的含量,增强杨树对叶锈病的抗性[7]。上述研究表明,植物防御信号转导受植物基因型、与病原菌的互作及环境等因素的影响,不同信号转导通路在传递信号时也可能会存在相互交叉或重叠[32]。本研究发现,接种腐皮镰刀菌后,杨树根中乙烯受体ETR、乙烯信号通路主要组分EIN3、ERF1、ERF2上调表达,乙烯通路负调控因子MPK6下调表达。水杨酸信号通路关键基因上调表达,茉莉酸信号通路关键调节因子下调表达。推测乙烯信号通路在杨树-腐皮镰刀菌互作中起关键作用,水杨酸通路可能协同乙烯通路共同参与了杨树对腐皮镰刀菌的防御反应。
苯丙烷生物合成途径是植物体内物质合成与代谢的重要途径,也是植物抗性防御系统的重要组成部分。该途径产生的酚类、黄酮类等代谢产物可以抑制病原菌的生长,同时具有强大的抗氧化作用,帮助植物抵御自由基的伤害,在抗病过程中起化学屏障作用[33-34]。对杨树-叶锈菌互作的研究表明,叶锈菌侵染可诱导原花青素等化学物质的积累,从而阻止病斑的进一步扩展[35]。苯丙烷生物合成途径的另一个重要产物是木质素,是由酚类物质经过复杂过程结合而成的多聚体,也是植物细胞壁的主要成分[36]。当植物遭受病原菌侵染后,细胞壁会积累大量的木质素。细胞壁的木质化可以阻止病原菌的扩散,减少真菌溶解酶类对植物细胞的渗透和毒害作用,为抵抗病原菌的入侵提供了物理屏障[37]。研究发现,杨树溃疡病菌(Dothiorella gregaria)及其菌丝体提取物能诱导杨树细胞壁木质素的积累,而且抗病品种中木质素积累的速度和水平远远高于感病品种[38]。对土传病害的研究表明,棉花在应对大丽轮枝菌入侵时,Gh4CL、GhC3H、GhCCR4等木质素合成相关基因的表达量均显著提高,抗黄萎病棉花品种中的木质素含量明显高于感病棉花品种[39-40]。烟草经青枯病菌(Ralstonia solanacearum)侵染可诱导抗病品种中CCoAOMT、4CL、CCR等木质素合成相关基因上调表达[41]。本研究发现,CCR、4CL、C3’H和COMT等与木质素合成相关的基因被腐皮镰刀菌诱导上调表达,推测细胞壁增厚可能是杨树应对该病原菌侵染的策略之一。
-
本研究利用转录组测序技术鉴定到327个腐皮镰刀菌侵染诱导的差异表达基因,这些基因主要富集到糖酵解/糖异生、植物激素信号转导、苯丙烷生物合成、果糖和甘露糖代谢以及淀粉和蔗糖代谢等通路上。进一步的分析发现,杨树SWEETs家族中有9个家族成员在侵染48 h和72 h后均上调表达,乙烯信号通路关键成员除EIN3在侵染发生48 h时上调表达,ETR、ERF1、ERF2在侵染48 h和72 h后均上调表达,木质素合成关键酶基因CCR、4CL、C3’H、COMT上调表达,推测糖代谢和转运、乙烯信号通路、木质素积累等可能是杨树响应腐皮镰刀菌侵染的主要事件。后续将对挖掘到的关键候选基因进行功能验证。
杨树响应腐皮镰刀菌侵染的转录组学分析
Transcriptome analysis of Populus in response to Fusarium solani (Mart.) Sacc inoculation
-
摘要:
目的 基于RNA-Seq测序技术,初步探究了杨树-腐皮镰刀菌(Fusarium solani (Mart.) Sacc)互作过程中相关基因的表达、主要信号通路和代谢途径,筛选了杨树防御腐皮镰刀菌侵染的关键基因,为进一步揭示此类病害的分子机制奠定基础。 方法 以生长2个月的银腺杨84K为材料,用1 × 107 个·mL−1的腐皮镰刀菌孢子液分别侵染根系,侵染0 h(对照组)、48 h和72 h(接菌组)后取根组织进行转录组测序,挖掘杨树响应腐皮镰刀菌侵染的相关基因。 结果 (1)与对照组0 h相比,侵染48 h和72 h分别检测到8 939个和8 246个DEGs(Differentially Expressed Genes)。(2)GO分析发现,差异表达基因主要富集在在单一生物体过程、对刺激反应、碳水化合物代谢、生物调节和对激素响应等过程。(3)KEGG分析表明,糖酵解/糖异生、碳代谢、植物激素信号转导和苯丙烷生物合成等途径在病原菌侵染后发生显著变化。(4)杨树糖转运蛋白SWEETs家族中21个成员的表达受腐皮镰刀菌侵染诱导。乙烯受体ETR、乙烯信号通路主要组分EIN3、ERF1、ERF2上调表达,木质素合成关键酶基因CCR、4CL、C3’H、COMT表达量显著升高。 结论 杨树可能通过调节体内糖代谢和转运,激活乙烯信号通路,促进木质素积累、细胞壁增厚等策略,响应腐皮镰刀菌的侵染。 Abstract:Objective To study the differentially expressed genes, related signaling pathways and metabolic pathways between poplar- Fusarium solani (Mart.) Sacc interactions, and screen the key genes of poplar against Fusarium. solani (Mart.) Sacc inoculation for laying a foundation for further revealing the molecular mechanism of this disease. Methods The roots of 2-month-old seedlings of 84K were inoculated with 1 × 107 spores·mL−1 of F. solani (Mart.) Sacc spore solution. Zero-hour (control group), 48 h and 72 h (inoculated groups) after inoculation, root tissues were taken for transcriptome sequencing to explore genes related to poplar response to F. solani (Mart.) Sacc infection. Result (1) Compared with the control group, 8939 and 8246 DEGs (Differentially Expressed Genes) were detected in 48 h and 72 h inoculated samples, respectively. (2) GO analysis found that the differentially expressed genes were mainly enriched in single-organism process, response to stimulus, carbohydrate metabolism process, biological regulation and response to hormone. (3) KEGG pathway enrichment analysis showed that the DEGs were significantly enriched in glycolysis/gluconeogenesis, carbon metabolism, plant hormone signal transduction and phenylpropanoid biosynthesis. (4) The expressions of 21 genes in the SWEETs family of poplar were induced by F. solani (Mart.) Sacc. Ethylene receptor ETR, and the main components of the ethylene signaling pathway EIN3, ERF1, ERF2 were up-regulated. The expression levels of key enzyme genes involved in lignin synthesis including CCR, 4CL, C3’H and COMT were significantly increased. Conclusion Poplar may respond to the invasion of F. solani (Mart.) Sacc by regulating sugar metabolism and transport, activating the ethylene signaling pathway, promoting lignin accumulation, and thickening cell wall. -
Key words:
- Populus
- / Fusarium solani (Mart.) Sacc
- / inoculation process
- / transcriptome
-
图 1 腐皮镰刀菌(F. solani (Mart.) Sacc)侵染84K杨幼苗情况
Figure 1. Colonies of F. solani (Mart.) Sacc and the inoculated 84K seedlings
图 3 杨树接种腐皮镰刀菌后差异表达基因火山图
Figure 3. Volcano map of DEGs in poplar root inoculated with F. solani (Mart.) Sacc
图 8 乙烯、茉莉酸和水杨酸途径中的差异表达基因
Figure 8. DEGs in ethylene, jasmonic acid and salicylic acid signaling pathway
表 1 qRT-PCR 引物序列信息
Table 1. Primer sequences used for qRT-PCR analysis
序号 No. 基因号 Gene ID 引物序列 Primer sequence 1 Potri.005G187300 F:GTAGAGACCCTTTCGTTGCC R:CTTTTCCGGGACCAATCTCTAG 2 Potri.012G031400 F:CATGATCACCACCATCGCAA R:CGCGAGAAAGAGATCATGGC 3 Potri.015G101500 F:GTCTGGTCTGTCTTGCTCCT R:TGGTGATGAGAAGGATGGCA 4 Potri.003G166800 F:TCTTCCGAACATCTTGGGCT R:ACGTCAGGTTTTGCATCGAC 5 Potri.013G013800 F:GAGTGCCGAATGTTGTTGGT R:TCAGATTGCCAGCTTCCTCA 6 Potri.012G001400 F:CTGTCGCAGCATCAGGAAAT R:CTGCAGCGCTATCTTGTGAG 7 PagActin F:ACCCTCCAATCCAGACACTG R:TTGCTGACCGTATGAGCAAG  下载: 导出CSV
下载: 导出CSV
表 2 转录组测序数据质控分析
Table 2. Quality test of transcriptome sequencing data
样品
Sample原始数据量
Raw reads/bp高质量数据量
Clean reads/bp准确率
Clean ratio/%Q20
Content of Q20/%Q30
Content of Q30/%GC含量
Content of GC/%0 h-1 40 228 010 40 032 220 99.51 97.89 93.63 44.19 0 h-2 41 478 100 41 262 044 99.48 97.76 93.37 44.14 0 h-3 42 237 814 42 014 800 99.47 97.76 93.39 44.05 48 h-1 40 187 478 40 029 556 99.61 97.94 93.75 45.24 48 h-2 37 900 752 37 782 074 99.69 98.11 94.18 45.38 48 h-3 41 773 292 41 533 374 99.43 97.61 93.05 45.43 72 h-1 37 472 500 37 366 426 99.72 98.08 94.19 45.71 72 h-2 38 707 252 38 547 366 99.59 97.74 93.41 45.79 72 h-3 47 140 310 46 972 756 99.64 97.86 93.67 45.47
下载: 导出CSV
表 3 杨树SWEETs基因表达情况
Table 3. Expression changes of poplar SWEET genes
基因名
Gene name基因号
Gene ID48 h表达变化
expression changes at 48 h72 h表达变化
expression changes at 72 hSWEET1a Potri.002G072600 down down SWEET1b Potri.002G072800 down down SWEET1c Potri.002G072700 — — SWEET1d Potri.005G187300 up up SWEET2a Potri.001G383000 up down SWEET2b Potri.011G103600 down down SWEET2c Potri.001G355500 down down SWEET3b Potri.015G021900. down down SWEET3c Potri.012G031400 up up SWEET4 Potri.001G344300 up up SWEET5 Potri.015G074300 — up SWEET7 Potri.003G143100 up up SWEET9 Potri.019G030500 — — SWEET10a Potri.015G101400 — — SWEET10b Potri.015G101500 up up SWEET10c Potri.015G101600 — — SWEET10d Potri.012G103200 — — SWEET11 Potri.015G101700 down up SWEET15a Potri.001G060900 up up SWEET15b Potri.003G166800 up up SWEET16a Potri.013G013800 up up SWEET16b Potri.013G013900 up up SWEET17a Potri.013G014500 down down SWEET17b Potri.013G014400 down down SWEET17c Potri.005G023900 down down SWEET17d Potri.008G220600 down down 注:“-” 表示与对照组的结果相比,基因表达未发生显著变化
Notes: “-” means that the gene expression in the inoculated grouped did not changed significantly compared with that of control group
下载: 导出CSV
-
[1] 马雪俊. 杨树主要病虫害的种类、为害及防治[J]. 种子科技, 2022, 40(17):99-101. [2] YUAN M H, NGOU B P M, DING P T, et al. PTI-ETI crosstalk: an integrative view of plant immunity[J]. Current Opinion in Plant Biology, 2021, 62: 102030. doi: 10.1016/j.pbi.2021.102030 [3] YUAN M H, JIANG Z Y, BI G Z, et al. Pattern-recognition receptors are required for NLR-mediated plant immunity[J]. Nature, 2021, 592(7852): 105-109. doi: 10.1038/s41586-021-03316-6 [4] SHANG-GUAN K K, WANG M, HTWE N M P S, et al. Lipopolysaccharides trigger two successive bursts of reactive oxygen species at distinct cellular locations[J]. Plant Physiology, 2018, 176(3): 2543-2556. doi: 10.1104/pp.17.01637 [5] SU Y Y, LI H G, WANG Y L, et al. Poplar miR472a targeting NBS-LRRs is involved in effective defence against the necrotrophic fungus Cytospora chrysosperma[J]. Journal of Experimental Botany, 2018, 69(22): 5519-5530. [6] 田呈明, 梁英梅, 康振生, 等. 杨树与栅锈菌互作的细胞学研究[J]. 林业科学, 2002, 38(3):87-93. [7] ULLAH C, TSAI C J, UNSICKER S B, et al. Salicylic acid activates poplar defense against the biotrophic rust fungus Melampsora larici-populina via increased biosynthesis of catechin and proanthocyanidins[J]. New Phytologist, 2018, 221(2): 960-975. [8] DANG F F, LIN J H, LI Y J, et al. SlWRKY30 and SlWRKY81 synergistically modulate tomato immunity to Ralstonia solanacearum by directly regulating SlPR-STH2[J]. Horticulture Research, 2023, 10(5): uhad050. doi: 10.1093/hr/uhad050 [9] SHIMIZU K, SUZUKI H, UEMURA T, et al. Immune gene activation by NPR and TGA transcriptional regulators in the model monocot Brachypodium distachyon[J]. The Plant Journal, 2022, 110(2): 470-481. doi: 10.1111/tpj.15681 [10] YANG Y L, LI H G, LIU M Y, et al. PeTGA1 enhances disease resistance against Colletotrichum gloeosporioides through directly regulating PeSARD1 in poplar[J]. International Journal of Biological Macromolecules, 2022, 214: 672-684. doi: 10.1016/j.ijbiomac.2022.06.099 [11] YE S L, JIANG Y Z, DUAN Y J, et al. Constitutive expression of the poplar WRKY transcription factor PtoWRKY60 enhances resistance to Dothiorella gregaria Sacc. in transgenic plants[J]. Tree Physiology, 2014, 34(10): 1118-1129. doi: 10.1093/treephys/tpu079 [12] 李伶俐, 韩正敏, 吕明亮, 等. 苏北地区杨树枯萎病病原菌的鉴定[C]. 江苏省植物病理学会. 第十一次会员代表大会暨学术研讨会, 中国江苏扬州, F, 2008 . [13] HAN M, XU X L, LI X, et al. New Insight into Aspartate Metabolic Pathways in Populus: Linking the Root Responsive Isoenzymes with Amino Acid Biosynthesis during Incompatible Interactions of Fusarium solani[J]. International Journal of Molecular Sciences, 2022, 23(12): 6368. doi: 10.3390/ijms23126368 [14] 叶小真, 杨泽慧, 张清华, 等. 桉树枯萎病菌Fusarium solani分子检测技术研究[J]. 森林与环境学报, 2019, 39(6):629-635. [15] Vettraino A M, Shrestha G P, Vannini A. First Report of Fusarium solani Causing Wilt of Olea europaea in Nepal[J]. Plant Disease, 2009, 93(2): 200. [16] 王新荣 李 楠, 林月娥, 等. 苏铁镰刀菌球茎腐烂病化学防治技术研究[J]. 中国森林病虫, 2005, 24(6):34-37. [17] 陈世荣, 王建华, 储祥宏, 等. 杨树枯萎病的发生与防治初探[J]. 江苏林业科技, 2013, 40(4):31-33. [18] CHEN L Q, HOU B H, LALONDE S, et al. Sugar transporters for intercellular exchange and nutrition of pathogens[J]. Nature, 2010, 468(7323): 527-532. doi: 10.1038/nature09606 [19] ZHANG L, WANG L J, ZHANG J, et al. Expression and localization of SWEETs in Populus and the effect of SWEET7 overexpression in secondary growth[J]. Tree Physiology, 2021, 41(5): 882-899. doi: 10.1093/treephys/tpaa145 [20] 游 昕. 茄镰孢菌(Fusarium solani)对欧美杨细菌性溃疡病发生的影响分析 [D]. 北京: 北京林业大学, 2013. [21] ROLLAND F, BAENA-GONZALEZ E, SHEEN J. SUGAR SENSING AND SIGNALING IN PLANTS: Conserved and Novel Mechanisms[J]. Annual Review of Plant Biology, 2006, 57(1): 675-709. doi: 10.1146/annurev.arplant.57.032905.105441 [22] LEMONNIER P, GAILLARD C, VEILLET F, et al. Expression of Arabidopsis sugar transport protein STP13 differentially affects glucose transport activity and basal resistance to Botrytis cinerea[J]. Plant Molecular Biology, 2014, 85(4-5): 473-484. doi: 10.1007/s11103-014-0198-5 [23] CHEN H Y, HUH J H, YU Y C, et al. The Arabidopsis vacuolar sugar transporter SWEET2 limits carbon sequestration from roots and restricts Pythium infection[J]. The Plant Journal, 2015, 83(6): 1046-1058. doi: 10.1111/tpj.12948 [24] CHEN J J, PIAO Y L, LIU Y M, et al. Genome-wide identification and expression analysis of chitinase gene family in Brassica rapa reveals its role in clubroot resistance[J]. Plant Science, 2018, 270: 257-267. doi: 10.1016/j.plantsci.2018.02.017 [25] SUN M X, ZHANG Z Q, REN Z Y, et al. The GhSWEET42 glucose transporter participates in Verticillium dahliae infection in cotton[J]. Frontiers in Plant Science, 2021, 12: 690754. doi: 10.3389/fpls.2021.690754 [26] ROBERT-SEILANIANTZ A, GRANT M, JONES J D G. Hormone crosstalk in plant disease and defense: more than just JASMONATE-SALICYLATE antagonism[J]. Annual Review of Phytopathology, 2011, 49(1): 317-343. doi: 10.1146/annurev-phyto-073009-114447 [27] BARI R, JONES J D G. Role of plant hormones in plant defence responses[J]. Plant Molecular Biology, 2008, 69(4): 473-488. [28] 赵光栋. GhbHLH122影响乙烯的生物合成进而调控棉花对枯萎病抗性的分子机制研究 [D]. 泰安: 山东农业大学, 2022. [29] GUO W F, JIN L, MIAO Y H, et al. An ethylene response-related factor, GbERF1-like, from Gossypium barbadense improves resistance to Verticillium dahliae via activating lignin synthesis[J]. Plant Molecular Biology, 2016, 91(3): 305-318. doi: 10.1007/s11103-016-0467-6 [30] HELLIWELL E E, WANG Q, YANG Y N. Transgenic rice with inducible ethylene production exhibits broad-spectrum disease resistance to the fungal pathogens Magnaporthe oryzae and Rhizoctonia solani[J]. Plant Biotechnology Journal, 2012, 11(1): 33-42. [31] DI X T, GOMILA J, TAKKEN F L W. Involvement of salicylic acid, ethylene and jasmonic acid signalling pathways in the susceptibility of tomato to Fusarium oxysporum[J]. Molecular Plant Pathology, 2017, 18(7): 1024-1035. doi: 10.1111/mpp.12559 [32] LATTANZINO V, LATTANZINO V M T, CARDINALI A. Role of phenolics in the resistance mechanisms of plants against fungal pathogens and insects[J]. phytochemistry, 2006, 37(2): 23-67. [33] DONG N Q, LIN H X. Contribution of phenylpropanoid metabolism to plant development and plant–environment interactions[J]. Journal of Integrative Plant Biology, 2021, 63(1): 180-209. doi: 10.1111/jipb.13054 [34] 王硕. 西瓜枯萎病抗性反应中苯丙烷代谢及关键基因Cl4CL的克隆 [D]. 保定: 河北农业大学, 2021. [35] MANOELA M, G. RALPH S, MELLWAY R, et al. The transcriptional response of hybrid poplar (Populus trichocarpa x P. deltoides) to infection by Melampsora medusae leaf rust involves induction of flavonoid pathway genes leading to the accumulation of proanthocyanidins[J]. Molecular plant-microbe interactions:MPMI, 2007, 20(7): 816-831. doi: 10.1094/MPMI-20-7-0816 [36] UMEZAWA T. The cinnamate/monolignol pathway[J]. Phytochemistry Reviews, 2009, 9(1): 1-17. [37] LIU C, YU H S, VOXEUR A, et al. FERONIA and wall-associated kinases coordinate defense induced by lignin modification in plant cell walls[J]. Science advances, 2023, 9(10): eadf7714. doi: 10.1126/sciadv.adf7714 [38] 胡景江 朱 玮, 文建雷. 杨树细胞壁HRGP和木质素的诱导积累与其对溃疡病抗性的关系[J]. 植物病理学报, 1999, 29(2):151-156. [39] 吴立柱, 王省芬, 张 艳, 等. 酸不可溶性木质素和漆酶在棉花抗黄萎病中的作用[J]. 作物学报, 2014, 40(7):1157-1163. [40] ZHU Y T, HU X Q, WANG P, et al. GhODO1, an R2R3-type MYB transcription factor, positively regulates cotton resistance to Verticillium dahliae via the lignin biosynthesis and jasmonic acid signaling pathway[J]. International Journal of Biological Macromolecules, 2022, 201: 580-591. doi: 10.1016/j.ijbiomac.2022.01.120 [41] LI Y Y, WANG L, SUN G W, et al. Digital gene expression analysis of the response to Ralstonia solanacearum between resistant and susceptible tobacco varieties[J]. Scientific Reports, 2021, 11(1): 3887. doi: 10.1038/s41598-021-82576-8 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 615
- HTML全文浏览量: 191
- PDF下载量: 11
- 被引次数: 0
