

-
土壤酶是一类主要来源于土壤微生物的生物催化剂,是微生物参与生物地球化学循环过程的核心动力[1]。为了获取碳(C)、氮(N)、磷(P)等构成生命有机体的基础元素完成自身新陈代谢的需求,微生物将分泌相应的胞外酶转化土壤有机质中C、N、P元素[2],微生物分泌碳转化酶β-葡萄糖苷酶(BG)、氮转化酶β-乙酰葡糖胺糖苷酶(NAG)和磷转化酶磷酸单酯酶(AP)。通常认为,土壤酶活性变化指示了微生物养分需求特征和土壤养分状况[3-6]。
随着生态化学计量学理论的应用,反映微生物和土壤酶活性内在关系的土壤酶化学计量的概念被提出[7-8]。Sinsabaugh等[9]研究发现,在全球尺度上,土壤C、N、P酶化学计量比总体上呈1∶1∶1的关系,这一理论揭示了土壤微生物养分需求与土壤养分有效性之间生物地球化学平衡模式。土壤酶活性对土壤养分变化和环境胁迫响应极为敏感[3, 10],有研究认为,土壤酶生态化学计量比可以指示土壤微生物能量和养分资源限制状况及土壤质量、生物地球化学循环的特征[11-12]。Xu等[13]调查中国东南地区从南到北森林样带C、N、P养分获得酶活性及其化学计量特征发现,土壤酶C∶N和N∶P较低的比值指示了我国热带森林生态系统土壤微生物受到P养分元素的限制。近些年,从土壤酶化学计量角度分析土壤微生物的养分限制逐渐成为土壤生态学领域研究的热点话题之一。
林分是土壤养分的重要影响因素[14],但目前关于不同林分对土壤酶活性及其生态化学计量比影响特征的研究相对较少,其中,凋落物和根系是土壤养分的主要输入源,不同林分类型(如树种)的凋落物输入和根系变化影响着土壤养分的有效性[15-16]。左巍等[17]对青藏高原不同树种凋落物养分状况的研究发现,华北落叶松、云杉、白桦林之间凋落物养分状况差异显著。弓文艳等[18]对不同林分土壤C、N含量变化的研究表明,落叶松林土壤C、N和C/N比值显著高于油松林和落叶松-油松-刺槐混交林,不同林分类型土壤C、N含量之间的差异与其凋落物有关。另外,不同树种产生的根系分泌物也不相同[19]。土壤是土壤微生物直接的能量和养分来源,不同树种的凋落物和根系分泌物变化所引起的土壤养分变化也会带来土壤微生物对能量和养分需求的变化,那么不同树种土壤微生物能量和养分需求将如何变化?目前相关方面的研究报道相对不足,很大程度上限制了对微生物参与的生物地球化学过程的深入理解。因此,有必要开展林分对土壤酶及其生态化学计量比的研究,以进一步理解土壤微生物的养分需求特征。
神农架地处我国秦巴山地,是湖北境内长江与汉水的分水岭、南水北调中线工程重要的水源涵养地、三峡库区最大的天然绿色屏障,拥有当今世界中纬度地区保持完好的亚热带森林生态系统,生态地位十分重要,因而备受关注。神农架人工林现保存面积42605 hm2,占湖北省森林面积30%左右[20],其中,水杉(Metasequoia glyptostroboides Hu et W. C. Cheng)、柳杉(Cryptomeria japonica var. sinensis Miquel)、落叶松(Larix gmelinii (Ruprecht) Kuzeneva)和华山松(Pinus armandii Franch)是该地区的主要造林树种。因此,本研究选取神农架上述4种典型人工林土壤为研究对象,分析其土壤酶活性及其生态学计量比变化特征,并进一步分析土壤酶活性及其生态学计量比的影响因素,以期更好地认识本区域土壤微生物能量和养分资源的限制状况,为该区域林地养分循环的改善提供一定的理论参考。
-
神农架林区地处湖北西北部,由兴山县、房县、巴东县的边缘部分组成(109°56′~110°58′ E,31°15′~31°57′ N),海拔1 755 m。属大巴山脉东延之余脉,为我国中部山地与东部丘陵低山区的过渡地带,相对高差达2 700 m。属北亚热带季风气候,是亚热带气候向温带气候过渡区域,主要受东南季风影响,年平均气温10.6℃,年平均日照率42%,相对湿度73%~75%,平均年蒸发量500~800 mm,年降水量800~2 500 mm。降水多集中于夏季,冬季较少,4—l0月的降水量占全年的70%以上,降水量随海拔的增高而增加。随海拔升高,依次呈现出亚热带、暖温带、中温带等多种气候类型。植被分布具有明显的垂直地带性,随海拔升高依次为常绿阔叶林、常绿落叶阔叶混交林、落叶阔叶林、针阔混交林、针叶林及灌丛。土壤具有自然的垂直带,随海拔升高自下而上土壤类型依次为:山地黄棕壤、山地棕壤、山地暗棕壤、山地棕色针叶林土和山地草甸土[21-23]。
本研究选取湖北林科院神农架生态站(31°18′00″~31°42′00″ N、109°56′24″~110°42′36″ E,海拔1 580 m)附近的水杉、柳杉、落叶松和华山松纯林人工林为研究对象,4种林分类型林龄大约40 a,林下植被群落类型相近,人为干扰少,具体林分立地状况见表1。
表 1 4种林分的基本概况
Table 1. Basic characteristics of four types of forest stands
林分类型
Forest type海拔
Altitude/m坡度
Slope/(°)林分密度
Stand density/(株·hm−2)平均树高
Mean height of tree/m平均胸径
Mean diameter/cmMG 1 650 21~27 875±164 19.5 21.74 CF 1 640 9~15 792±206 24.5 20.45 LG 1 790 9~15 843±182 22.4 21.70 PA 1 580 3~7 956±139 12.0 14.68 注:MG: 水杉林;CF: 柳杉林;LG:落叶松林;PA:华山松林。下同
Notes:MG: Metasequoia glyptostroboides forest;CF:Cryptomeria japonica forest;LG:Larix gmelinii forest;PA:华山松林Pinus armandii forest.the same below. -
2019年8月生长季,在每个森林类型分别设置3个20 m×20 m的样方,样方间距>50 m。在每个样方内,利用土钻(直径4 cm)按照5点采样法,去除表面凋落物后,采集土壤0~10 cm层土壤样品。土样用低温保鲜箱运回实验室处理,同一个林分所有样方采集的土样进行均匀混合,并用镊子去除石砾、植物根系,过2 mm筛,一部分鲜样用于土壤含水量测定;一部分土样自然风干后用于土壤酶活性和pH测定;另一部分土壤经自然风干后,研磨过0.149 mm筛,用于土壤有机碳、总氮和总磷指标的测定。
-
土壤有机碳(SOC)含量采用重铬酸钾外加热氧化法测定;土壤总氮(TN)含量采用半微量开氏法测定;土壤总磷(TP)含量采用HClO4-H2SO4消煮,用流动分析仪(Bran Luebbe AA3, Hamburg, Germany)测定;pH值采用水土质量比为2.5∶1测定。每个样品重复测定3次。
-
土壤酶活性采用参考文献[24]中比色法测定。称取土壤5 g,根据测定相应的酶种类加入对应的提取液和缓冲液,经振荡、水浴加热和过滤后,进行测定,其中,磷酸单酯酶(AP)提取样品放入恒温箱(37℃)培养24 h后进行测定;土壤β-葡萄糖苷酶(BG)、β-乙酰葡糖胺糖苷酶(NAG)和AP测定的底物分别为对-硝基苯-β-D吡喃葡萄糖苷、对-硝基苯-β-D吡喃葡萄糖苷和磷酸苯二钠。
-
采用Turner等[25]的研究方法,将分解几丁质的相关酶作为氮循环的特征酶进行计算。BG:NAG、BG∶AP和NAG∶AP分别代表土壤酶C∶N、C∶P和N∶P。
单因素方差采用纽曼-科伊尔斯检验(SNK)分析不同林型土壤化学性质、土壤酶活性及其化学计量比之间的显著性差异,显著性水平设定为P<0.05。对酶的活性及其化学计量比进行主成分分析(PCA);土壤化学性质对土壤酶活性及其化学计量比的影响采用冗余分析(RDA);采用Pearson法进行土壤酶及其生态化学计量比与土壤化学性质的回归分析。采用Excel 2013和SPSS 19.0软件进行数据的分析和处理,采用Canoco5.0软件进行主成分分析(PCA)和冗余分析(RDA),采用Origin 2019软件绘图。
-
不同林分土壤化学性质间存在差异(表2),其中,柳杉林土壤SOC含量最高,且与水杉林和落叶松林的差异显著(P<0.05),华山松林土壤SOC含量也显著高于水杉林(P<0.05)。柳杉林土壤TN显著高于其他3个林分类型(P<0.05),其他3个林分间土壤TN含量差异不显著(P>0.05);柳杉林和华山松林土壤TP含量显著高于水杉林和落叶松林(P<0.05),柳杉林和华山松林土壤TP间差异显著(P<0.05),水杉林和落叶松林间土壤TP含量差异不显著(P>0.05);华山松林土壤C∶N最高,与其他3个林分C∶N之间差异显著(P<0.05),落叶松林和柳杉林土壤C∶N也显著高于水杉林(P<0.05);落叶松林土壤C∶P、N∶P显著高于其他3个林分(P<0.05),华山松林土壤C∶P、N∶P最低,显著低于水杉林和柳杉林(P<0.05),水杉林和柳杉林之间土壤C∶P、N∶P差异不显著(P>0.05);水杉林和柳杉林土壤pH值较高,与落叶松林和华山松林土壤pH值之间差异显著(P<0.05),落叶松林土壤pH值最低,与华山松林土壤pH值之间差异显著(P<0.05)。
表 2 4种林分类型土壤化学性质
Table 2. Soil chemical properties of four types of forest stands
林分类型
Forest type有机碳
SOC/(g·kg−1)全氮
TN/(g·kg−1)全磷
TP/(g·kg−1)碳氮比
C∶N碳磷比
C∶P氮磷比
N∶PpH值 MG 32.08±0.70 a 2.54±0.01 a 0.61±0.01 a 12.61±0.30 a 52.23±1.80 b 4.14±0.05 b 7.10±0.22 c CF 46.91±0.70 c 3.50±0.02 b 0.86±0.01 b 13.42±0.28 b 54.33±0.40 b 4.05±0.09 b 7.13±0.12 c LG 37.00±6.88 ab 2.82±0.53 a 0.57±0.16 a 13.12±0.04 b 66.38±8.31 c 5.06±0.62 c 5.53±0.05 a PA 40.89±0.70 bc 2.70±0.02 a 1.06±0.12 c 15.13±0.05 c 38.98±4.65 a 2.58±0.30 a 6.34±0.07 b 注:不同小写字母代表不同林分类型之间差异性显著(P<0.05),下同。
Notes: Different lowercase letters indicate significant difference among different forest stands (P<0.05). SOC:soil organic carbon;TN: total nitrogen;TP:total phosphorus;C:N:the ratio of carbon and nitrogen;C:P:the ratio of carbon and phosphorus;N:P:the ratio of nitrogen and phosphorus.the same below. -
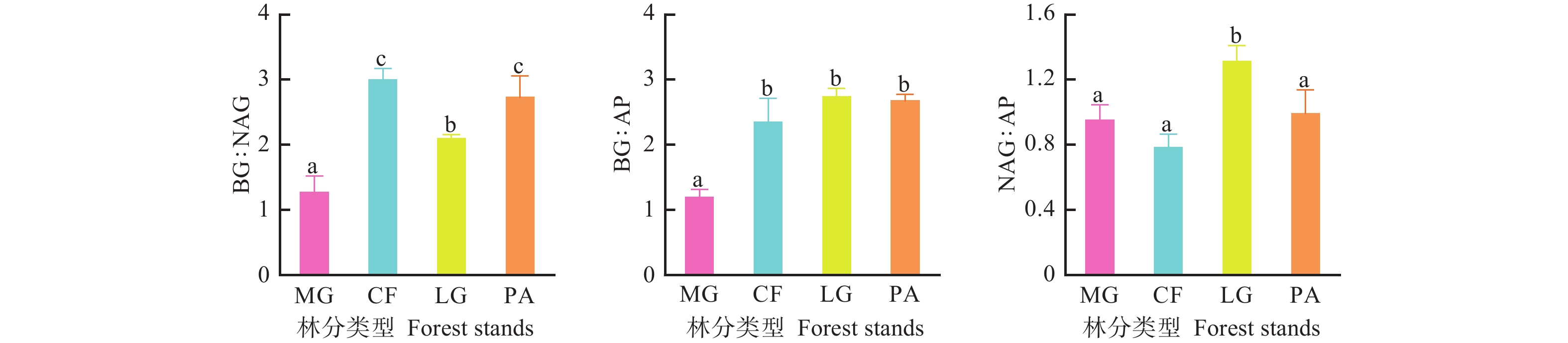
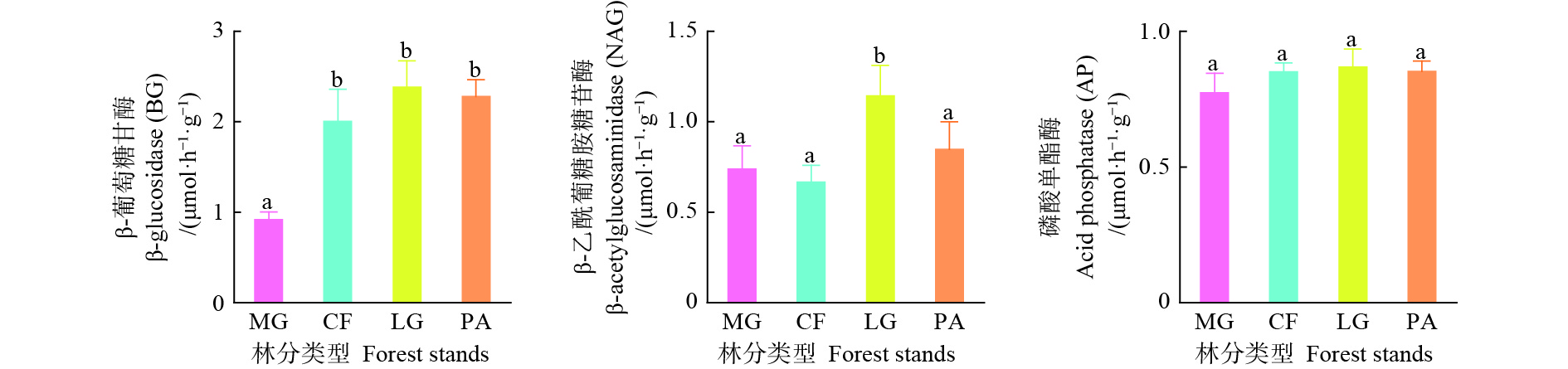
不同林分类型之间土壤酶活性存在差异(图1),落叶松林土壤酶活性最高,其中,水杉林BG活性显著低于其他3个林分(P<0.05),其他3个林分之间土壤BG活性差异性不显著(P>0.05);落叶松林土壤NAG活性显著高于其他3个林分(P<0.05),但其他3个林分间土壤NAG活性差异不显著(P>0.05);不同林分间土壤AP活性差异不显著(P>0.05)。
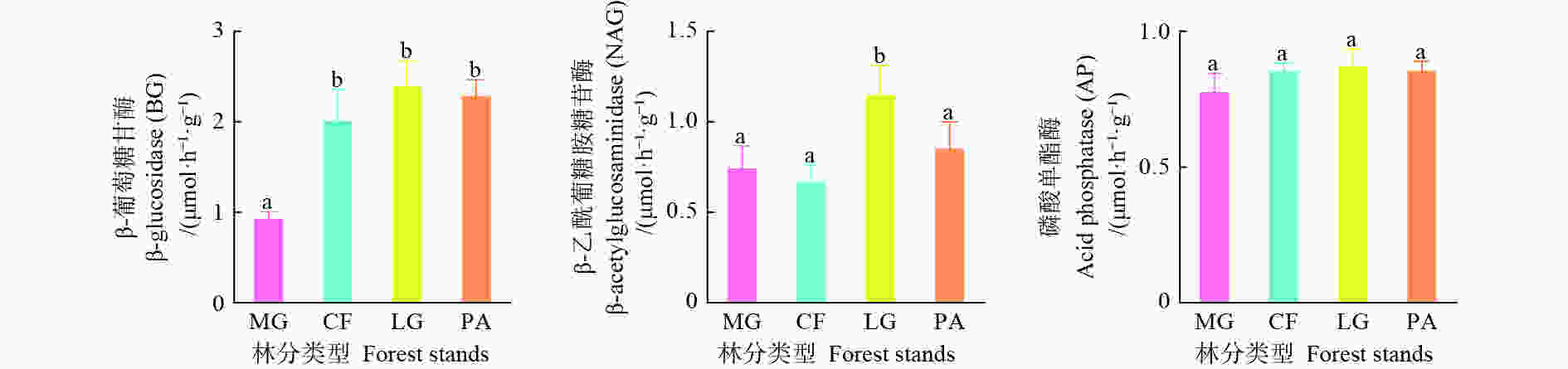
图 1 不同林分土壤酶活性变化特征
Figure 1. The various of soil enzyme activity in different forest stands
-
不同林分土壤酶的生态化学计量比存在差异(图2)。不同林分土壤酶BG∶NAG(酶C∶N)大小顺序为柳杉林>华山松林>落叶松林>水杉林,柳杉林和华山松林土壤酶BG∶NAG与落叶松林和水杉林间差异显著(P<0.05),落叶松林和水杉林间土壤酶BG∶NAG差异也显著(P<0.05);水杉林土壤酶BG∶AP显著低于其他3个林分(P<0.05),其他3个林分之间差异不显著(P>0.05);不同林分土壤酶NAG∶AP(酶N∶P)大小顺序为落叶松林>华山松林>水杉林>柳杉林,落叶松林土壤酶NAG∶AP与其他3个林分间差异显著(P<0.05),其他3个林分间差异不显著(P>0.05)。
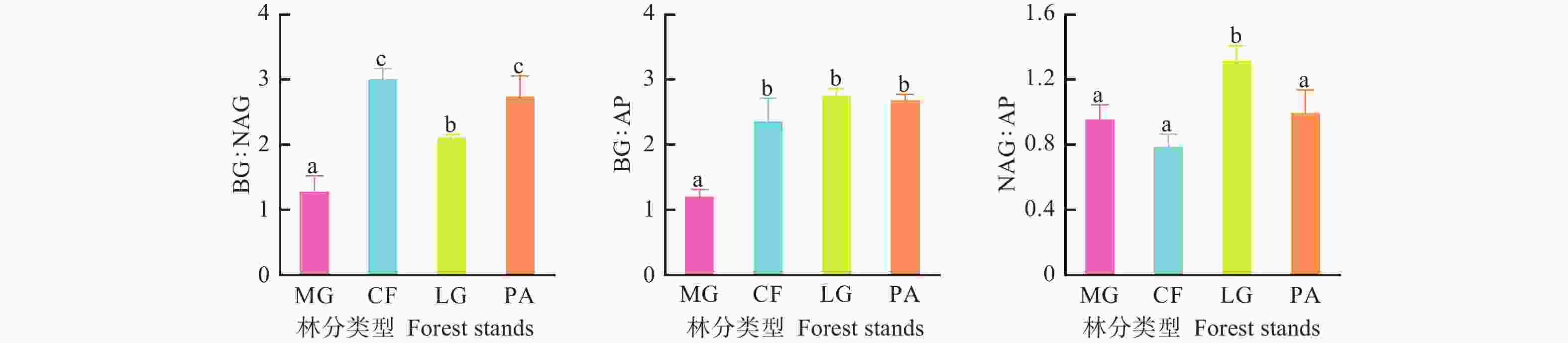
图 2 不同林分土壤酶化学计量比变化特征
Figure 2. The various of soil enzyme stoichiometry in different forest stands
-
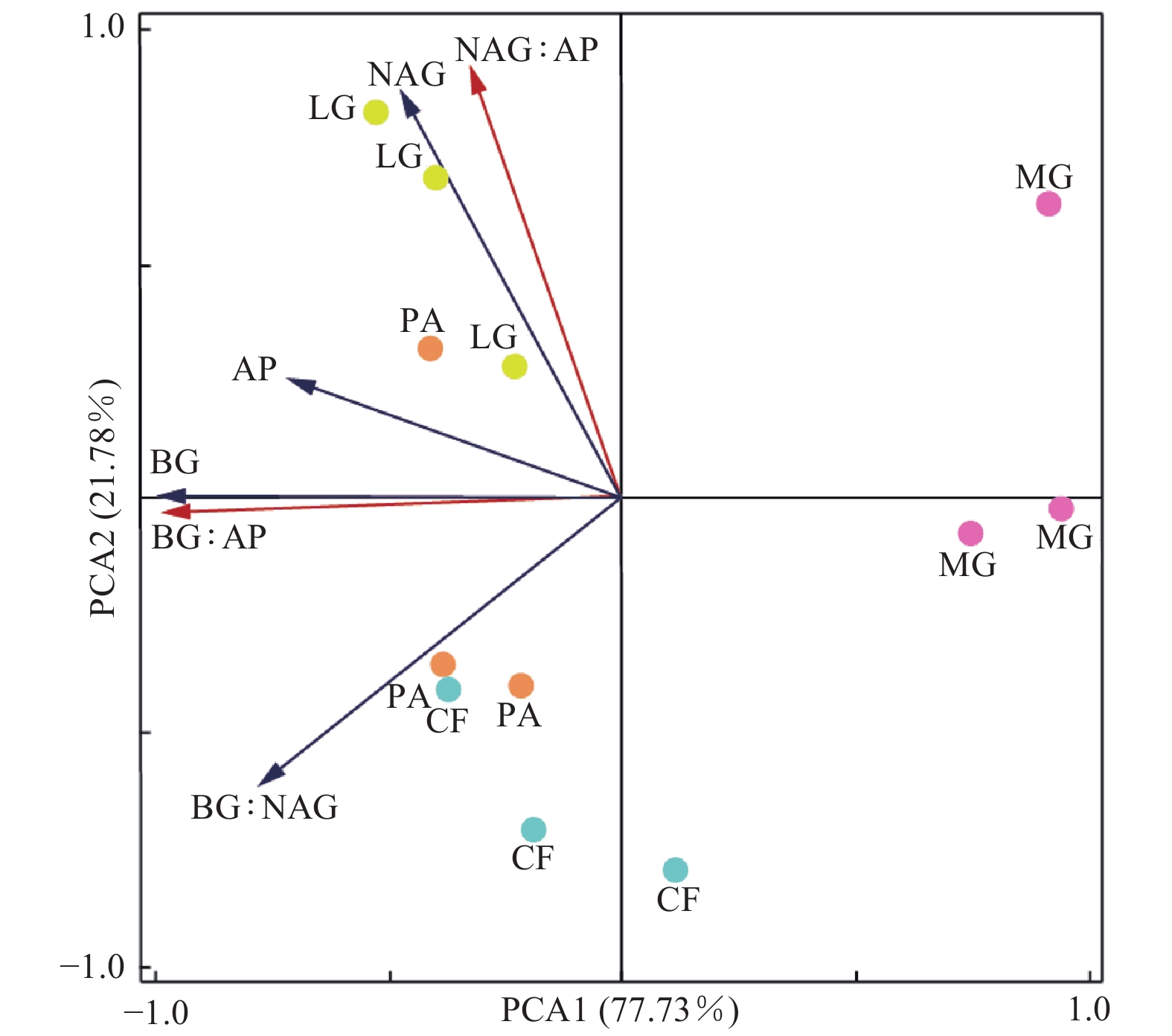
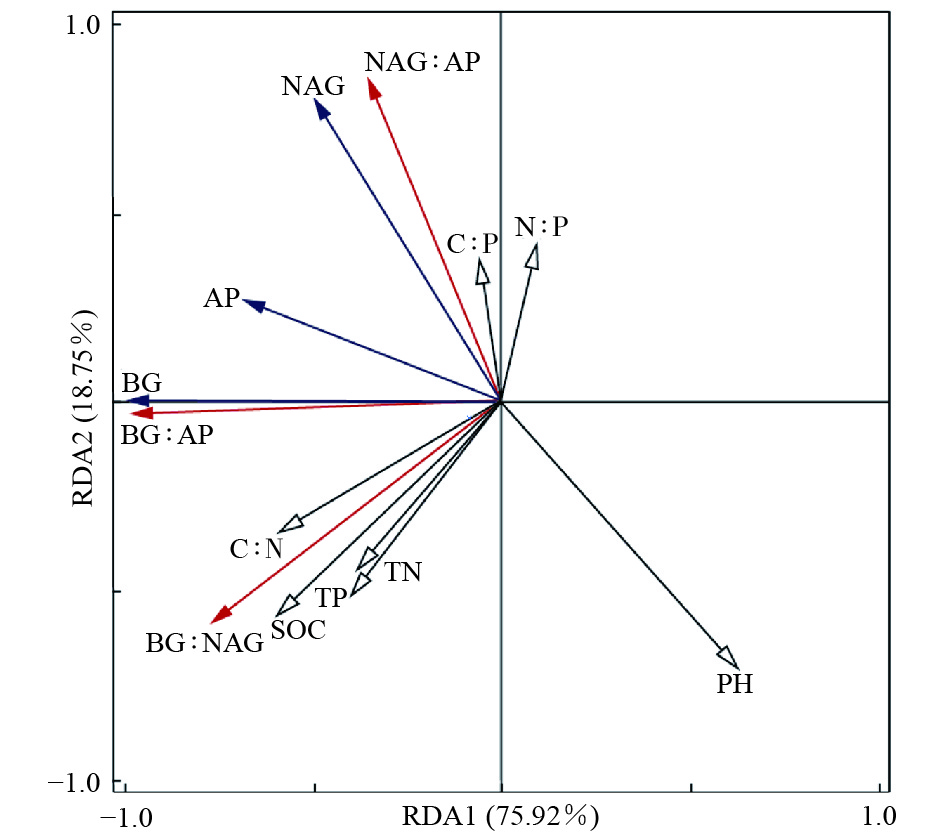
不同林分土壤酶活性及其生态化学计量比的主成分分析结果(图3)表明:PCA1轴可以解释不同林分差异的77.73%,该轴与BG酶(R=−0.99)、AP酶(R=−0.72)、BG∶AP(R=−0.99)和BG∶NAG(R=−0.78)具有较高的相关性;水杉林与其他3个林分沿PCA1具有明显的分异。PCA2轴可以解释不同林分类型之间差异的21.78%,该轴与NAG酶(R=0.88)、BG∶NAG(R=−0.62)和NAG∶AP(R=0.93)具有较高的相关性;落叶松林与柳杉林之间沿PCA2轴具有明显的分异。RDA分析结果(图4)显示:7个土壤化学性质指标共解释了土壤酶活性及计量比变异的94.67%。土壤pH(P=0.01)和SOC(P=0.002)是影响土壤酶活性和酶化学计量比的主要因子,分别解释了土壤酶活性和土壤酶化学计量比40.23%和41.42%的变异。
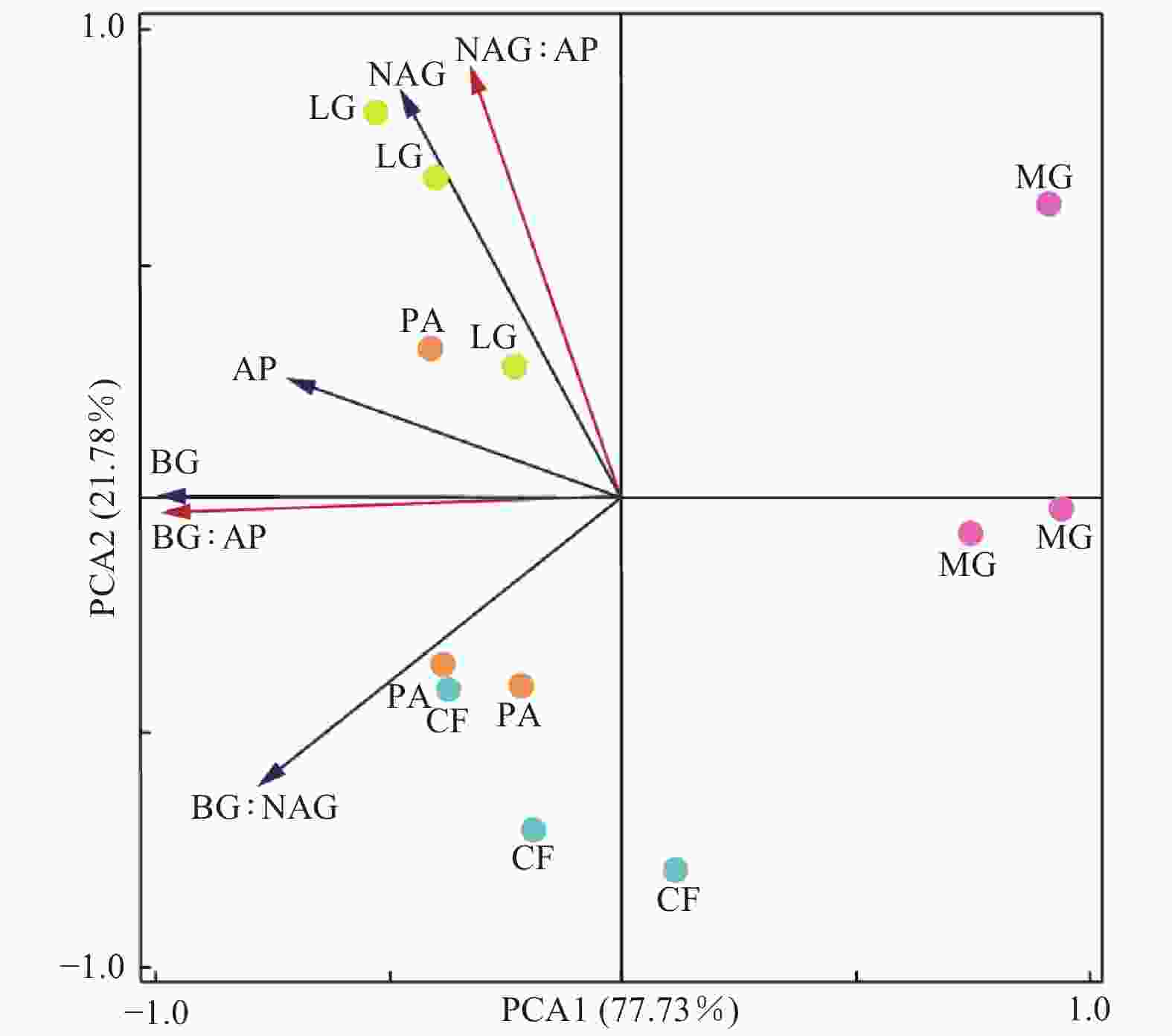
图 3 土壤酶活性及其化学计量比的主成分分析
Figure 3. Principle analysis (PCA) of soil enzyme activity and stoichiometry in different forest stands
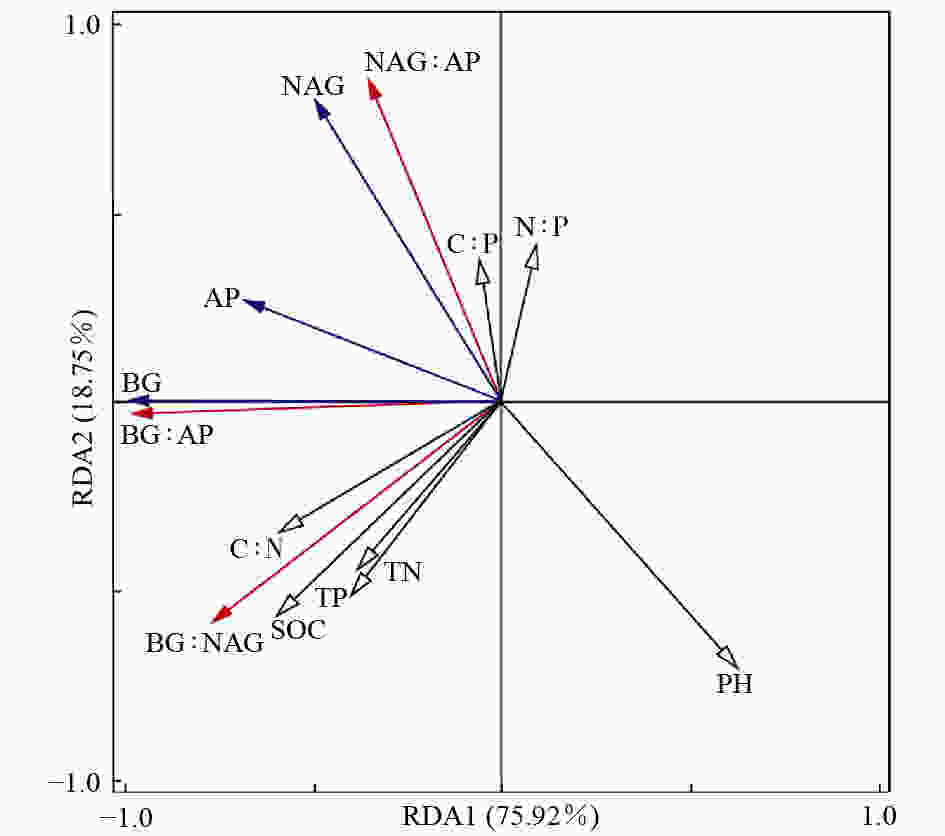
图 4 土壤酶活性及其计量比与土壤化学性质的冗余分析
Figure 4. Redundancy analysis (RDA) of the effect of soil chemical properties on soil enzyme activity and stoichiometry in different forest stands
土壤酶及其化学计量比与土壤化学性质Pearson相关分析(表3)表明:土壤BG和NAG活性与土壤pH显著(P<0.05)或极显著(P<0.01)负相关;土壤BG∶NAG与土壤SOC极显著(P<0.01)正相关,与TN、TP和C∶N显著正相关(P<0.05);土壤BG∶AP与土壤C∶N显著正相关(P<0.05),与土壤pH显著负相关(P<0.05);土壤NAG∶AP与土壤pH极显著负相关(P<0.01)。
表 3 土壤酶及其化学计量比与土壤化学性质相关分析
Table 3. The relationship between soil enzyme activities or stoichiometry and soil chemical properties.
指标 Indicator SOC TN TP C:N C:P N:P pH BG 0.586* 0.363 0.388 0.570 0.071 −0.082 −0.660* NAG −0.121 −0.115 −0.230 −0.023 0.387 0.340 −0.872** AP 0.603* 0.455 0.312 0.240 0.070 0.004 −0.472 BG:NAG 0.814** 0.589* 0.645* 0.648* −0.224 −0.359 −0.009 BG:AP 0.548 0.319 0.389 0.614* 0.054 −0.108 −0.658* NAG:AP −0.313 −0.291 −0.362 −0.097 0.417 0.384 −0.875** 注:*: P <0.05;**: P <0.01. BG:β-葡萄糖甘酶;NAG:β-乙酰葡糖胺糖苷酶;AP:磷酸单酯酶。
Notes:*:P<0.05;**:P<0.01. BG:β-glucosidase;NAG:β-acetylglucosidase;AP:acid phosphatase. -
不同林分之间土壤酶活性存在差异,冗余分析结果表明:土壤酶活性主要与土壤SOC和pH值有关,其中,土壤BG和AP活性与土壤SOC含量呈显著正相关,这与Sinsabaugh等[9]对全球尺度土壤酶活性研究结果和Guo等[26]对中国东南部喀斯特地貌和非喀斯特地貌森林土壤酶活性研究结果一致。土壤BG与pH值呈显著负相关,土壤NAG与pH值呈极显著负相关,这与Xu等[13]对中国东南部南北森林样带土壤酶活性研究结果相反,这可能主要是因为后者为区域尺度,pH值变化范围较大,而本文是局域尺度,pH值变化范围较小。本研究中4个林分处于同一个区域,立地条件基本相似,凋落物和根系可能是不同树种调控土壤养分的主要途径。因此,未来应进一步分析不同树种的凋落物和细根质量及数量等相关指标与土壤酶活性的关系。
柳杉林和华山松林土壤BG活性显著高于水杉林,该研究结果一方面可能与不同树种凋落物输入差异有关,因为柳杉林和华山松林属于常绿针叶林,水杉林和落叶松林属于落叶型针叶林,落叶型针叶林凋落物输入明显高于常绿针叶林,针叶林凋落物中木质素、多酚类等难溶性物质含量较高[27],加之,凋落物长期大量积累,土壤表层透气性差,凋落物更不易分解,所以落叶型针叶林对土壤有机碳分解影响较小,水杉林土壤BG活性较低;另一方面可能与不同植物的根系渗出物速率和成分不同造成根际碳含量的差异有关[19]。另外,落叶松林土壤BG活性与水杉林之间差异显著,可能还与落叶松林凋落物树脂、单宁等酸性物质含量高,易导致土壤酸化有关[28]。本研究中,落叶松林土壤pH值为5.53,显著低于其它3个林分。土壤水解酶活性最适宜的pH值为5左右[9],说明落叶松林土壤偏酸性提高了土壤酶活性。落叶松林土壤NAG活性显著高于其他3个林分,可能也与落叶松林土壤偏酸性有关。不同林分之间土壤AP活性差异不显著,说明土壤酶活性除了受土壤养分和pH值影响外,可能还受微生物群落组成和结构等生物因素影响[9, 29]。
-
土壤BG、NAG和AP活性代表了微生物C、N、P获得能力。柳杉林、华山松林、落叶松林土壤BG∶NAG显著高于水杉林,说明柳杉林、华山松林和落叶松林土壤微生物所需的有效碳源高于水杉林。土壤BG∶NAG与土壤C∶N呈显著正相关,反映了柳杉林、华山松林和落叶松林土壤有机质质量可能低于水杉林。柳杉林、华山松林土壤BG∶NAG较高,说明其土壤有机质质量最低,这与柳杉林、华山松林土壤C/N比较高的结果一致;水杉林土壤BG∶AP也显著低于其他3个林分,且土壤BG∶AP与C∶N呈显著正相关,进一步说明了柳杉林、华山松林和落叶松林土壤有机质质量相对较低;落叶松林土壤NAG∶AP活性显著高于其他林分,这可能主要与落叶松林土壤酸化有关。土壤NAG∶AP与土壤pH值呈极显著负相关,这与张星星等[30]和Xu等[13]研究结果一致。另外,冗余分析得出,土壤SOC是土壤酶活性的主要调控因素。因此,4个林分土壤酶化学计量比可以反映其土壤养分状况。
本文所研究区域土壤BG∶NAG∶AP对数转换后均值为1.00∶0.80∶0.81,同全球尺度上土壤C、N、P酶化学计量比1∶1∶1有所偏离[9],反映了本区域微生物对土壤碳源需求较高。这主要与本研究中4种林分均为针叶纯林有关,因为,针叶林凋落物相比阔叶林凋落物输入量少,且难分解,养分归还缓慢,土壤有机质质量相对不高[17, 27]。本研究区域BG:NAG比值(2.27±0.72)处于热带森林生态系统土壤BG:NAG(1.83±0.31)[12]和全球生态系统土壤BG:NAG(1.81±0.47)范围内,反映了本区域土壤微生物对C和N元素需求基本保持相对平衡的模式。另外,本区域土壤氮磷比值(4∶1)低于全球尺度氮磷比值(13∶1)[11]和中国区域尺度森林土壤氮磷比值(14∶1),说明本区域土壤微生物可能受到氮元素养分的限制[13],该结果与以往报道的全球和我国热带及亚热带地区森林生态系统土壤微生物存在磷元素养分限制的结果不同[12-13],而与Guo等[26]对中国西南地区喀斯特地貌森林生态系统研究结果一致。本研究认为,相比全球和区域尺度,在局部区域尺度上,林龄、林分类型、土壤母质类型等因素均会造成土壤养分变化[30-31],进而影响微生物对酶活性的表达,如张星星等[30]对中亚热带不同森林类型研究发现,不同母质类型下天然林和杉木人工林土壤BG∶AP和NAG∶AP均存在显著差异。
-
本研究区域神农架4种针叶人工林之间土壤酶活性及其生态化学计量比存在差异;林分对土壤酶活性及其生态化学计量比的影响主要是通过其对土壤有机碳和pH值调控来实现的;本研究区域人工林土壤微生物可能受到土壤氮限制。因此,未来对本区域人工林土壤养分管理应考虑微生物的氮素需求。
神农架4种典型针叶人工林土壤酶活性及其生态化学计量特征
Soil Enzyme Activity and Its Ecological Stoichiometry in Four Typical Coniferous Planted Forests in Shennongjia National Nature Reserve, China
-
摘要:
目的 探讨土壤酶活性及其生态化学计量比对土壤微生物养分的限制和土壤养分有效性的指示作用。 方法 以神农架地区水杉、柳杉、落叶松和华山松4种人工林纯林土壤为研究对象,测定不同林分土壤酶活性及其生态化学计量比和土壤化学性质。 结果 柳杉林、华山松林和落叶松林土壤β-葡萄糖苷酶(BG)活性显著高于水杉林,落叶松林土壤β-乙酰葡糖胺糖苷酶(NAG)活性显著高于其他3个林分,不同林分间土壤磷酸单酯酶(AP)差异不显著;水杉林土壤BG∶NAG和BG∶AP显著低于其他3个林分,落叶松林土壤BG∶NAG显著低于柳杉林和华山松林,落叶松林土壤NAG∶AP显著高于其他3个林分,其中,本区域4种林分土壤NAG∶AP(1.01±0.22)指示4种类型人工林土壤微生物可能受到土壤氮限制。冗余分析显示:土壤有机碳和pH值是土壤酶活性的重要影响因素。 结论 林分是土壤酶活性及其生态化学计量比的重要影响因素,其对土壤有机碳和pH值的影响是调控土壤酶活性变化的主要因素。 Abstract:Object To study the role of soil enzyme activity and its ecological stoichiometry in limiting soil microbial nutrients and indicating soil nutrient availability. Method Soil enzyme activity and its ecological stoichiometry, along with soil physical and chemical properties, were measured in Metasequoia glyptostroboides, Cryptomeria japonica var. sinensis, Larix gmelinii and Pinus armandii forests in Shennongjia National Nature Reserve. Result The soil β-glucosidase (BG) activity in C. japonica, P. armandii and L. gmelinii forests was significantly higher than that in M. glyptostroboides forest. The soil β-acetylglucosidase (NAG) activity in L. gmelinii forest was significantly higher than that in other forests. There was no significant difference in soil acid phosphatase (AP) activity among the four types of forest stands. Soil BG∶NAG and BG∶AP in M. glyptostroboides forest were significantly lower than that in other forests. Soil BG∶NAG in L. gmelinii forest was also significantly lower than that in C. japonica and P. armandii forests. Soil NAG∶AP in L. gmelinii forest was significantly higher than that in other forests. Soil NAG∶AP (1.01±0.22) in the region indicated the soil microbes were limited by soil nitrogen. Redundancy analysis showed that soil organic carbon and pH value were important factors influencing soil enzyme activity. Conclusion Forest stand is an important factor influencing soil enzyme activity and the ecological stoichiometry of soil enzyme. The effect on soil organic C and pH is the key factor regulating soil enzyme activity. -
图 1 不同林分土壤酶活性变化特征
Figure 1. The various of soil enzyme activity in different forest stands
图 2 不同林分土壤酶化学计量比变化特征
Figure 2. The various of soil enzyme stoichiometry in different forest stands
图 3 土壤酶活性及其化学计量比的主成分分析
Figure 3. Principle analysis (PCA) of soil enzyme activity and stoichiometry in different forest stands
图 4 土壤酶活性及其计量比与土壤化学性质的冗余分析
Figure 4. Redundancy analysis (RDA) of the effect of soil chemical properties on soil enzyme activity and stoichiometry in different forest stands
表 1 4种林分的基本概况
Table 1. Basic characteristics of four types of forest stands
林分类型
Forest type海拔
Altitude/m坡度
Slope/(°)林分密度
Stand density/(株·hm−2)平均树高
Mean height of tree/m平均胸径
Mean diameter/cmMG 1 650 21~27 875±164 19.5 21.74 CF 1 640 9~15 792±206 24.5 20.45 LG 1 790 9~15 843±182 22.4 21.70 PA 1 580 3~7 956±139 12.0 14.68 注:MG: 水杉林;CF: 柳杉林;LG:落叶松林;PA:华山松林。下同
Notes:MG: Metasequoia glyptostroboides forest;CF:Cryptomeria japonica forest;LG:Larix gmelinii forest;PA:华山松林Pinus armandii forest.the same below. 下载: 导出CSV
下载: 导出CSV
表 2 4种林分类型土壤化学性质
Table 2. Soil chemical properties of four types of forest stands
林分类型
Forest type有机碳
SOC/(g·kg−1)全氮
TN/(g·kg−1)全磷
TP/(g·kg−1)碳氮比
C∶N碳磷比
C∶P氮磷比
N∶PpH值 MG 32.08±0.70 a 2.54±0.01 a 0.61±0.01 a 12.61±0.30 a 52.23±1.80 b 4.14±0.05 b 7.10±0.22 c CF 46.91±0.70 c 3.50±0.02 b 0.86±0.01 b 13.42±0.28 b 54.33±0.40 b 4.05±0.09 b 7.13±0.12 c LG 37.00±6.88 ab 2.82±0.53 a 0.57±0.16 a 13.12±0.04 b 66.38±8.31 c 5.06±0.62 c 5.53±0.05 a PA 40.89±0.70 bc 2.70±0.02 a 1.06±0.12 c 15.13±0.05 c 38.98±4.65 a 2.58±0.30 a 6.34±0.07 b 注:不同小写字母代表不同林分类型之间差异性显著(P<0.05),下同。
Notes: Different lowercase letters indicate significant difference among different forest stands (P<0.05). SOC:soil organic carbon;TN: total nitrogen;TP:total phosphorus;C:N:the ratio of carbon and nitrogen;C:P:the ratio of carbon and phosphorus;N:P:the ratio of nitrogen and phosphorus.the same below.
下载: 导出CSV
表 3 土壤酶及其化学计量比与土壤化学性质相关分析
Table 3. The relationship between soil enzyme activities or stoichiometry and soil chemical properties.
指标 Indicator SOC TN TP C:N C:P N:P pH BG 0.586* 0.363 0.388 0.570 0.071 −0.082 −0.660* NAG −0.121 −0.115 −0.230 −0.023 0.387 0.340 −0.872** AP 0.603* 0.455 0.312 0.240 0.070 0.004 −0.472 BG:NAG 0.814** 0.589* 0.645* 0.648* −0.224 −0.359 −0.009 BG:AP 0.548 0.319 0.389 0.614* 0.054 −0.108 −0.658* NAG:AP −0.313 −0.291 −0.362 −0.097 0.417 0.384 −0.875** 注:*: P <0.05;**: P <0.01. BG:β-葡萄糖甘酶;NAG:β-乙酰葡糖胺糖苷酶;AP:磷酸单酯酶。
Notes:*:P<0.05;**:P<0.01. BG:β-glucosidase;NAG:β-acetylglucosidase;AP:acid phosphatase.
下载: 导出CSV
-
[1] Luo L, Meng H, Gu J D. Microbial extracellular enzymes in biogeochemical cycling of ecosystems[J]. Journal of Environmental Management, 2017, 197: 539-549. doi: 10.1016/j.jenvman.2017.04.023 [2] Sinsabaugh R L, Belnap J, Findlay S G,<italic> et al</italic>. Extracellular enzyme kinetics scale with resource availability[J]. Biogeochemistry, 2014, 121(2): 287-304. doi: 10.1007/s10533-014-0030-y [3] 许淼平, 任成杰, 张 伟, 等. 土壤微生物生物量碳氮磷与土壤酶化学计量对气候变化的响应机制[J]. 应用生态学报, 2018, 29(7):2445-2454. [4] Allison V J, Condron L M, Peltzer D A,<italic> et al</italic>. Changes in enzyme activities and soil microbial community composition along carbon and nutrient gradients at the Franz Josef chronosequence, New Zealand[J]. Soil Biology and Biochemistry, 2007, 39(7): 1770-1781. doi: 10.1016/j.soilbio.2007.02.006 [5] 丁怡飞, 曹永庆, 姚小华, 等. 鼠茅草间作对油茶林地土壤养分及酶活性的影响[J]. 林业科学研究, 2018, 31(2):170-175. [6] 赵睿宇, 李正才, 王 斌, 等. 毛竹林地覆盖和翻耕对土壤酶活性及土壤养分含量的影响[J]. 林业科学研究, 2019, 32(5):67-73. [7] Hill B H, Elonen C M, Jicha T M,<italic> et al</italic>. Sediment microbial enzyme activity as an indicator of nutrient limitation in Great Lakes coastal wetlands[J]. Freshwater Biology, 2006, 51(9): 1670-1683. doi: 10.1111/j.1365-2427.2006.01606.x [8] Schimel J P, Weintraub M N. The implications of exoenzyme activity on microbial carbon and nitrogen limitation in soil: a theoretical model[J]. Soil Biology and Biochemistry, 2003, 35(4): 549-563. doi: 10.1016/S0038-0717(03)00015-4 [9] Sinsabaugh R L, Lauber C L, Weintraub M N,<italic> et al</italic>. Stoichiometry of soil enzyme activity at global scale[J]. Ecology Letters, 2008, 11(11): 1252-1264. doi: 10.1111/j.1461-0248.2008.01245.x [10] Fanin N, Moorhead D, Bertrand I. Eco-enzymatic stoichiometry and enzymatic vectors reveal differential C, N, P dynamics in decaying litter along a land-use gradient[J]. Biogeochemistry, 2016, 129(1-2): 21-36. doi: 10.1007/s10533-016-0217-5 [11] Sinsabaugh R L, Hill B H, Shah J J F. Ecoenzymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment[J]. Nature, 2009, 462(7274): 795-798. doi: 10.1038/nature08632 [12] Waring B G, Weintraub S R, Sinsabaugh R L. Ecoenzymatic stoichiometry of microbial nutrient acquisition in tropical soils[J]. Biogeochemistry, 2014, 117(1): 101-113. doi: 10.1007/s10533-013-9849-x [13] Xu Z, Yu G, Zhang X,<italic> et al</italic>. Soil enzyme activity and stoichiometry in forest ecosystems along the North-South Transect in eastern China (NSTEC)[J]. Soil Biology and Biochemistry, 2017, 104: 152-163. doi: 10.1016/j.soilbio.2016.10.020 [14] 薛 立, 邝立刚, 陈红跃, 等. 不同林分土壤养分、微生物与酶活性的研究[J]. 土壤学报, 2003, 40(2):280-285. doi: 10.3321/j.issn:0564-3929.2003.02.018 [15] 张雅茜, 方 晰, 冼应男, 等. 亚热带区4种林地土壤微生物生物量碳氮磷及酶活性特征[J]. 生态学报, 2019, 39(14):5326-5338. [16] 李春萍, 王世伟, 丁俊杰, 等. 施氮水平对核桃细根呼吸速率及相关酶活性的影响[J]. 林业科学研究, 2019, 32(6):56-62. [17] 左 巍, 贺康宁, 田 赟, 等. 青海高寒区不同林分类型凋落物养分状况及化学计量特征[J]. 生态学杂志, 2016, 35(9):2271-2278. [18] 弓文艳, 陈丽华, 郑学良. 基于不同林分类型下土壤碳氮储量垂直分布[J]. 水土保持学报, 2019, 33(1):152-157, 164. [19] Yin H J, Wheeler E, Phillips R P. Root-induced changes in nutrient cycling in forests depend on exudation rates[J]. Soil Biology and Biochemistry, 2014, 78: 213-221. doi: 10.1016/j.soilbio.2014.07.022 [20] 袁传武, 余崇彪, 石冰天, 等. 神农架林区森林资源可持续发展途径探讨[J]. 湖北林业科技, 2007(5):47-50. doi: 10.3969/j.issn.1004-3020.2007.05.014 [21] 周志文, 潘剑君, 居为民, 等. 神农架不同坡位3种林型土壤碳氮比分布特征[J]. 水土保持学报, 2014, 28(4):210-217. [22] 崔鸿侠, 肖文发, 黄志霖, 等. 神农架3种针叶林土壤碳储量比较[J]. 东北林业大学学报, 2014, 42(3):69-72. doi: 10.3969/j.issn.1000-5382.2014.03.016 [23] 谢宗强, 申国珍, 周友兵, 等. 神农架世界自然遗产地的全球突出普遍价值及其保护[J]. 生物多样性, 2017, 25(5):490-497. doi: 10.17520/biods.2016268 [24] 关松荫. 土壤酶及其研究法[J]. 北京: 农业出版社, 1986. [25] Turner B L, Wright S J. The response of microbial biomass and hydrolytic enzymes to a decade of nitrogen, phosphorus, and potassium addition in a lowland tropical rain forest[J]. Biogeochemistry, 2014, 117(1): 115-130. doi: 10.1007/s10533-013-9848-y [26] Guo Z, Zhang X, Green S M,<italic> et al</italic>. Soil enzyme activity and stoichiometry along a gradient of vegetation restoration at the Karst Critical Zone Observatory in Southwest China[J]. Land Degradation and Development, 2019, 30(16): 1916-1927. doi: 10.1002/ldr.3389 [27] 曾曙才, 苏志尧, 古炎坤, 等. 广州白云山风景名胜区主要林分类型凋落物的研究[J]. 应用生态学报, 2003, 14(1):154-156. doi: 10.3321/j.issn:1001-9332.2003.01.035 [28] 吉艳芝, 冯万忠, 陈立新, 等. 落叶松混交林根际与非根际土壤养分、微生物和酶活性特征[J]. 生态环境, 2008, 17(1):339-343. [29] 赵京京, 王超群, 董玉红, 等. 细菌肥料对湿地松幼龄林生长及土壤性质的影响[J]. 林业科学研究, 2019, 32(1):153-159. [30] 张星星, 杨柳明, 陈 忠, 等. 中亚热带不同母质和森林类型土壤生态酶化学计量特征[J]. 生态学报, 2018, 38(16):5828-5836. [31] 乔 航, 莫小勤, 罗艳华, 等. 不同林龄油茶人工林土壤酶化学计量及其影响因素[J]. 生态学报, 2019, 39(6):1887-1896. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 4514
- HTML全文浏览量: 2190
- PDF下载量: 99
- 被引次数: 0
