


-
竹卵圆蝽(Hippotiscus dorsalis (Stål))是广泛分布于我国各竹产区的一种重要害虫。竹卵圆蝽的若虫和成虫在竹秆和竹枝的节上吸食汁液,致使被害枝条落叶枯死,严重者可致整株竹子死亡[1-2]。据2015年统计,浙江省安吉县毛竹林竹卵圆蝽的有虫株率在56.78%以上,单株最高虫口达到254头,且近几年来其危害呈上升的趋势[3]。因此,竹卵圆蝽仍是我国部分竹产区高危险性的害虫。
依据竹卵圆蝽群聚吸汁和越冬若虫上竹危害的生物学特性,我国学者提出了人工捕捉、粘虫胶带粘虫、竹腔注射化学农药、喷撒化学农药等防治方法[4]。这些方法在短期内取得较好的防治效果,但也存在很多不足。竹卵圆蝽的臭腺能分泌防御性化学物质,这些化学物质可引起人的皮肤发黄、起泡、溃疡等症状,故人工捕捉竹卵圆蝽时常常会伤及作业人员。化学农药的不合理使用,易产生害虫抗药性增强、次要害虫爆发、环境污染、人畜中毒、林产品品质下降等风险[5-8]。
生物防治具有靶标性强、对人畜安全、环境兼容性好等优点,是害虫绿色防控的重要措施。通过对竹卵圆蝽天敌昆虫的调查,已经找到数种寄生性和捕食性的天敌昆虫[2, 9-10],但这些天敌昆虫在自然界中的数量有限,人工繁育技术不成熟,难以在生产中大规模应用。昆虫病原真菌因其易于室内大规模繁育、储存和运输,施用技术简单,而成为最具开发潜力和应用前景的生物防治制剂之一[7, 11]。本研究通过组织分离法,从罹病的竹卵圆蝽体内分离病原真菌,通过形态学、DNA条形码相结合的方法对病原真菌进行物种鉴定,采用生物测定法评价不同菌株的毒力,筛选出对竹卵圆蝽若虫具有高毒力的菌株,研究结果为竹卵圆蝽的生物防治提供科学依据。
-
作者于2017年在浙江省杭州市富阳区大源镇毛竹(Phyllostachys edulis (Carrière) J. Houz.)竹秆上发现有死亡的竹卵圆蝽,且在死亡虫体表面长出白色的菌丝,初步确定其被病原真菌侵染。将死亡虫体用镊子从竹秆上取下,置于无菌的离心管中,每管1头,带回实验室用于病原真菌的分离。
从离心管中取出虫体,放在流水下冲洗10 min,依次使用75%乙醇洗涤30 s,10%次氯酸钠洗涤10 min,无菌水洗涤5次。洗涤后的虫体转移到无菌滤纸上,吸干表面残留的水分,转接到含链霉素(200 mg·L−1)的PDA固体培养基上,25 ℃暗培养,观察病原真菌在PDA培养基上的培养性状和生长状况。
-
观察并描述病原真菌在PDA固体培养基上形成菌落的形态特征。在VHX5000显微镜(KEYENCE公司,日本)下观察病原真菌的菌丝和分生孢子,使用显微镜的测量软件VHX-H2M2测定分生孢子大小。参考Zimmermann对昆虫病原真菌的形态特征和培养性状的描述[12],对分离病原真菌进行形态学鉴定。
采用DNA条形码的方法对病原真菌进行分子鉴定[13]。用无菌吸头从平板上刮取菌丝和分生孢子,置于2 mL的无菌离心管中,在Tissue lyser-48研磨仪上研磨粉碎,具体研磨步骤参考耿显胜等的方法[14]。采用引物对ITS1/ITS4进行PCR扩增[15-16],PCR扩增的反应体系为:2 × PCR Mix 10 μL,10 μmol·L−1的正、反向引物各0.5 μL,DNA模板1 μL,加ddH2O补足20 μL。PCR扩增的反应条件为:94 ℃预变性5 min;94 ℃变性35 s,57 ℃退火35 s,72 ℃延伸40 s,共35个循环;最后72 ℃延伸10 min。PCR产物经凝胶电泳分析,阳性的PCR扩增产物送铂尚生物技术有限公司测序。测序序列使用DNAMAN 6.0进行多序列比对分析,使用NCBI(National Center for Biotechnology Information,NCBI)和BOLD(The Barcode of Life Data System,BOLD)数据库进行在线比对分析。
-
采用浸渍法测定分离菌株的毒力。选取生长健壮且无杂菌污染的4株昆虫病原真菌LYC4、LYC10、LYC11和LYC15转接到PDA固体培养基上,25 ℃下暗培养15 d,让其产生足够数量的分生孢子。刮取分生孢子,置于含0.05% Tween-80的无菌水中,使用无菌玻棒将刮取的孢子块分散成单个的分生孢子,使用金属过滤器过滤除去菌丝和未分散开的孢子块。采用血球细胞计数法测定孢子的浓度,用含0.05% Tween-80的无菌水将分生孢子调成1 × 107和1 × 108孢子·mL−1的悬浮液。将竹卵圆蝽3龄若虫置于分生孢子悬浮液中浸泡2 s,然后转移到含湿润滤纸的无菌培养皿(直径9 cm)中,并在培养皿中加入一段新鲜的毛竹竹秆,用于饲喂竹卵圆蝽若虫。每皿转接竹卵圆蝽若虫6头,每个处理为4皿。试验以含0.05% Tween-80的无菌水浸泡的竹卵圆蝽3龄若虫为对照处理组。接虫后的培养皿置于人工气候箱内培养13 d,培养条件为温度25 ℃,光照强度2 000 lx,光周期10L:14D。
接虫后每天观察竹卵圆蝽的活动情况,统计死亡虫体的数量,计算逐日的累计死亡率和校正累计死亡率:
$ 累计死亡率\text=\frac{累计死亡虫数}{试虫总数}\times 100\text{%} $
$\begin{array}{c} 校正累计死亡率\text=\\ \dfrac{处理组累计死亡率-对照组累计死亡率}{1-对照组累计死亡率}\times 100\text{%} \end{array} $
将校正累计死亡率转换成死亡率概率值,统计的天数转换成以10为底的对数,采用SPSS24.0软件进行线性回归分析,获取回归方程和决定系数,通过回归方程计算出致死中时间LT50值。采用SigmaPlot 12.5软件制作校正累计死亡率的散点图。
-
室内筛选出的高毒力菌株LYC11转接到PDA固体培养基上,25 ℃下暗培养15 d。采用血球细胞计数法将分生孢子使用含0.05% Tween-80的无菌水调制成5 × 107、1 × 107、 1 × 106、1 × 105和1 × 104孢子·mL−1的悬浮液,用含0.05% Tween-80的无菌水作为对照处理,每个处理重复3次。
在浙江省安吉县成片毛竹林开展生物测定。安吉县位于浙江省西北部,处于北亚热带季风气候区。2019年4月,该县天气以阴雨天为主,月平均最高气温23 ℃,月平均最低气温12 ℃。在毛竹林捕捉聚集于竹秆上的竹卵圆蝽若虫,置于预先配制好的分生孢子悬浮液中浸泡5 s,取出后接于袖笼内,袖笼套在毛竹竹秆上。接虫后第18 天,统计袖笼内竹卵圆蝽的总数和死亡虫体数量,计算累计死亡率。采用SPSS24.0软件的Probit回归法获取回归方程和致死中浓度LC50值。
-
本研究使用16头竹卵圆蝽僵虫进行分离培养,共得到7株昆虫病原真菌,7株昆虫病原真菌的编号分别为LYC4、LYC5、LYC6、LYC8、LYC10、LYC11和LYC15。观察分离菌株在PDA固体培养基上的培养性状,该病原真菌在培养基的接种点处长出菌落,菌落包围接种的僵虫,初期菌落为白色绒毛状,后期菌落呈淡黄色粉状。将菌落置于显微镜下观察,其菌丝体白色,分生孢子球形,直径2.93±0.35 μm(图1B),分生孢子常数百个聚集成致密的球状体,球状体白色,直径34.13±5.06 μm(图1A)。依据该菌的形态特征和在PDA固体培养基上的培养性状,参考Zimmermann对球孢白僵菌(Beauveria bassiana (Balsamo) Vuillemin)的描述[12],初步鉴定为球孢白僵菌。

图 1 竹卵圆蝽僵虫分离的球孢白僵菌形态特征
Figure 1. The morphological characteristics of B. bassiana isolated from H. dorsalis
-
采用引物对ITS1/ITS4进行PCR扩增, 7个样品都能够扩增出约600 bp的特异性条带(图2),条带的大小与预期一致。PCR扩增的产物测序和序列分析,得到7个菌株530 bp的ITS rDNA序列。多序列比对表明,7个菌株ITS rDNA序列仅有1个核苷酸位点的差异,同源性为99.97%。ITS rDNA序列提交到DNA条形码鉴定数据库BOLD,进行DNA条形码鉴定,所有7株昆虫病原真菌均鉴定为球孢白僵菌。ITS rDNA序列同时提交到NCBI数据库在线比对,结果表明,LYC4、LYC6、LYC8、LYC10、LYC11、LYC15和LYC5菌株与NCBI数据库中球孢白僵菌(GenBank登录号:MN122432、MK229193、MG345086、MK490863、MH367274、MN401662)ITS rDNA序列的同源性为100.00%。
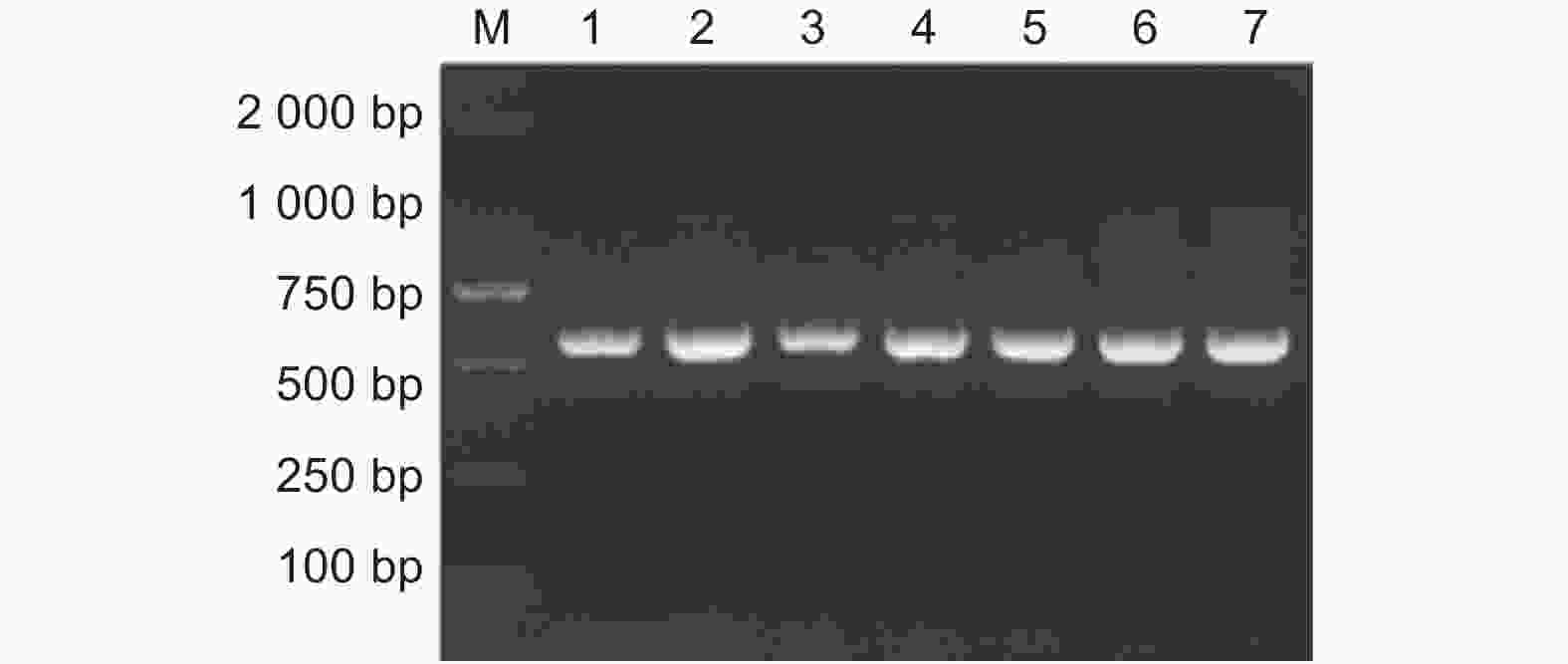
图 2 球孢白僵菌 ITS rDNA 序列的 PCR 扩增
Figure 2. PCR products of ITS rDNA fragment from B. bassiana.
-
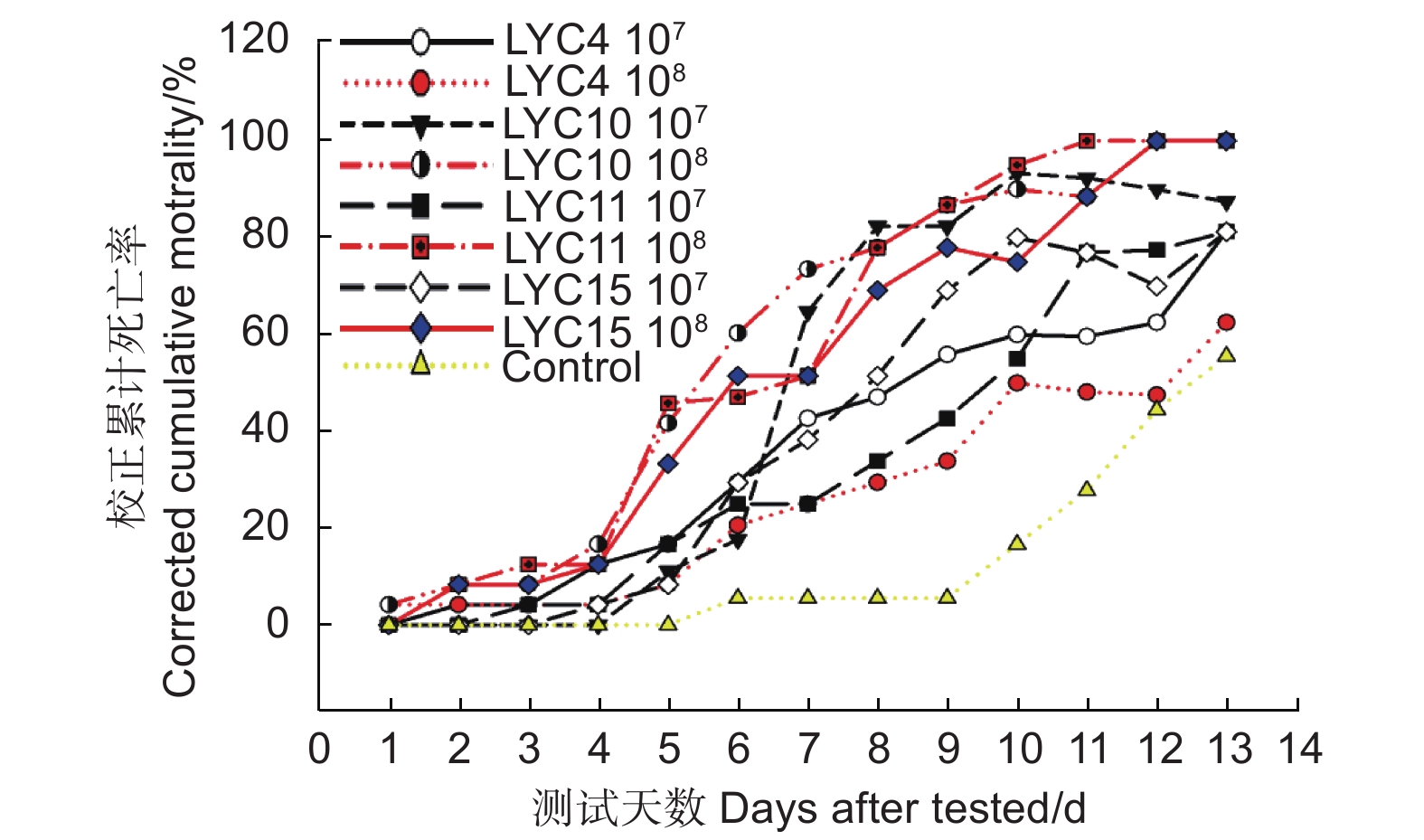
本研究使用4株球孢白僵菌的2种不同浓度的分生孢子悬浮液进行室内毒力测定。研究表明:被球孢白僵菌侵染的竹卵圆蝽若虫的活动性降低,取食减少,并逐渐死亡。虫体死亡后,首先在足部以及躯干的体节处长出白色的菌丝,后期整个虫体被球孢白僵菌包裹。通过连续13 d观察和记录累计死亡率,所有4个菌株对竹卵圆蝽若虫都具有很高的致病性。从校正累计死亡率曲线图上看出:最初的1~4 d,4个菌株对竹卵圆蝽若虫的致病力均较低,从第5天开始校正累计死亡率迅速上升,LYC11菌株在第11 天时,校正累计死亡率达到100.00%,而LYC10和LYC15菌株均在第12 天时,校正累计死亡率达到100.00%(图3)。研究还发现,LYC10、LYC11和LYC15菌株的分生孢子浓度为108孢子·mL−1时,其校正累计死亡率高于分生孢子浓度为107孢子·mL−1的值。
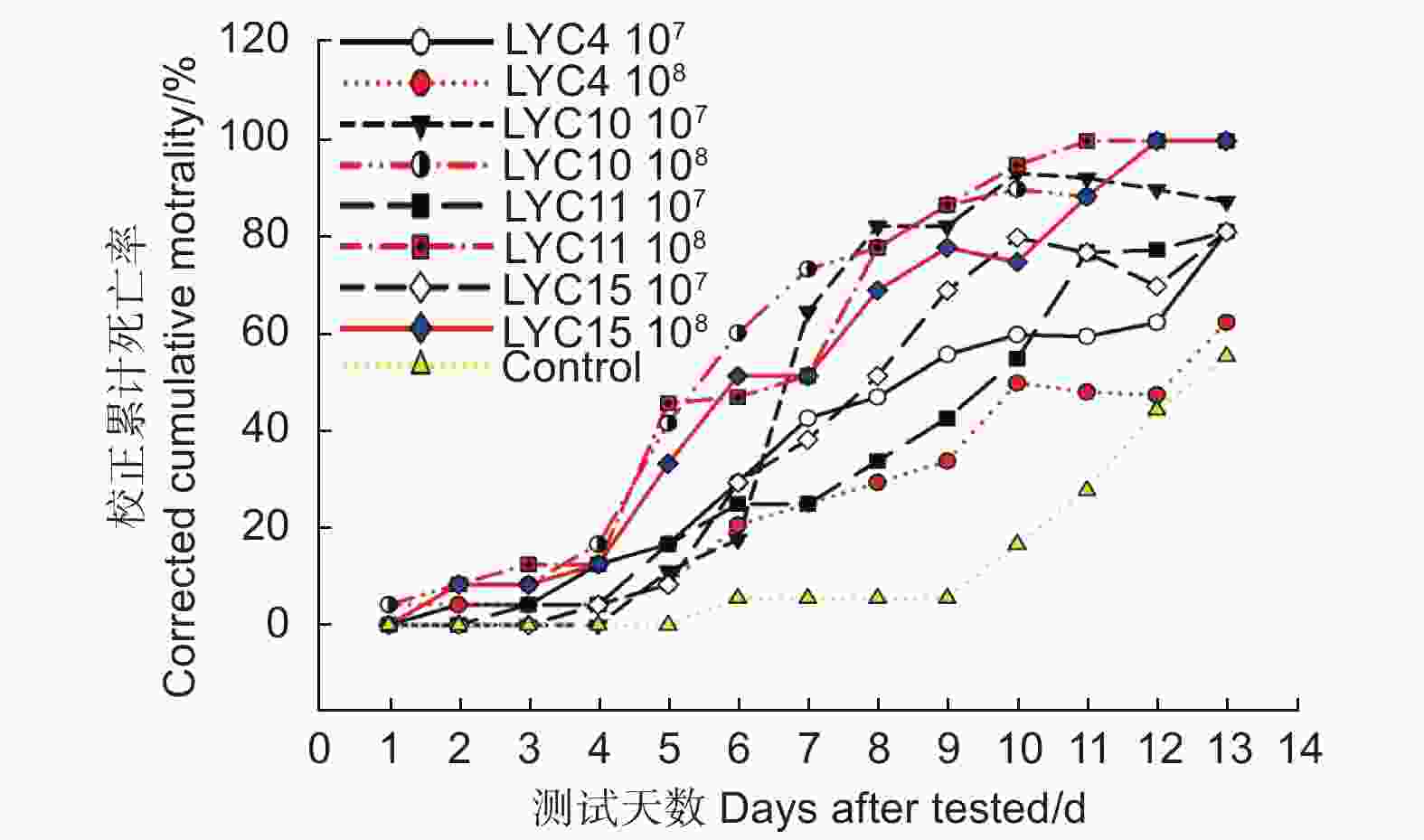
图 3 竹卵圆蝽若虫接种不同浓度球孢白僵菌后的逐日校正累计死亡率
Figure 3. Corrected cumulative mortality of H. dorsalis over time after inoculation with various
采用SPSS24.0进行线性回归分析,获取回归方程和决定系数,依据回归方程计算LT50。所有处理的决定系数R2都在0.800以上,死亡率和时间之间存在显著的线性关系(表1)。4个菌株对竹卵圆蝽若虫的致死中时间LT50在5.04~14.03之间。当分生孢子浓度为108孢子·mL−1时,LYC10、LYC11、LYC15和LYC4菌株的致死中时间LT50分别为5.04 、5.45 、6.17 和14.03 d(表1)。
表 1 球孢白僵菌菌株对竹卵圆蝽若虫的致病性和LT50
Table 1. Pathogenicity and LT50 of the tested B. bassiana strains against H. dorsalis larvae
菌株
Strains浓度
Concentrations/
(分生孢子conidia·mL−1)回归方程
Regression
equation决定系数R2
Correlation
coefficient R2LT50/d 累计死亡率
Cumulative mortality/%校正累计死亡率
Corrected cumulative
mortality/%LYC4 1 × 107 y=3.255x + 1.967 0.954 8.55 91.667 81.251 1 × 108 y=2.06x + 2.637 0.802 14.03 83.333 62.499 LYC10 1 × 107 y=8.611x−2.221 0.927 6.90 94.444 87.499 1 × 108 y=3.351x + 2.647 0.892 5.04 100.000 100.000 LYC11 1 × 107 y=4.38x + 0.869 0.942 8.77 91.667 81.251 1 × 108 y=4.295x + 1.838 0.878 5.45 100.000 100.000 LYC15 1 × 107 y=5.338x + 0.145 0.937 8.12 91.667 81.251 1 × 108 y=3.756x + 2.032 0.925 6.17 100.000 100.000 校正累计死亡率越高,毒力越强;致死中时间LT50越小,毒力越强。综合以上研究结果,确定4株菌株对竹卵圆蝽若虫的毒力从高到低依次为:LYC10>LYC11>LYC15>LYC4。
-
林间生物测定表明:LYC11菌株对竹卵圆蝽若虫具有很高的防治效果,随着分生孢子浓度升高,累计死亡率快速升高。在接菌后第18天,5×107、1×107、1×106、1×105、1×104孢子·mL−1 5个浓度的分生孢子的累计死亡率分别为79.80%、78.89%、77.99%、54.12%、57.14%。
Probit回归分析表明:LYC11菌株的累计死亡率与分生孢子浓度之间的回归方程为y=0.234x−1.025;Pearson拟合优度检验结果为χ2=3.745,P=0.290>0.050,差异不显著,模型拟合良好。依据置信限度表得出LYC11菌株对竹卵圆蝽若虫的致死中浓度LC50为2.43 × 104孢子·mL−1。
-
昆虫病原真菌是一类广泛存在于自然界的昆虫种群抑制因子,能够引起昆虫疾病的流行,且具有较高的选择性、害虫对其抗性发展较慢、可通过寄主传递和扩散等优良特性,而广泛用于害虫的生物防治[17-19]。目前,已经发现约100属700多种昆虫病原真菌,这些真菌中的绝大多数属于子囊菌门(Ascomycota)和 虫霉门(Entomophthoromycota)[19-20]。 球孢白僵菌属于子囊菌门真菌,也是一种兼性营养的昆虫病原真菌,广泛存在于温带和热带地区的受感染昆虫虫体内[12,21]。本研究从毛竹林的竹卵圆蝽僵虫上分离到7株病原真菌,经PDA固体培养基培养和形态特征观察,分离的菌株与Zimmermann描述的球孢白僵菌的培养性状和形态特征一致[12]。提取分离菌株的总DNA,采用DNA条形码法进行物种鉴定,7个菌株均被鉴定为球孢白僵菌。
尽管球孢白僵菌能够侵染15个目的700多种昆虫,然而球孢白僵菌种内存在广泛的自然变异,不同菌株的寄主范围和毒力存在差异,针对靶标昆虫而筛选出具有更强毒力的菌株,是利用球孢白僵菌进行生物防治的关键[12,22-23]。本研究采用室内生物测定法比较了4株球孢白僵菌的毒力,4个菌株的致死中时间LT50为5.04~14.03 d, LYC10和LYC11菌株的毒力最强。
林间应用白僵菌进行生物防治时,其杀虫效果和持久性受到阳光、雨水、温度、湿度、林木表面化学物质和微生物群落等生态因子的影响[24-25],室内筛选出的高毒力的菌株还需要进行林间防治效果测定。因LYC11菌株的产孢能力强于LYC10菌株,且LYC10菌株和LYC11菌株对竹卵圆蝽若虫的致死中时间LT50差异小,故本研究采用LYC11菌株进行林间防治效果测定。结果表明,LYC11菌株在林间施用时对竹卵圆蝽若虫也具有很高的致病能力,但该菌株所有5个浓度分生孢子的累计死亡率均低于室内测定时第13天的结果,这可能是由毛竹林的生态因子造成的不利影响。进一步研究需要明确毛竹林影响球孢白僵菌杀虫效果和持久性的关键生态因子,针对这些关键生态因子,优化菌剂的配方和类型、施用菌剂的方式和时间点,减小生态因子的影响,进而获得最好的杀虫效果。
-
本研究从竹卵圆蝽僵虫体内获得7株球孢白僵菌。室内生物测定表明,LYC10、LYC11、LYC15和LYC4菌株对竹卵圆蝽若虫均具有很高的毒力,LYC10菌株的毒力最强;当分生孢子浓度为108孢子·mL−1时,4个菌株的致死中时间LT50分别为5.04、5.45、6.17、14.03 d。林间生物测定表明,LYC11菌株的回归方程为y=0.234x−1.025,LYC11菌株对竹卵圆蝽若虫的致死中浓度LC50为2.43 × 104孢子·mL−1;接菌后第18 天,LYC11菌株所有5个浓度分生孢子的累计死亡率均低于室内测定时第13天的结果。
竹卵圆蝽致病球孢白僵菌的分离和毒力测定
Isolation and Virulence of Beauveria bassiana Against Hippotiscus dorsalis
-
摘要:
目的 筛选对竹卵圆蝽若虫具有高毒力的致病球孢白僵菌菌株,为竹卵圆蝽的生物防治提供科学依据。 方法 采用组织分离法从竹卵圆蝽僵虫体内分离昆虫病原真菌,通过形态学、DNA条形码相结合的方法对病原真菌进行鉴定,采用生物测定法评价分离真菌菌株的毒力。 结果 从竹卵圆蝽僵虫体内获得7株病原真菌,这些菌株的形态特征、培养性状和ITS rDNA序列均与球孢白僵菌一致,因此,鉴定为球孢白僵菌。室内和林间生物测定结果表明,当分生孢子浓度为108孢子·mL−1时,LYC10、LYC11、LYC15和LYC4菌株对竹卵圆蝽若虫的致死中时间LT50分别为5.04、5.45、6.17和14.03 d;LYC11菌株林间的致死中浓度LC50为2.43 × 104孢子·mL−1。 结论 从竹卵圆蝽僵虫体内分离到7株球孢白僵菌,不同菌株的毒力存在差异,LYC10菌株的毒力最强;LYC11菌株林间的累计死亡率低于室内测定的结果。 Abstract:Objective To select highly pathogenic strain of Beauveria bassiana against Hippotiscus dorsalis nymph, so as to support the biological control of H. dorsalis. Method The entomopathogenic fungi were isolated from muscardine cadaver of H. dorsalis with the tissue isolation approach. The species identification was conducted based on morphological method combined with DNA barcoding. The virulence of B. bassiana isolates was evaluated by bioassay. Result In the study, seven strains of entomopathogenic fungi were isolated from the muscardine cadaver of H. dorsalis. The morphological characteristics, cultural characteristics and ITS rDNA sequences of all isolates were consistent with those of B. bassiana, so these isolates were identified as B. bassiana. The results of bioassay in the laboratory showed that, when the conidia concentration was 108 conidia·mL−1, the LT50 of LYC10, LYC11, LYC15 and LYC4 strains were 5.04 days, 5.45 days, 6.17 days and 14.03 days, respectively. In the field, the LC50 of LYC11 strain was 2.43 × 104 conidia·mL−1. Conclusion In the study, seven strains of B. bassiana were isolated from H. dorsalis. It is showed the pathogenicity of different strains is different, and the pathogenicity of LYC10 strains is the strongest. The cumulative mortality of LYC11 strain in bamboo forest is lower than that measured in the laboratory. -
图 1 竹卵圆蝽僵虫分离的球孢白僵菌形态特征
Figure 1. The morphological characteristics of B. bassiana isolated from H. dorsalis
图 2 球孢白僵菌 ITS rDNA 序列的 PCR 扩增
Figure 2. PCR products of ITS rDNA fragment from B. bassiana.
图 3 竹卵圆蝽若虫接种不同浓度球孢白僵菌后的逐日校正累计死亡率
Figure 3. Corrected cumulative mortality of H. dorsalis over time after inoculation with various
表 1 球孢白僵菌菌株对竹卵圆蝽若虫的致病性和LT50
Table 1. Pathogenicity and LT50 of the tested B. bassiana strains against H. dorsalis larvae
菌株
Strains浓度
Concentrations/
(分生孢子conidia·mL−1)回归方程
Regression
equation决定系数R2
Correlation
coefficient R2LT50/d 累计死亡率
Cumulative mortality/%校正累计死亡率
Corrected cumulative
mortality/%LYC4 1 × 107 y=3.255x + 1.967 0.954 8.55 91.667 81.251 1 × 108 y=2.06x + 2.637 0.802 14.03 83.333 62.499 LYC10 1 × 107 y=8.611x−2.221 0.927 6.90 94.444 87.499 1 × 108 y=3.351x + 2.647 0.892 5.04 100.000 100.000 LYC11 1 × 107 y=4.38x + 0.869 0.942 8.77 91.667 81.251 1 × 108 y=4.295x + 1.838 0.878 5.45 100.000 100.000 LYC15 1 × 107 y=5.338x + 0.145 0.937 8.12 91.667 81.251 1 × 108 y=3.756x + 2.032 0.925 6.17 100.000 100.000  下载: 导出CSV
下载: 导出CSV
-
[1] 徐天森, 林四四, 吕若清. 竹卵圆蝽的研究I. 生物学特性[J]. 林业科学研究, 1988, 1(6):633-640. [2] 徐天森, 王浩杰. 中国竹子主要害虫[M]. 北京: 中国林业出版社, 2004. [3] 黄继育, 何防震, 诸炜荣. 安吉县竹卵圆蝽在竹林中的种群动态模型及预测预报[J]. 林业科技通讯, 2015(11):47-49. [4] 徐天森, 林四四, 吕若清. 竹卵圆蝽的研究II. 天敌及药物防治技术[J]. 林业科学研究, 1989, 2(2):119-123. [5] 尹新明. 杀虫生物学[M]. 北京: 中国农业出版社, 2016. [6] Lacey L A, Frutos R, Kaya H K, et al. Insect pathogens as biological control agents: do they have a future [J] Biological Control, 2001, 21(3): 230-248. [7] Hajek A. Natural enemies: an introduction to biological control [M]. Cambridge, UK: Cambridge University Press, 2004. [8] Chandler D, Bailey A S, Tatchell G M, et al. The development, regulation and use of biopesticides for integrated pest management[J]. Philosophical Transactions of the Royal Society B, 2011, 366(1573): 1987-1998. doi: 10.1098/rstb.2010.0390 [9] 钟武洪, 张贤开, 雷绍清, 等. 竹卵圆蝽生物学特性及其天敌种类调查研究[J]. 湖南林业科技, 2005, 32(5):40-42. doi: 10.3969/j.issn.1003-5710.2005.05.013 [10] 郭 瑞, 何孙强, 王义平. 浙江省竹林害虫竹镂舟蛾和竹卵圆蝽的寄生蜂[J]. 环境昆虫学报, 2016, 38(3):476-481. [11] 熊 琦, 谢映平, 薛皎亮, 等. 桃小食心虫病原真菌的研究及应用[M]. 北京: 中国农业科学技术出版社, 2015. [12] Zimmermann G. Review on safety of the entomopathogenic fungi Beauveria bassiana and Beauveria brongniartii[J]. Biocontrol Science and Technology, 2007, 17(6): 553-596. doi: 10.1080/09583150701309006 [13] Ratnasingham S, Hebert P D N. BOLD: The Barcode of Life Data System (www. barcodinglife. org)[J]. Molecular Ecology Notes, 2007, 7(3): 355-364. doi: 10.1111/j.1471-8286.2007.01678.x [14] 耿显胜, 陈奕洁, 石 坚, 等. 不同寄主竹种上竹瘿广肩小蜂生物学特性研究[J]. 应用昆虫学报, 2019, 56(2):220-226. [15] Bellemain E, Carlsen T, Brochmann C, et al. ITS as an environmental DNA barcode for fungi: an in silico approach reveals potential PCR biases[J]. BMC Microbiology, 2010, 10(1): 189. doi: 10.1186/1471-2180-10-189 [16] Schoch C L, Seifert K A, Huhndorf S, et al. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi[J]. Proceedings of the National Academy of Sciences, 2012, 109(16): 6241-6246. doi: 10.1073/pnas.1117018109 [17] 林乃铨. 害虫生物防治[M]. 北京: 科学出版社, 2010. [18] Lacey L A, Grzywacz D, Shapiro-Ilan D J, et al. Insect pathogens as biological control agents: back to the future[J]. Journal of Invertebrate Pathology, 2015, 132: 1-41. doi: 10.1016/j.jip.2015.07.009 [19] Mascarin G M, Jaronski S T. The production and uses of Beauveria bassiana as a microbial insecticide[J]. World Journal of Microbiology and Biotechnology, 2016, 32(11): 177. doi: 10.1007/s11274-016-2131-3 [20] Jaber L R, Ownley B H. Can we use entomopathogenic fungi as endophytes for dual biological control of insect pests and plant pathogens [J] Biological Control, 2017, 107: 50-59. [21] Rehner S A, Minnis A M, Sung G H, et al. Phylogeny and systematics of the anamorphic, entomopathogenic genus Beauveria[J]. Mycologia, 2011, 103(5): 1055-1073. doi: 10.3852/10-302 [22] Valero-Jiménez C A, Debets A J M, van Kan J A L, et al. Natural variation in virulence of the entomopathogenic fungus Beauveria bassiana against malaria mosquitoes[J]. Malaria Journal, 2014, 13: 479. doi: 10.1186/1475-2875-13-479 [23] Corrêa B, Duarte V D S, Silva D M, et al. Comparative analysis of blastospore production and virulence of Beauveria bassiana and Cordyceps fumosorosea against soybean pests[J]. BioControl, 2020, 65(3): 323-337. doi: 10.1007/s10526-020-09999-6 [24] Jackson M A, Dunlap C A, Jaronski S T. Ecological considerations in producing and formulating fungal entomopathogens for use in insect biocontrol[J]. Biocontrol, 2010, 55(1): 129-145. doi: 10.1007/s10526-009-9240-y [25] Jaronski S T. Ecological factors in the inundative use of fungal entomopathogens[J]. BioControl, 2010, 55(1): 159-185. doi: 10.1007/s10526-009-9248-3 -

 点击查看大图
点击查看大图

图(3) / 表(1)
计量
- 文章访问数: 3243
- HTML全文浏览量: 2092
- PDF下载量: 46
- 被引次数: 0


