



-
木材是木本植物次生木质部长年累积的产物,次生木质部发育是一个动态过程,包括维管形成层分裂、木质部与韧皮部细胞分化、次生细胞壁形成等各个阶段[1]。其中,木质部细胞的分化是木材形成的关键。木本植物中杨树(Populus)凭借其基因组小、适应性强、生长速度快、易于无性繁殖等特点,已被公认是林木分子生物学研究的模式物种[2]。近年来,随着对木质部发育相关研究的逐步深入,已有大量研究表明杨树木质部发育受多种遗传和内源因子调控,如激素、转录因子及多肽信号等[3]。其中,转录因子作为调控木质部发育的关键因子存在,已有大量研究报道转录因子参与调控木质部发育,如:XND1(XYLEM NAC DOMAIN 1)负调控木质部导管分化[4];油菜素内酯对木质部分化起到正向调控作用,且有利于促进应力木的形成[5];生长调节因子PagGRF12a被显性抑制可有效增加木质部宽度,促进木质部发育[6]。综上可见,木质部发育受复杂的转录因子调控,从而影响木材形成过程。
KNOX家族是一类重要的同源异型盒基因,广泛存在并参与生物的发育进程,并通过调控植物分生组织影响不同组织、器官的发育[7]。拟南芥(Arabidopsis thaliana )中,I类KNOX包括4个成员分别是:STM(SHOOT MERISTEMLESS)、KNAT1(KNOTED1-LIKE FROM ARABIDOPSIS THALIANA1)、KNAT2和KNAT6。已有研究表明KNAT2与KNAT6分别在茎顶端分生组织的底部与边缘处表达[8];与此同时,本研究团队前期对‘84K’杨(Populus alba × Populus glandulosa)中KNAT2和KNAT6的同源基因PagKNAT2/6a与PagKNAT2/6b的功能展开研究,已发现PagKNAT2/6b通过直接激活XND1a的表达抑制木质部分化及次生细胞壁的合成[9],但PagKNAT2/6a作为与PagKNAT2/6b进化关系最近的成员,其在杨树木质部发育中的调控功能未知。
本研究为探究PagKNAT2/6a在木质部发育中的功能,以PagKNAT2/6a为研究对象,通过‘84K’杨树遗传转化技术获得异常表达PagKNAT2/6a转基因株系,借助GUS染色实验和实时荧光定量(qRT-PCR)比较分析PagKNAT2/6a与PagKNAT2/6b组织表达特异性,同时借助茎部解剖学分析其异常表达PagKNAT2/6a转基因株系茎中木质部分化差异,通过比较分析PagKNAT2/6a与PagKNAT2/6b在杨树木质部中的表达定位及木质部分化中的功能差异发现该基因在木质部发育中可能存在的功能保守性。本研究对PagKNAT2/6a与PagKNAT2/6b在杨树木质部发育中功能保守性的解析,以期为KNOX类基因在木本植物木质部发育上的保守性和分化性提供理论支持,并为KNOX基因在定向分子设计育种工作开展中提供重要的基因资源。
-
通过NCBI中的BLAST(
https://blast.ncbi.nlm.nih.gov )检索‘84K’杨树(Populus alba × Populus gllandulosa)中PagKNAT2/6a与PagKNAT2/6b基因序列,然后利用MEGA软件中Clustal Co程序进行多序列比对。 -
取生长3~4周的PPagKNAT2/6ba::GUS组培苗整株浸泡于固定液(90%丙酮)中,4 ℃固定过夜;固定后的材料用配置好的GUS染色缓冲液(0.2 mol·L−1磷酸二氢钠,0.2 mol·L−1磷酸氢二钠,2 mmol·L−1铁氰化钾,2 mmol·L−1亚铁氰化钾,现用现配)在冰上洗涤3次,然后将材料转移至GUS染色液(1 mmol·L−1 x-Gluc先溶于DMF,后溶于染色缓冲液,并加0.2%的TritonX-100,现配现用)中,抽真空20~30 min,后置于37 ℃摇床70 r·min−1,染色10 h。最后用75%乙醇进行脱色,取脱色后的茎或叶片,将其固定于5%~6%琼脂中,待凝固后使用振动式切片机切片,后使用显微镜对GUS信号的定位进行观察。GUS染色实验包含2个转基因株系,每个转基因株系设置单株重复。
-
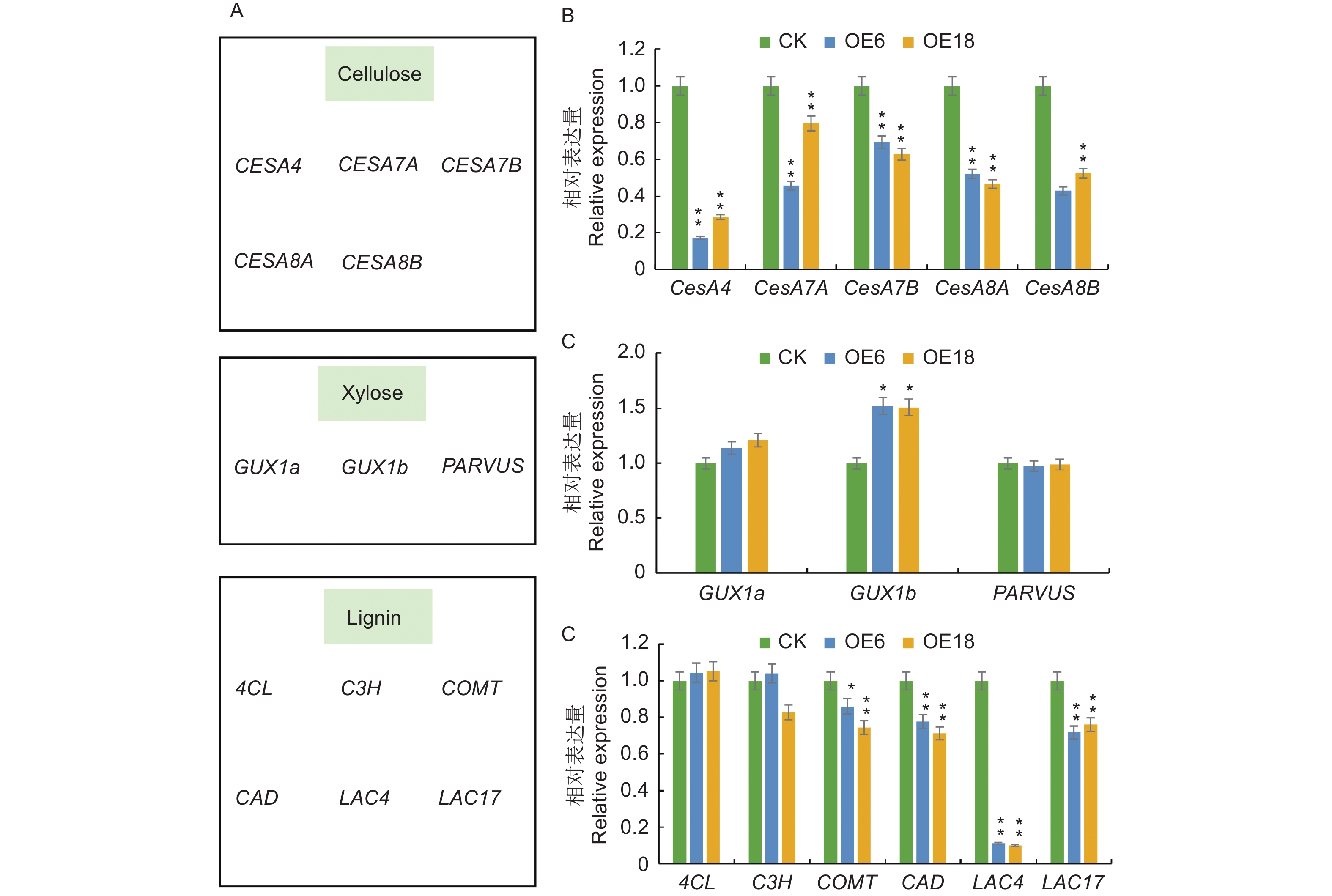
采用华越洋植物RNA提取试剂盒,分别提取‘84K’杨,过表达株系(OE6,OE18)的相同节间总RNA,然后以RNA为模板按照 HiScript II Q RT SuperMix for Qpcr(+ gDNA wiper)试剂盒说明书合成cDNA,并利用 ChamQ Universal SYBR qPCR Master Mix 试剂盒进行qRT-PCR(反应体系:SYBR Premix Ex Taq Tm 5 μL,引物各0.5 μL,cDNA 1 μL,ddH2O 3 μL),以Actin作为内参基因,每个样本设4个生物学重复。最后采用2−△△CT法计算茎植物组织相对表达量。同时,本研究选择了细胞壁三大素(纤维素,半纤维素,木质素)部分合成相关的基因(CESA4、CESA7A、CESA7B、CESA8A、CESA8B、GUX1a、GUX1b、PARVUS、4CL、C3H、COMT、CAD、LAC4、LAC17)通过qRT-PCR技术进行分析PagKNAT2/6a可能的下游靶基因,所有引物见表1。
基因名
Gene nameTranscript ID
in Populus正向引物 (5′→3′)
Forward primers (5′→3′)反向引物 (5′→3′)
Reverse primers (5′→3′)注
NotesKNAT2/6a Potri.008G188700 CGCAGATTGCACGTTTCTTA AGGCCTTTCAAGATCGGATT RT-PCR Actin Potri.001G309500 AAACTGTAATGGTCCTCCCTCCG GCATCATCACAATCACTCTCCGA RT-PCR CesA4 Potri.002G257900 CAGTCCTGTCCTCAGTGCAA CAACACTTCCTGCTGACGAA RT-PCR CesA7A Potri.006G181900 TGCAGTGCTAACCAATGCTC TGTTGGCGTAACGATCATGT RT-PCR CesA7B Potri.018G103900 TGCAGTGCTTACCAATGCTC TGTTGGCGTATCGATCATGT RT-PCR CesA8A Potri.011G069600 AAAACACCTGACGAGGGATG GGCTTTCTTGTGGTGTTGGT RT-PCR CesA8B Potri.004G059600 TGCTGTTTCTTGGCCTCTTT GTATCTTCGGCTGCTTTTGC RT-PCR GUX1a Potri.005G061600 CCAAAAGCTGAGAAGGATGC TGCAGTTTGAAGGCTCAATG RT-PCR GUX1b Potri.007G107200 TGGCATCGTATTCCAAAACA GCAATCGTAGTCTCGGAAGC RT-PCR PARVUS Potri.014G040300 ACATGGACTTGGTGGGGATA TAAGAATCCAAGGCGAATGG RT-PCR 4CL1 Potri.001G036900 CTTTCCTAGGCGCTTCACAC GGCAGAGTCTACGCACATGA RT-PCR C3H Potri.006G033300 GTGGAGGTGCCAAGAAACAT GATTTGGGAAATCTGCCTCA RT-PCR COMT Potri.012G006400 GATGCGCCTGTCATGTTAGA GAGGACCTTGTCCTGGTTCA RT-PCR CAD Potri.009G095800 AAGCTTGATGGCAAGCTGAT CCTCTCGAATGCTGTGTTGA RT-PCR LAC4 Potri.010G193100 GTTTCCGGGACCCACTATTT CCCAGGCTGTATTGGACACT RT-PCR LAC17 Potri.006G087100 AATGTGCCACAAACAGTGGA CGGAGGTGAAGACACCATTT RT-PCR KNAT2/6a Potri.008G188700 GGGGACAAGTTTGTACAAAAAAGCAGGCT
GCATGGAAGGAACTTACGGTCTACACTCAGGGGACCACTTTGTACAAGAAAGCTGGGT
ATCAGTCTTCAGCAAGGAACTGCCCAGAClone Table 1. The primer sequences used in this study.
-
将新鲜杨树茎的节间切成1 cm长,一端用胶水固定于样品架,利用震荡切片机对样品进行切片,切片厚度一般设置为60 μm,将切片收集至ddH2O中,待染色观察。染色时直接将染色液(0.5%甲苯胺蓝)滴到切片上,1 min左右后用蒸馏水冲洗在荧光共聚焦显微镜下观察并拍照。
-
用双面刀片迅速取下野生型及PagKNAT2/6a异常表达的转基因植株新鲜的茎段,后将其置于装有4%多聚甲醛中固定2 h,后经过脱水,临界点干燥,将茎段垂直放置于样品台上,喷金处理使茎段横切面均匀喷上金粉,最后用扫描电镜观察并拍照。
-
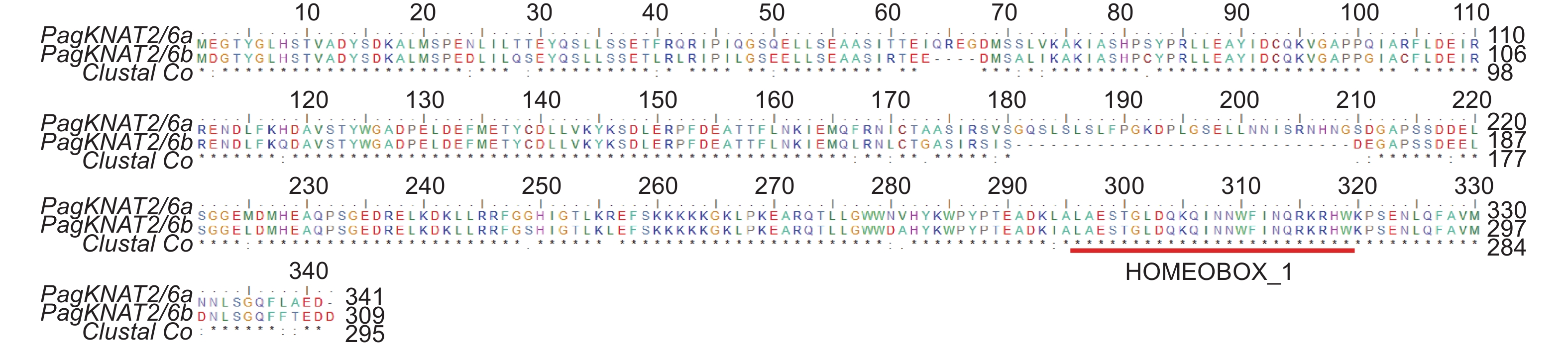
根据PagKNAT2/6a(Pop_A08G045880)与PagKNAT2/6b(Pop_A10G047641)进化关系,本研究首先分析了PagKNAT2/6a与PagKNAT2/6b氨基酸序列的相似性,如图1所示,PagKNAT2/6a与PagKNAT2/6b氨基酸序列高度相似,且都包含HOMEBOX-1保守的同源盒,表明PagKNAT2/6a与PagKNAT2/6b基因同源性相对较高,序列相对保守,为此推测PagKNAT2/6a与PagKNAT2/6b基因在杨树发育中可能存在功能保守性。
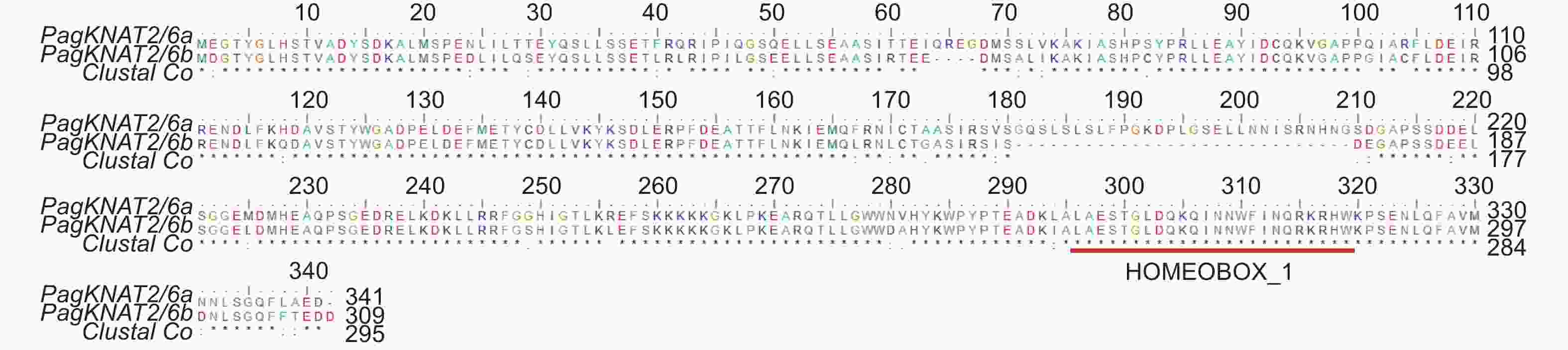
Figure 1. PagKNAT2/6a and PagKNAT2/6b amino acid sequence alignment.
-
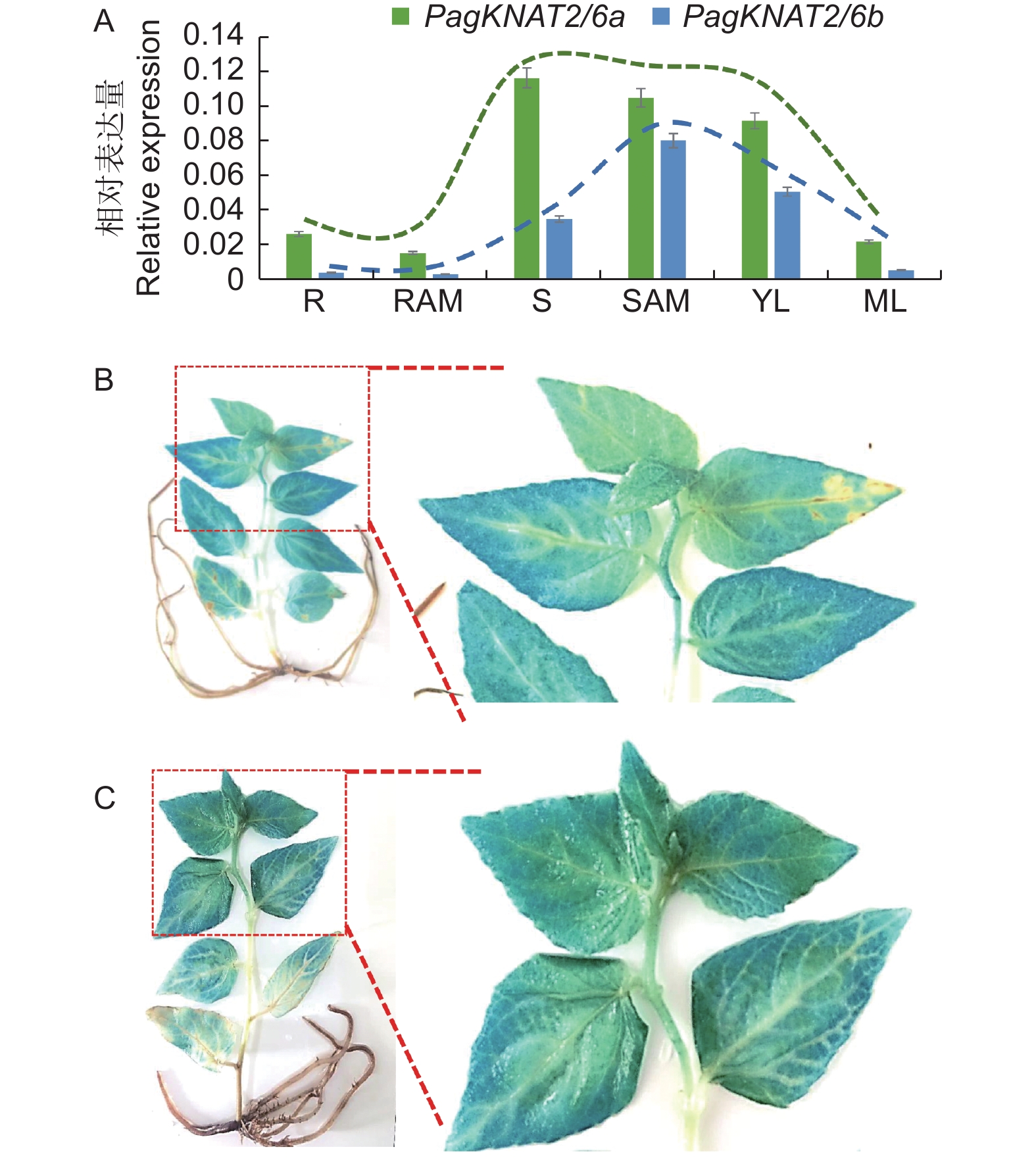
为比较分析PagKNAT2/6a与PagKNAT2/6b在杨树不同组织中的表达特异性,本研究首先借助实时荧光定量PCR技术,检测PagKNAT2/6a与PagKNAT2/6b基因在‘84K’杨树根、根尖、茎、茎尖、幼嫩叶及成熟叶6个部位中的组织表达变化(图2A),结果显示PagKNAT2/6a与PagKNAT2/6b基因均在杨树茎、茎尖以及幼嫩叶片中表现出较高的表达量。与此同时,本研究克隆了PagKNAT2/6a与PagKNAT2/6b基因启动子序列,将其融合至含有GUS报告基因的载体中,通过稳定的杨树遗传转化方法获得PPagKNAT2/6a::GUS与PPagKNAT2/6b::GUS转基因杨树并进行GUS染色实验分析其表达定位(图2B与2C)。结果显示PagKNAT2/6b基因(图2B)在幼嫩节间与叶片中GUS信号强于PagKNAT2/6a(图2C),且在GUS信号观察中发现PagKNAT2/6a与PagKNAT2/6b在幼嫩叶片与幼嫩茎段中的表达均强于成熟叶片与根。
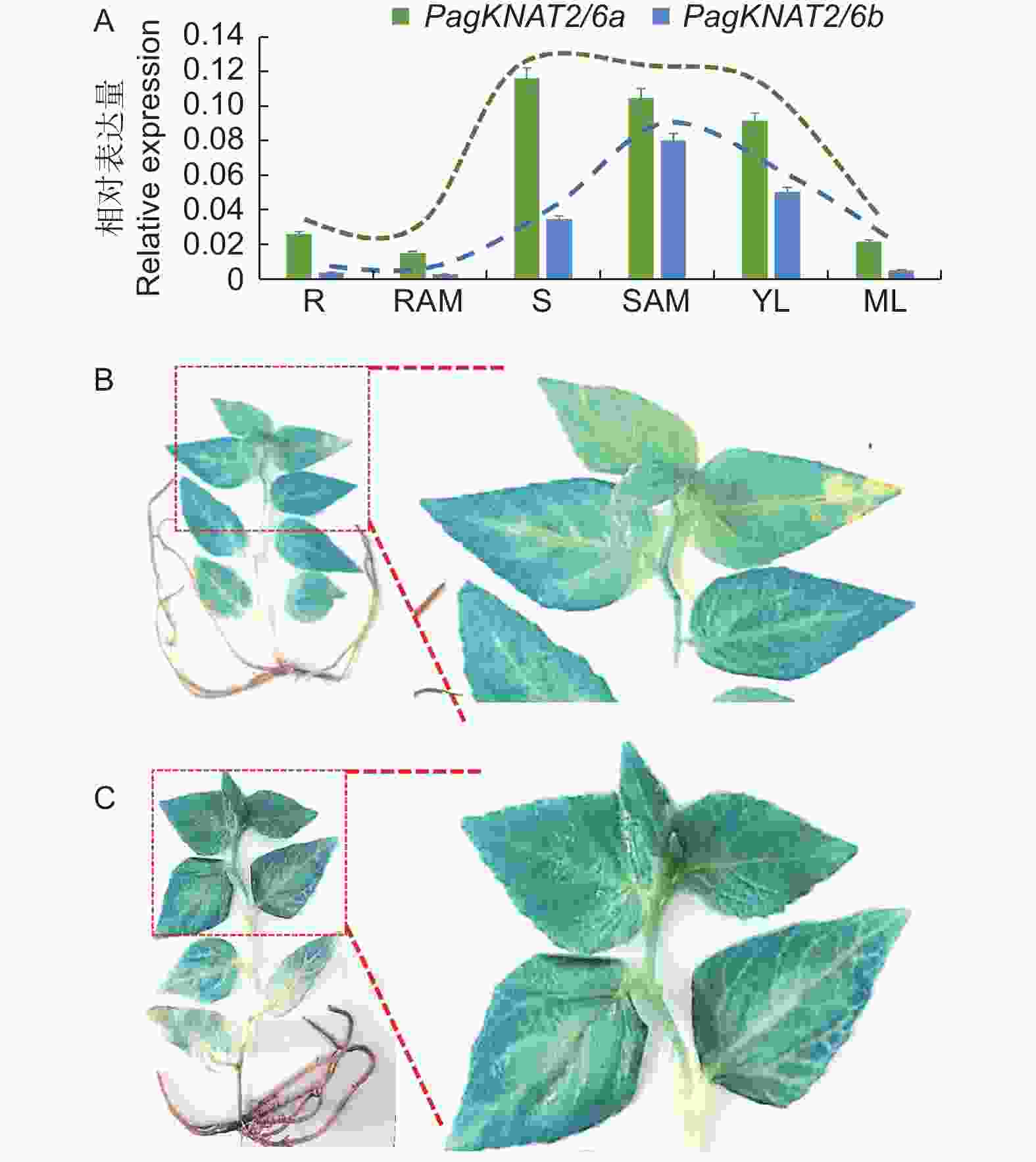
Figure 2. Tissue expression specificity analysis of PagKNAT2/6a and PagKNAT2/6b genes
随后,为深入研究PagKNAT2/6a与PagKNAT2/6b基因在茎中的表达特性,通过GUS染色放大图发现PagKNAT2/6a与PagKNAT2/6b均特异在木质部与韧皮部表达(图3A、C),但利用Popgenie网站分析发现PagKNAT2/6b表达量在木质部表达量显著高于PagKNAT2/6a(图3B、3D)。根据上述结果推测,杨树PagKNAT2/6a与PagKNAT2/6b基因可能均参与杨树茎部木质部发育方面的调控。
Figure 3. PagKNAT2/6a and PagKNAT2/6b genes are highly expressed in stems
-
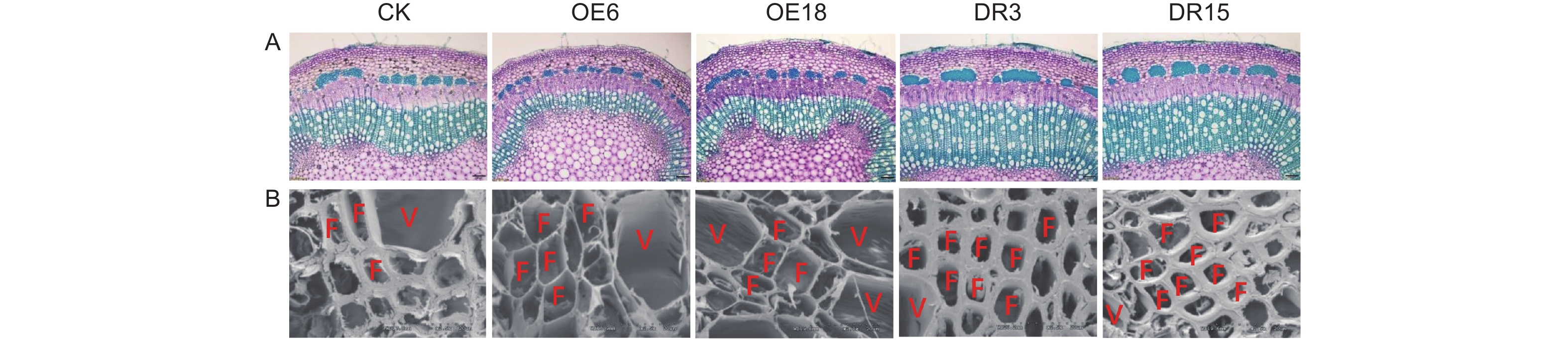
基于PagKNAT2/6a与PagKNAT2/6b在杨树茎中木质部区域特异性表达,为深入比较分析PagKNAT2/6a与PagKNAT2/6b在木质部发育中的功能,本研究在前期分别创制了PagKNAT2/6a过量表达转基因杨树(PagKNAT2/6a OE,OE6与OE18)和PagKNAT2/6a抑制表达转基因杨树(PagKNAT2/6a SRDX,DR3与DR15)转基因株系(已发表)。与此同时,对生长一个月的不同转基因株系的第八节间分别进行切片(图4A)及扫描电镜分析(图4B)。结果显示:PagKNAT2/6a OE株系的木质部宽度和次生壁沉积厚度显著窄于野生型(CK),而PagKNAT2/6a SRDX株系茎中木质部显著宽于CK。该结果与研究团队前期发现的PagKNAT2/6b基因对木质部的影响相似,说明PagKNAT2/6a与PagKNAT2/6b在木质部分化调控上的保守性。
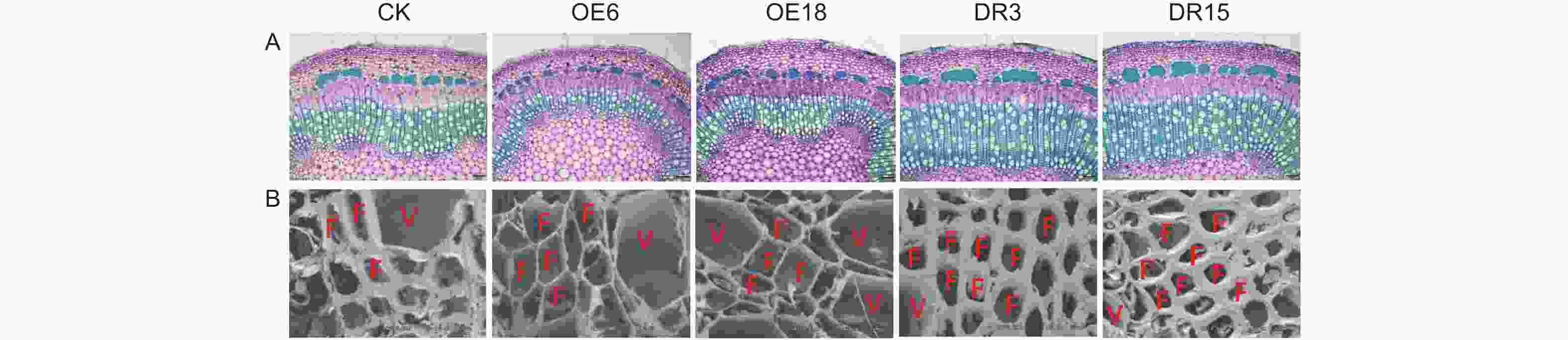
Figure 4. Abnormal expression of PagKNAT2/6a affects woody partialization and xylem cell wall thickness in stems
-
众所周知,次生细胞壁是木质部的主要组成部分,其组成成分对木材品质有直接的影响,基于PagKNAT2/6b基因抑制次生壁合成相关基因的表达从而抑制杨树茎中木质部分化的研究基础[10];为此,推测PagKNAT2/6a基因可能通过调控次生壁合成基因来影响木质部的发育。为进一步解释PagKNAT2/6a与PagKNAT2/6b异常表达与次生壁合成相关基因之间的调控关系,本研究通过qRT-PCR分析了细胞壁三大素(纤维素、半纤维素、木质素)合成相关基因在CK、PagKNAT2/6a OE植株茎中的表达变化(图5)。结果分析发现:相比于CK,参与调控次生壁合成的纤维素、木糖(半纤维素中主要组分)、木质素等相关基因(COMT、CAD、LAC4、LAC17、CesA4、CesA7A、CesA7B、CesA8A、CesA8B)在PagKNAT2/6a OE茎中均表现出下调表达的趋势,其中,纤维素合成相关的CesA类基因的下调趋势最显著(图5C)。根据以上结果可知,PagKNAT2/6a基因通过调控次生壁合成相关基因的表达影响木质部发育,且PagKNAT2/6a与PagKNAT2/6b在对次生壁合成相关基因的调控上具有保守性。
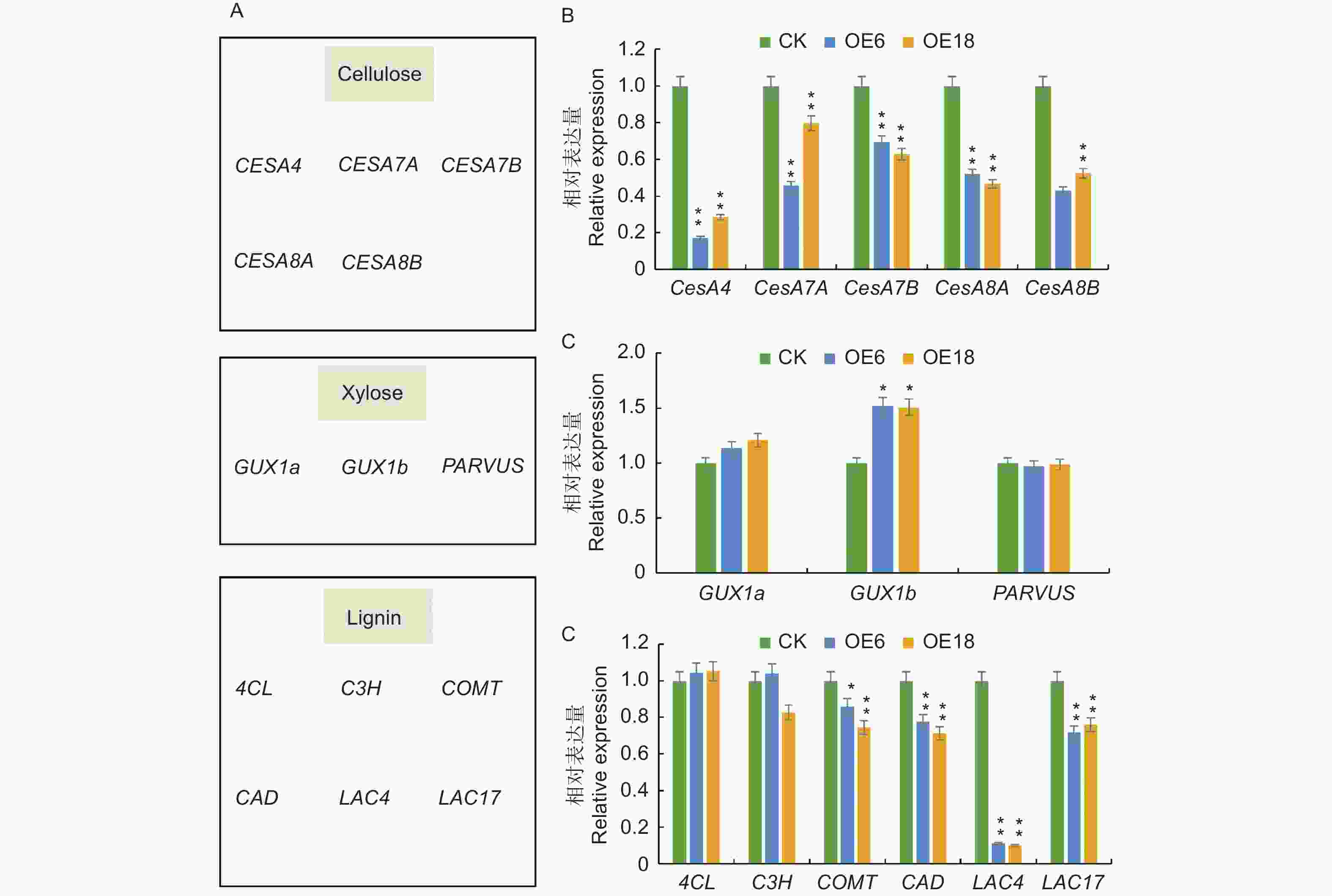
Figure 5. Aberrant expression of PagKNAT2/6a affects the expression of genes related to cell wall synthesis
-
次生木质部由维管形成层细胞通过平周、垂周分裂不断增殖,位于内侧的细胞不断分化,伴随次生壁形成和细胞内容物溶解从而形成成熟的次生木质部[11]。次生木质部发育是木材生产的生物学基础,其发育调控原理一直是林木分子生物学基础研究中关注的核心科学问题。已有研究表明,木质部发育受多种因子调控,如细胞比例,次生细胞壁的结构组成,植物激素和关键基因等。目前,次生壁合成被认为由三级调控网络调控,调控木质部细胞分化的VND类转录因子,其可直接调控第二级转录因子MYB46、MYB83的表达,后者又作为重要开关,调控细胞壁三大素合成相关酶基因的表达[12]。与此同时,已有研究发现KNOX基因参与调控木本植物形成层分化,其中杨树中Ⅰ类KNOX成员ARK1与ARK2均在形成层区域表达[13-14],而PagKNAT2/6a与PagKNAT2/6b在形成层两侧表达,可见不同KNOX类成员分别影响形成层的分裂及子细胞的分化过程[15]。
木材来自形成层的活动,调控形成层活动的基因会影响木质部的发育,杨树中PagKNAT2/6a与PagKNAT2/6b均在形成层两侧区域有较高的表达量,分析PagKNAT2/6b在木质部发育中的功能,发现PagKNAT2/6b过表达会使木质部分化受到抑制[9]。本研究着重比较了PagKNAT2/6a与PagKNAT2/6b在对杨树茎中木质部的发育调控上的异同,结果发现其在木质部发育中的功能存在一定保守性,但从茎部切片结果可知(图4A),PagKNAT2/6b对木质部发育的影响大于PagKNAT2/6a。
-
本研究在探究PagKNAT2/6a基因在木质部发育中的功能,同时对杨树PagKNAT2/6a与PagKNAT2/6b在木质部发育中功能的保守性进行分析,确定了PagKNAT2/6a与PagKNAT2/6b在木质部发育与次生壁合成中的功能保守性,本研究不仅为木材形成调控网络建立提供有效线索,而且为木本植物分子育种工作提供理论基础。
Functional Conservation Existed in PagKNAT2/6a and PagKNAT2/6b During Xylem Development
- Received Date: 2023-06-09
- Accepted Date: 2024-01-22
- Available Online: 2024-04-27
Abstract:
 DownLoad:
DownLoad: