



-
瞬时表达分析是一种快速研究基因功能的方法,T-DNA虽然未被整合到宿主基因组上,但在其进入宿主细胞后,仍可以进行转录和翻译,在转化后12 h左右即可检测到目的基因的表达[1]。瞬时转化体系以其快捷、高效的优势被广泛应用于外源基因表达分析、基因沉默、启动子分析、蛋白互作等许多植物分子生物学领域[2-7]。瞬时表达系统的受体材料主要包括愈伤组织[8-9]、原生质体[10-11]、叶片[12]等。与其它受体材料相比,愈伤组织具有成本低、转化效率高、能够大量增殖培养等优势[13],因此,以愈伤组织为受体材料的瞬时转化体系在植物基因功能研究中具有重要的应用价值。但是,瞬时转化过程中一些参数和培养条件会影响转化效率,如农杆菌菌液的浓度、侵染时间和共培养时间等[14-15]。因此,转化条件的探索对转化体系的建立及转化效率的提高至关重要。
日本落叶松(Larix kaempferi (Lamb.) Carr.)具有材质优良、抗病性强和用途广等特点,是我国重要的人工造林树种。随着日本落叶松基因组的测定[16],日本落叶松中越来越多重要的基因被鉴定出来[17-19],因此,基因功能的解析已成为当前日本落叶松研究的主要目标。遗传转化技术是研究基因功能的重要手段,其中根癌农杆菌介导的遗传转化是最常用的转基因方法之一。自从Rossi等[20]于1993年在烟草中首次成功建立以来,该技术目前已在多种植物中成功应用。遗传转化包括稳定转化和瞬时转化,目前,已经建立了农杆菌介导的日本落叶松胚性愈伤组织稳定转化体系[21-23],对于其瞬时转化体系的研究仍未见报道。
本研究以农杆菌介导的日本落叶松胚性愈伤组织稳定转化体系为基础[21-23],利用携带β-葡糖醛酸酶基因(GUS)的pCAMBIA1305.1载体进行瞬时转化,通过分析侵染液浓度、侵染时间和共培养时间对GUS表达量和酶活性的影响,筛选高效的瞬时转化条件,为日本落叶松基因及其启动子的功能研究提供技术支持。
-
实验材料为本实验室保存的由日本落叶松未成熟种子诱导的胚性愈伤组织[22]。在固体增殖培养基[24]上每隔15 d继代一次,在液体增殖培养基上每7 d继代一次。增殖培养在暗环境(25 ± 2 ℃)下进行。
-
使用CTAB植物基因组DNA快速提取试剂盒(ZP309K-2,北京庄盟国际生物基因科技有限公司)进行总DNA的提取,具体操作按照说明书进行。通过琼脂糖凝胶电泳确定DNA的质量,使用分光光度计对DNA的浓度进行检测。合格的DNA于−80 ℃冰箱保存。
-
利用日本落叶松基因组数据[16],选取LaSCL6起始密码子上游2 500 bp序列进行引物设计。以DNA为模板,利用引物5′-AAACCCTCGTCATGGATTTG-3′和5′-TGACAGCAAAACCAAAAA-3′进行PCR扩增。PCR产物切胶回收后进行TA克隆和测序验证。以测序正确的质粒为模板,利用分别带有Hind III和Nco I酶切位点的引物5′-GACCTGCAGGCATGCAAGCTTAAACCCTCGTCATGGATTTG-3′和5′-TTACCCTCAGATCTACCATGGTGACAGCAAAACCAAAAA-3′进行PCR扩增(下划线表示酶切位点序列)。利用Hind III和Nco I酶分别消化pCAMBIA1305.1载体和PCR产物,酶切产物切胶回收后进行连接和转化,测序验证后获得PLaSCL6::GUS重组质粒。
-
通过冻融法将携带GUS的pCAMBIA1305.1载体(35S::GUS)和重组质粒(PLaSCL6::GUS)转入根癌农杆菌GV3101感受态细胞中。取200
$ \mathrm{\mu }\mathrm{L} $ 菌液,涂于含50 mg·L−1卡那霉素的固体Luria-Bertani(LB)培养基上,28 ℃倒置培养2 d。挑取单菌落于含50 mg·L−1卡那霉素的液体LB培养基中,28 ℃,200 rpm摇床上震荡培养至OD600为0.6~0.8。菌液4 ℃,4 000 rpm,离心10 min后,弃上清,收集菌体。使用含20 mg·L−1乙酰丁香酮的液体增殖培养基重悬菌体,使菌液OD600分别为0.05、0.1和0.2(表1),此时得到的溶液即为侵染液。处理
TreatmentOD600 侵染时间
infection time/min共培养时间
co-culture time/h∆CT值
∆CT valueGUS酶活性
GUS enzyme activity /(U·L−1)1 0.05 1 24 −11.630 0 ± 0.36 j 17.5818 ± 0.69 efgh 2 0.05 1 48 −7.936 7 ± 0.49 fgh 18.0120 ± 1.69 efgh 3 0.05 1 72 −6.237 5 ± 0.25 def 21.4831 ± 1.11 bc 4 0.05 5 24 −3.562 5 ± 0.14 abc 21.2536 ± 1.12 bcd 5 0.05 5 48 −7.530 0 ± 0.23 fgh 24.4378 ± 0.94 a 6 0.05 5 72 −4.440 0 ± 0.13 cd 25.7286 ± 0.50 a 7 0.05 10 24 −4.515 0 ± 0.24 cd 19.6185 ± 2.07 bcde 8 0.05 10 48 −6.755 0 ± 0.09 efg 17.3810 ± 1.82 efgh 9 0.05 10 72 −4.890 0 ± 0.20 cde 13.5370 ± 1.86 jkl 10 0.1 1 24 −2.318 3 ± 0.22 ab 15.3442 ± 1.51 ghijk 11 0.1 1 48 −7.355 0 ± 0.28 fgh 15.4590 ± 1.39 ghijk 12 0.1 1 72 −4.652 5 ± 0.14 cd 18.3276 ± 1.38 defg 13 0.1 5 24 −8.422 5 ± 0.44 gh 16.7499 ± 1.68 efghi 14 0.1 5 48 −6.694 2 ± 0.27 efg 18.7579 ± 1.64 cdef 15 0.1 5 72 −3.570 0 ± 0.2 abc 14.9713 ± 1.55 hijkl 16 0.1 10 24 −8.737 5 ± 0.41 hi 16.1761 ± 1.81 fghij 17 0.1 10 48 −6.612 5 ± 0.21 efg 21.7413 ± 1.76 b 18 0.1 10 72 −4.135 0 ± 0.18 bc 19.7906 ± 1.90 bcde 19 0.2 1 24 −10.403 0 ± 0.11 ij 17.1228 ± 1.55 efghi 20 0.2 1 48 −7.845 0 ± 0.20 fgh 16.8359 ± 1.89 efghi 21 0.2 1 72 −6.022 5 ± 0.09 def 18.3563 ± 1.04 defg 22 0.2 5 24 −10.515 0 ± 0.21 ij 14.2542 ± 1.88 ijkl 23 0.2 5 48 −6.025 0 ± 0.33 def 12.0453 ± 1.72 l 24 0.2 5 72 −2.274 2 ± 0.07 a 16.2048 ± 2.31 fghij 25 0.2 10 24 −10.280 0 ± 0.59 ij 13.0493 ± 1.59 kl 26 0.2 10 48 −6.035 0 ± 0.31 def 17.6104 ± 1.94 efgh 27 0.2 10 72 −3.925 0 ± 0.17 abc 14.9713 ± 1.39 hijkl Table 1. Effects of infection solution concentration, infection time and co-culture time on the expression and enzyme activity of GUS
胚性愈伤组织液体增殖培养7 d后,使用抽滤脱菌装置抽干水分,置于侵染液中进行侵染,侵染时间分别为1、5和10 min(表1)。侵染之后使用抽滤脱菌装置将混合物抽干,之后将愈伤组织转移至含有20 mg·L−1乙酰丁香酮的固体增殖培养基BM中,23 ℃,黑暗条件下分别共培养24、48和72 h(表1),随后收集材料,−80 ℃保存。
-
以瞬时转化的胚性愈伤组织为材料,使用RNA提取试剂盒提取总RNA(ER101,北京全式金生物技术),具体操作按照说明书进行。使用核酸检测仪及琼脂糖凝胶电泳进行RNA浓度及纯度检测。使用反转录试剂盒(P10720,北京全式金生物技术)将检测合格的RNA反转录为cDNA。以cDNA为模板进行半定量RT-PCR和qRT-PCR检测,LaEF1A1为内参基因,引物序列为5′-GACTGTACCTGTTGGTCGTG-3′和5′-CCTCCAGCAGAGCTTCAT-3′[25]。以pCAMBIA1305.1载体中的GUS作为检测基因,引物序列为5′-CGAAGCGAGCAATGTGATGG-3′和5′-GATCCGCAAGACGCATCAAC-3′。
半定量RT-PCR反应体系:1 μL cDNA,12.5 μL KOD酶,引物各0.75 μL,10 μL RNase-free H2O。反应条件:94 ℃ 2 min;98 ℃ 10 s,55 ℃ 5 s,68 ℃ 15 s,32个循环;68 ℃ 2 min。PCR产物通过琼脂糖凝胶电泳进行检测。
qRT-PCR反应体系:12.5 μL TB Green® Premix Ex TaqTM,2 μL cDNA,引物各0.5 μL,9.5 μL RNase-free H2O。反应条件:95 ℃ 30 s;95 ℃ 5 s,60 ℃ 30 s,40个循环;95 ℃ 10 s;65 ℃ 5 s;95 ℃ 5 s。每个反应进行3次重复,表达量以∆CT值体现,结果以平均值 ± 标准差(Mean ± SD)的形式表示。
-
将−80 ℃保存的实验样品置于液氮中研磨成粉末,以1∶9的比例置于0.01 mol·L−1 PBS缓冲液中,充分混匀后,4 ℃离心15 min,5 000 rpm,取上清移入新的管中,放到冰上备用。之后使用分光光度计检测上清中总蛋白含量,随后置于−20 ℃备用。使用β-葡萄糖苷酸酶酶联免疫分析试剂盒(LB5023A,武汉力博瑞生物科技有限公司)进行GUS酶活性检测,具体操作步骤按照说明书进行。使用酶标仪在450 nm波长下测定标准品与待测样品的吸光度,以标准品的浓度为横坐标,OD450处的值为纵坐标,使用Excel绘制出标准曲线。根据待测样品的OD450处的值计算样品相应的浓度,再乘以稀释倍数,即为样品总蛋白的浓度。每个样品检测3次,检测结果以平均值 ± 标准差(Mean ± SD)的形式表示。
-
使用PlantCARE(
http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ )对启动子CaMV35S和PLaSCL6的顺式作用元件进行分析。使用TBtools软件绘制启动子元件图。 -
采用SPSS软件进行显著性分析。柱状图使用Origin 2021软件绘制。
-
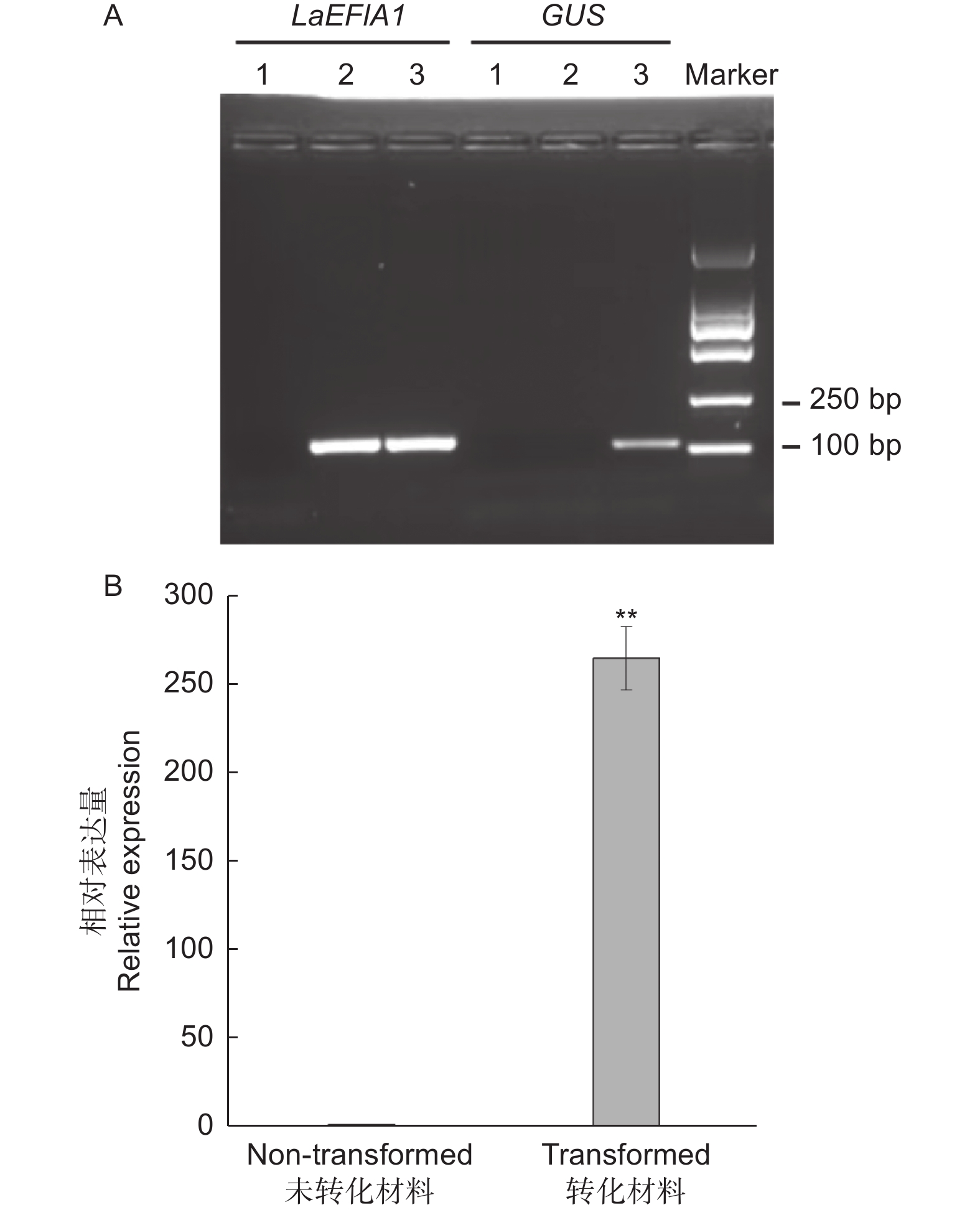
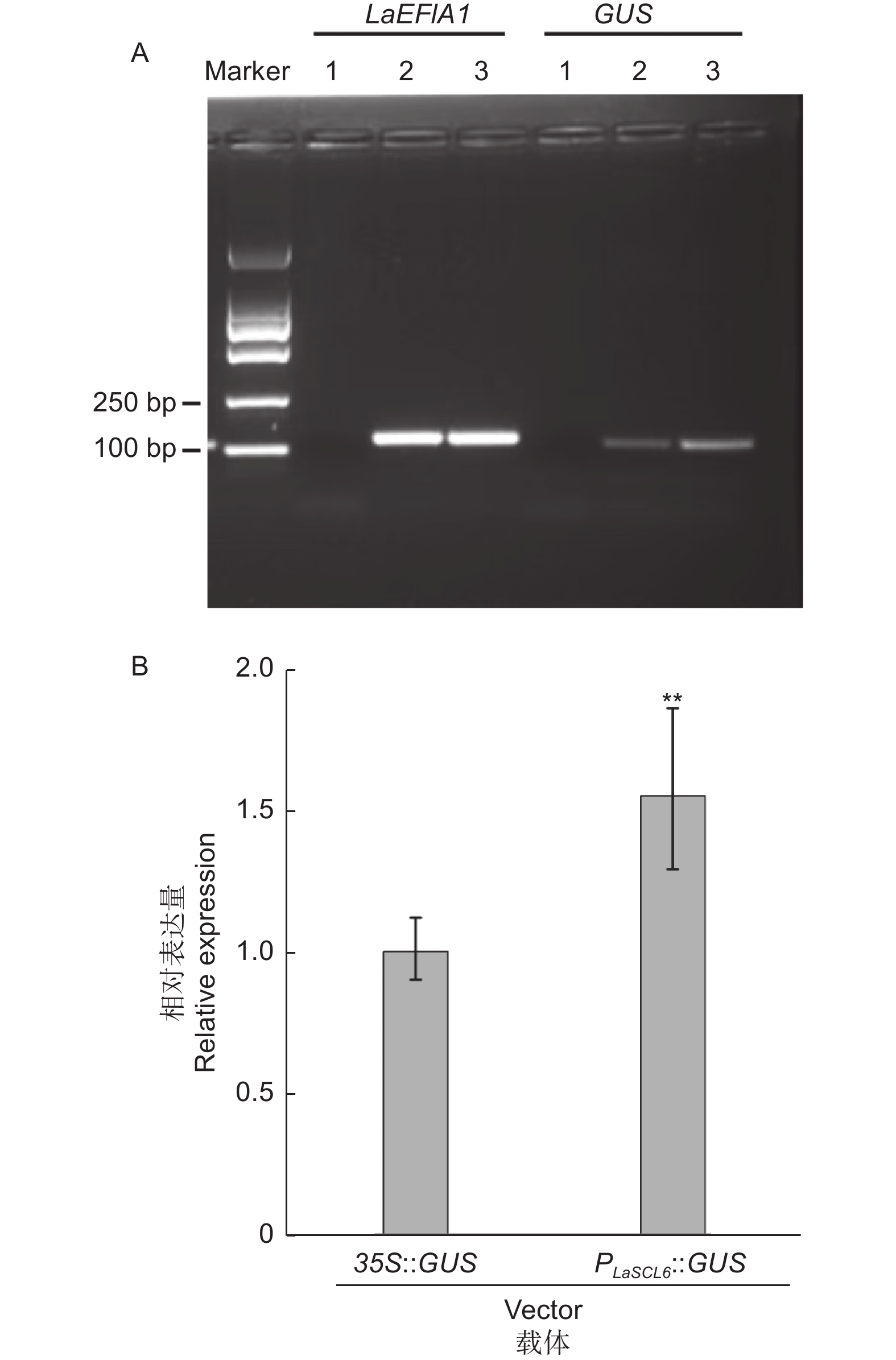
使用载体35S::GUS,以OD600=0.1的侵染液侵染10 min,共培养48 h为条件进行瞬时转化。半定量RT-PCR结果显示,在转化的材料中可以检测到GUS的扩增片段,而在未转化的材料中未检测到(图1A)。qRT-PCR分析也进一步验证了半定量RT-PCR的结果(图1B)。这两个结果表明,以 OD 600=0.1的侵染液侵染10 min,共培养48 h为瞬时转化条件,载体35S::GUS可以导入日本落叶松胚性愈伤组织的细胞中,且CaMV35S启动子可以驱动GUS的表达。
Figure 1. Semi-quantitative (A) and quantitative (B) RT-PCR analysis of GUS driven by CaMV35S
-
侵染液浓度、侵染时间和共培养时间分别设计了3个梯度,使用载体35S::GUS进行瞬时转化。qRT-PCR结果显示,GUS在不同的条件下均能表达,但表达量差别明显(表1)。当侵染液OD600=0.2,侵染5 min,共培养72 h时,GUS的表达量最高,∆CT值为−2.274 2;当侵染液OD600=0.05,侵染1 min,共培养24 h时,GUS的表达量最低,∆CT值为−11.630 0。GUS酶活性在所有材料中均能被检测到,但在不同的条件下,其大小不同(表1)。当侵染液OD600=0.05,侵染5 min,共培养72 h时,GUS酶活性最高,为25.728 6 U·L−1;当侵染液OD600=0.2,侵染5 min,共培养48 h时,GUS酶活性最低,为12.045 3 U·L−1。GUS表达量较高的5个材料是处理4、10、15、24和27,而酶活性较高的5个材料是处理3、4、5、6和17。这些结果表明,处理4中,即以侵染液OD600=0.05,侵染5 min,共培养24 h为转化条件时,GUS的表达量和酶活性都比较高。
-
分别使用载体35S::GUS和重组质粒PLaSCL6::GUS,以OD600=0.05的侵染液侵染5 min,共培养24 h为条件进行瞬时转化。半定量RT-PCR结果表明,两种转化材料中都可以检测到GUS的扩增片段,但转PLaSCL6::GUS的材料中电泳条带相对较亮(图2A)。qRT-PCR结果进一步表明,转PLaSCL6::GUS的材料中GUS表达量是转35S::GUS的1.55倍(P <0.01)(图2B)。
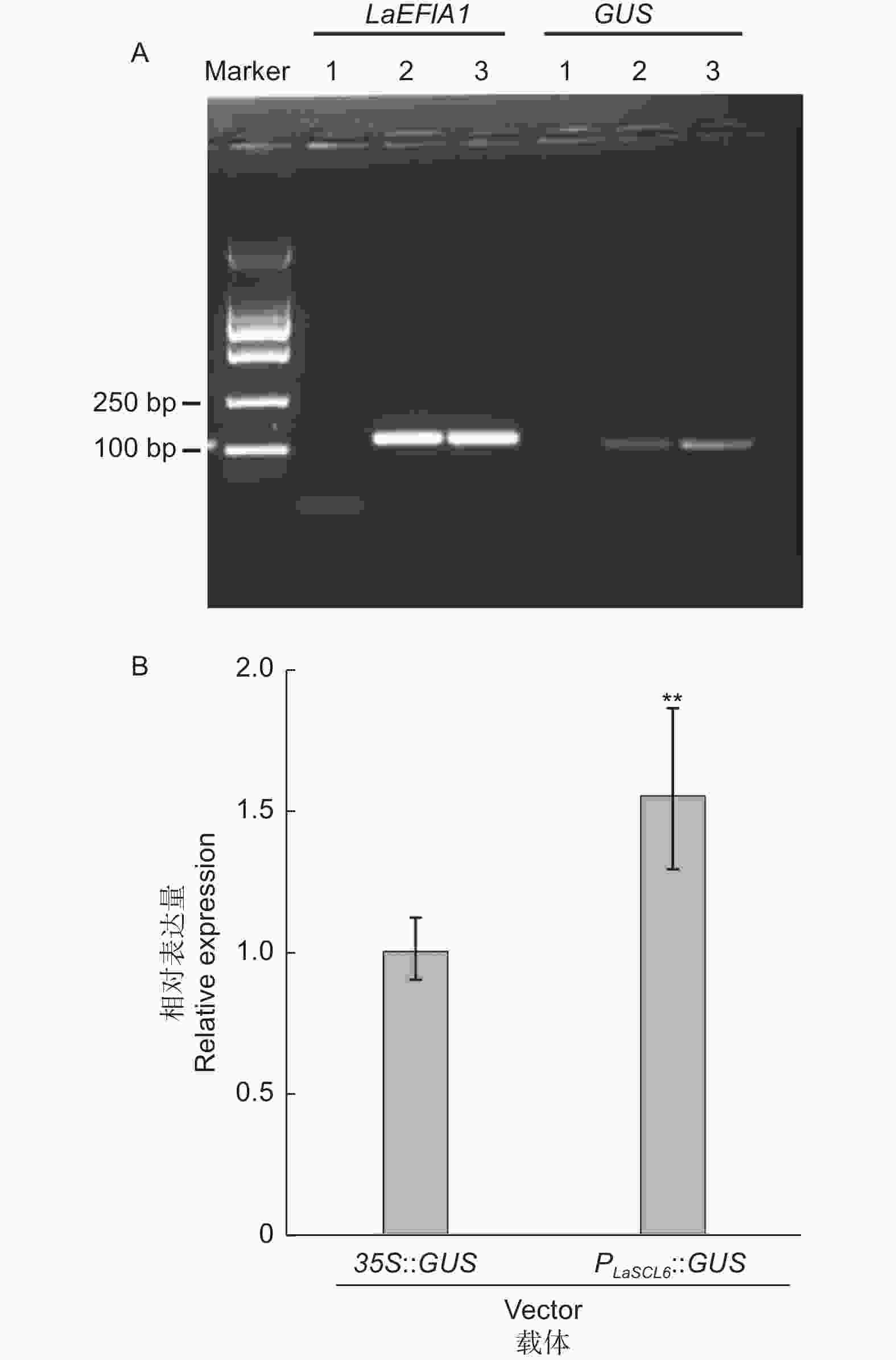
Figure 2. Semi-quantitative (A) and quantitative (B) RT-PCR analysis of GUS driven by promoter CaMV35S or PLaSCL6
-
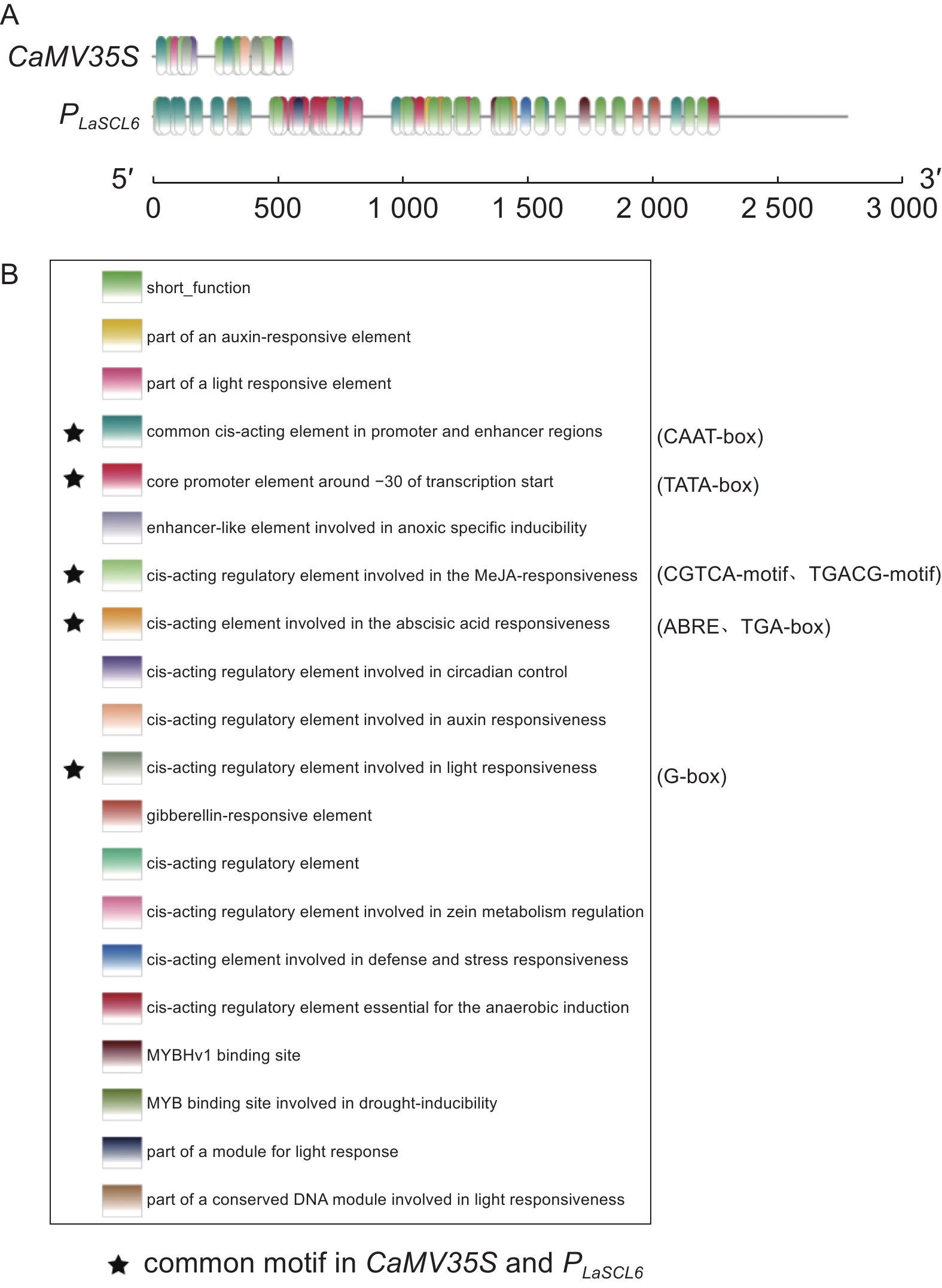
根据预测结果,PLaSCL6上调控元件的数量和种类均要明显多于CaMV35S。PLaSCL6具有132个调控元件,而CaMV35S仅有28个调控元件(图3A)。PLaSCL6上TATA-box有45个,而CaMV35S上仅有3个。此外,PLaSCL6中包含19种顺式作用元件,而CaMV35S上有11种,它们共有的调控元件有7种,分别是激素响应元件(ABRE、TGA-box、)、光响应元件(G-box)、参与MeJA反应的元件(CGTCA-motif、TGACG-motif)、−30附近的核心启动子元件(TATA-box)和常见的顺式作用元件(CAAT-box)(图3B)。此外,PLaSCL6上还包含多种其它顺式作用元件,如激素响应元件(GARE-motif、P-box、CGTCA-motif、TGACG-motif)、光响应元件(AE-box、Box 4、Box II、G-Box、I-box)、厌氧诱导必须的调控元件ARE、干旱诱导相关元件MBS和参与防御和应激反应的调控元件TC-rich-repeats等。
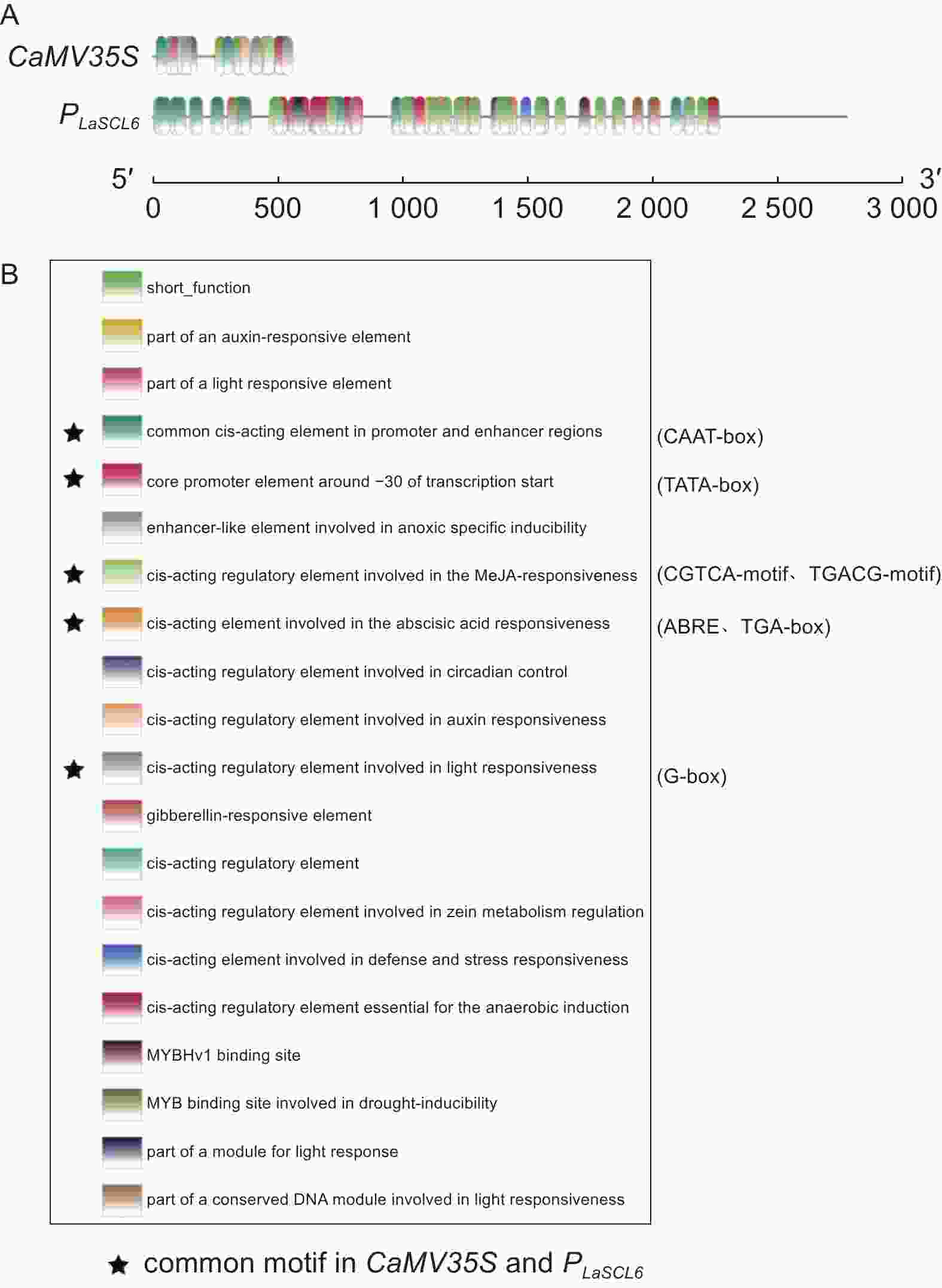
Figure 3. Analysis of cis-acting elements of the CaMV35S and PLaSCL6 promoters
-
植物的遗传转化受到多种因素的影响,要建立一个高效的瞬时转化体系,需对影响遗传转化的多种因素进行全面的探索与优化[26]。彭绿春等认为菌液浓度对云南杜鹃(Rhododendron yunnanense Franch.)愈伤组织中GUS的瞬时表达有显著影响[27]。侵染时间、共培养时间也是影响转化效率的重要因素[28],因为侵染时间过长容易导致外植体死亡,而侵染时间过短则受体材料上不能够附着足够的菌液[29],共培养时间的长短又直接影响着目的基因的整合和转化[30]。本研究以日本落叶松胚性愈伤组织为受体材料,探讨了农杆菌介导的瞬时转化体系中侵染液浓度、侵染时间和共培养时间对转化效率的影响。从GUS的表达量和酶活性来看,不同组合处理后转化效率差别较大。综合分析后,发现以侵染液浓度OD600为0.05,侵染5 min,共培养24 h为转化条件时,GUS的表达量和酶活性都比较高。研究结果也再次表明掌握好侵染液浓度、侵染时间和共培养时间有助于提高瞬时转化效率。另外,上述条件不仅可以用来进行日本落叶松胚性愈伤组织的瞬时转化,也为其它针叶树瞬时转化体系的建立和优化提供参考。
启动子是调控基因表达的重要元件,它的调控作用是由多层次、多因素共同作用的结果。对启动子结构、功能和作用机制的研究不仅有利于了解基因调控模式和信号传递途径,而且对于了解植物的生长发育机制和对外界环境的响应具有重要意义[31]。由于对日本落叶松基因的启动子研究不够深入,导致其在应用中具有不确定性。有些外源启动子在转基因植物中不一定能够诱导目的基因高效表达。例如,外源性启动子hsr203J在柑橘(Citrus sinensis Osbeck)中并未被激活[32]。落叶松PLkF3H2启动子的活性在本生烟草(Nicotiana benthamiana Domin)中略弱于CaMV35S启动子,并且随着PLkF3H2启动子长度的缩短,其活性降低[33]。因此,在未来日本落叶松启动子的克隆及应用研究仍然是重中之重。上述转化条件在启动子活性检测上的成功应用证明了该转化体系的高效性和稳定性。在此基础上发现CaMV35S活性低于PLaSCL6,这可能与PLaSCL6是落叶松内源启动子有关,也可能与PLaSCL6较CaMV35S具有更多的顺式作用元件有关(图3)。这一结果不仅验证了上述转化体系的有效性,也为日本落叶松遗传转化提供了一个候选启动子。
-
本研究分析了日本落叶松胚性愈伤组织瞬时转化过程中侵染液浓度、侵染时间及共培养时间对转化效率的影响。结果表明,当OD600=0.2的侵染液侵染5 min,共培养72 h时,GUS的表达量最高,∆CT值为−2.2742;当OD600=0.05的侵染液侵染5 min,共培养72 h时,GUS酶活性最高,为25.7286 U·L−1。综合考虑GUS的表达量和酶活性,当OD600=0.05的侵染液侵染5 min,共培养24 h时转化效率较高,这一条件可以用来进行日本落叶松胚性愈伤组织的高效转化。研究结果优化了日本落叶松瞬时遗传转化体系,不仅为日本落叶松的基因功能研究提供了技术支持,也为其它针叶树遗传转化体系的建立和完善提供了参考。
Optimization and Application of Transient Transformation System of Larix kaempferi
- Received Date: 2023-10-07
- Accepted Date: 2024-01-03
- Available Online: 2024-04-01
Abstract:

 DownLoad:
DownLoad: