



-
“演替”一词用于描述不同类型的植物在空间和时间上的不同尺度上的变化[1],生态演替是指随着时间的推移,群落中的一些个体被另外一些个体,或者一种植被群落类型被另外一种植被群落类型所代替,是生态系统一个动态的复杂的变化过程。在演替过程中,生态系统里的植被的物种多样性、优势种等都会发生显著的变化。森林演替的实质是群落中优势树种发生明显改变,引起整个森林组成的变化过程。这种变化过程从总体上讲是物种生态对策的差异,是由于物种特性的不同所致。而任何一个物种在不同的生境条件下,其适应和竞争能力的发挥有很大的差异,物种特性是在一定生境条件下长期进化适应的结果。通常情况下,森林生态系统为螺旋式上升演替,最终形成与当地气候匹配的顶级群落[2]。在顶级森林生态系统中,树木是决定森林生物多样性全波光谱的最主要成分,树种的分布类型决定着森林群落的结构[3]。
林火是森林生态系统中重要的干扰因子,大面积森林火烧后森林环境发生急剧变化,导致森林生态平衡的破坏[4-5]。林火作为一种重要的自然干扰,被认为是森林群落演替的重要驱动力,由火干扰引发的植被演替在塑造植物群落的结构、分布和维持其物种多样性等方面起着重要作用[6]。植被对火干扰的响应及火后植被更新动态有助于理解火的作用及预测火后植物种群结构和功能变化[7-8]。甘肃农业大学林火生态团队已对火烧迹地植物的群落结构、群落特征、植物多样性、树种种间关联性、植物生态位特征、土壤有机碳、土壤酶活性变化特征进行了研究[9-12]。分析火烧迹地上不同演替梯度上植物群落的数量特征,对于研究火烧迹地发生的次生演替机理具有重要意义。
-
研究区位于青藏高原东北边缘迭山北坡甘肃迭部县境内的迭部林区达拉沟林场和益哇沟林场,样地范围为34°10′40.66″~34°10′47.30″ N,103°12′48.69″~102°12′57.43″ E,海拔2 981~3 408 m。北亚热带与青藏高原东部边缘高寒气候的过渡性气候,年平均温度为7.5 ℃,年平均降水量 568 mm,年平均蒸发量 1 444.2 mm,年平均日照时数 2 308.0 h,年平均风速为1.8 m·s−1,年平均气温16.3 ℃,年相对湿度52%~76%,年均无霜期 134 d。受区域小地形影响,气候垂直变化明显。云杉-巴山冷杉林火烧迹地的坡向以北坡为主,火烧迹地草本层优势种有高山冷蕨(Cystopteris montana (Lam.) Bernh. ex Desv.)、野草莓 (Fragaria vesca L.)、密生薹草(Carex crebra V. Krecz.) 和总状橐吾(Ligularia botryodes(C. Winkl.) Hand.-Mazz.)等;灌木层优势种有菰帽悬钩子(Rubus pileatus Focke)、美丽悬钩子(R. amabilis Focke)、唐古特忍冬(Lonicera tangutica Maxim.)和扁刺蔷薇(Rosa sweginzowi Koehne)等;乔木层优势种为巴山冷杉(Abies fargesii Franch.)、云杉(Picea asperata Mast.)、岷江冷杉(A. fargesii var. faxoniana (Rehder & E. H. Wilson) Tang S. Liu)、青海云杉(P. crassifolia Kom.)等。
-
本文采用空间替代时间的方法来研究云杉-巴山冷杉林火烧迹地植物群落的演替。2019年7月—8月,通过查阅白龙江林业管理局林火记载文献,实地初步调查云杉-巴山冷杉林火烧迹地植被恢复现状,选取环境条件相似又具备演替梯度的4类植物群落(火后演替5 a、15 a和23 a及作为对照的顶级群落Climax)作为一个演替系列。
在2020年和2021年7~8 月完成了野外植物群落调查。根据不同演替阶段植物群落物种组成及其结构的复杂程度,基于火烧迹地地形,在5 a、15 a和23 a各设置6个20 m × 20 m 乔木样地(下坡位3个,中坡位2,上坡位1个);在与5 a、15 a和23 a 毗邻的Climax各设置3个样地(下坡位、中坡位和上坡位各设置1个)。记录样地的海拔、坡度、坡向、地理坐标;按照一定顺序挂牌编号,调查乔木层(胸径(DBH)≥ 1 cm 的木本植物),记录植物名称、DBH、树高、冠幅、活枝下高、生长状况、健康状况,同时记录火后残留树桩;采用“五点法”样方调查法,在乔木样地内,在东、南、西、北、中各设置5个5 m × 5 m 灌木样方,确定灌木名称、平均高度、株数、盖度和生活型;同时记录乔木树种幼苗的株数、高度、地茎;沿每个灌木样方对角线,设置3个1 m × 1 m的草本样方,确定草本植物的种类、数量、盖度、平均高度。
-
群落中物种优势度由重要值确定[11];根据研究区植物群落分布特征,选取Margalef丰富度指数、Shannon-Wiener多样性指数、Simpson优势度指数、Pielou均匀度指数4个有代表性的指标来表征群落α多样性;群落的异质性选取异质性指数HI[13];不同演替阶段群落间的β多样性选取Whittaker指数(0, 1)[14]。各指标的计算公式具体如下:
重要值=(相对高度+相对密度+相对盖度)/ 3
式中:相对高度 = 某一植物种的高度 / 各植物种高度之和 × 100;相对密度 = 某一植物种的个体数 / 全部植物种的个体数 × 100;相对盖度 = 某一植物种的盖度 / 各植物种的分盖度之和 × 100。
Margalef指数(Ma):
Shannon-Wiener指数(H'):
Simpson指数(P):
Pielou指数(E):
Whittaker指数(βH1
): 式中:S为出现在样方内的物种数;N 是样方数;
$ {P}_{i}={N}_{i}⁄N $ 为种i的相对重要值;$ \stackrel{-}{\alpha } $ 是平均样本的多样性。 -
采用软件 R 4.2.0 完成计算群落物种重要值、α物种多样和β物种多样性并作图;其中α物种多样性和β物种多样性在vegan包中完成。
-
对于不同的云杉-巴山冷杉林火烧迹地,5 a群落物种数最大为106种,23 a群落物种数最小,为64种。从科的分布水平来看,不同演替阶段的群落植物科的数量表现出先减少后增加的特性。5 a群落科数最多,为45科;随着火后群落的演替,从15 a、23 a到顶级群落,植物科的数量呈增加的趋势,各群落分别有32科、33科和43科(表1)。在群落的不同演替阶段,蕨类都为1科1属1种,只不过在5 a 群落中为蹄盖蕨科 Athyriaceae的中华蹄盖蕨(Athyrium sinense),而在其它3个演替阶段均为冷蕨科 Cystopteridaceae的高山冷蕨(Cystopteris montana)。在5 a、15 a、23 a和Climax 群落中,裸子植物占比分别是0.94%、2.94%、4.69%和6.98%,双子叶植物的占比分别是92.45%、91.18%、87.50%和83.72%,这表明云杉-巴山冷杉林火烧迹地随着演替的推移,不同群落中裸子植物种的占比逐渐增加,而双子叶植物的占比逐渐减少。同时说明,对于云杉-巴山冷杉林火烧迹地,群落的植物组成以双子叶植物为主。
演替阶段
Succession stages蕨类植物
Pteridophyte裸子植物
Gymnosperm被子植物 Angiosperm 合计
Total单子叶植物
Monocotyledon双子叶植物 Dicotyledon 5 a 科 Family 1 1 5 38 45 属 Genus 1 1 6 78 86 种 Species 1 1 6 98 106 15 a 科 Family 1 1 3 27 32 属 Genus 1 2 3 49 55 种 Species 1 2 3 62 68 23 a 科 Family 1 1 3 28 33 属 Genus 1 2 4 43 50 种 Species 1 3 4 56 64 Climax 科 Family 1 2 6 34 43 属 Genus 1 4 6 56 67 种 Species 1 6 7 72 86 Table 1. The species composition of plant communities at different succession stages
从表2 可以看出,对于不同演替阶段的植物群落,植物个体数量随群落演替逐渐减少,5 a群落植物个体总数最大为4 008 ± 298株·(400 m2)−1,Climax植物的个体总数最小为1 759 ± 90株·(400 m2)−1;草本层植物个体数量随着演替进程逐渐减少,而灌木层植物个体数量随着演替表现出先增加后减少。在5 a 和 15 a 中,零星分布有乔木幼苗36 ± 27株和98 ± 82株,平均株高分别是15.64 cm和21.08 cm,这些幼苗由根蘖苗和实生苗组成,根蘖苗以红桦(Betula albosinensis Burk.)、白桦(Betula platyphylla Sukaczev)、山杨(Populus davidiana Dode )为主,为聚集分布型;实生苗有云杉(Picea asperata Mast.)和巴山冷杉(Abies fargesii Franch.),为随机分布型,但没有形成乔木层。在 23 a 和Climax中,乔木的平均高度大于灌木,成为各演替阶段的优势群落(附表1、附表2)。发生重度林火后,云杉-巴山冷杉林火烧迹地表现出几乎一致的土壤和小气候环境,不同生态位的植物能在短时期侵入并定居,植物物种数达到最高。随着光照和土壤环境对植物生长的过滤,植物种间竞争加剧,植物个体数量呈现逐年下降趋势。同时,云杉-巴山冷杉林火烧迹地次生演替初期,植物群落并未形成乔木层,顶层仍以灌木为优势群落,随着群落的正向演替,乔木层优势凸显,云杉和巴山冷杉最终成为建群种和优势种。
演替阶段
Succession stage生活型(种)
Life form (Species)个体数量/(株·(400 m2)−1)
Numbers of individual乔木
Arbor灌木
Shrub草本
Herbage总计
Total乔木
Arbor灌木
Shrub草本
Herbage总计
Total5 a - 24 ± 3 76 ± 11 106 ± 14 - 131 ± 27 3 579 ± 256 4 008 ± 298 15 a - 17 ± 1 46 ± 13 68 ± 14 - 196 ± 32 2 773 ± 147 3 205 ± 187 23 a 7 ± 0 18 ± 2 39 ± 6 64 ± 8 151 ± 11 204 ± 41 2 077 ± 210 2 432 ± 262 Climax 12 ± 0 35 ± 6 39 ± 4 86 ± 10 112 ± 7 171 ± 18 1 476 ± 65 1 759 ± 90 Table 2. Life type composition of plant communities at different succession stages
株 -
附表1、附表2列出了4个植物群落草本层和灌木层重要值,附表3列出了23 a和Climax植物群落乔木层重要值等信息。
演替阶段
Succession stages物种
Species株数
Individual高
Height/cm盖度/%
Coverage重要值
Important value23 a 青海云杉 Picea crassifolia 64±11 179.00±132.50 28.26±12.65 32.86±5.63 红桦 Betula albosinensis 18±12 205.55±47.86 8.08±4.55 13.08±5.32 巴山冷杉 Abies fargesii 17±14 215.00±55.00 6.66±1.08 12.36±1.85 山杨 Populus davidiana 16±10 214.00±156.38 6.52±6.32 12.05±2.46 白桦 Betula platyphylla 13±3 197.80±58.85 7.45±1.23 11.47±0.08 云杉 Picea asperata 16±11 188.00±40.85 5.56±1.24 10.93±2.31 腺柳 Salix chaenomeloides 7±5 175.25±40.65 2.89±0.98 7.26±0.85 顶级群落 Climax 巴山冷杉 Abies fargesii 39±21 1 928.50±780.50 17.48±5.38 23.32±3.85 云杉 Picea asperata 15±8 1 608.50±500.00 12.98±4.23 13.61±4.08 岷江冷杉 Abies faxoniana 13±6 1 667.80±450.00 13.17±2.85 13.23±1.78 紫果云杉 Picea purpurea 4±1 2 135.60±566.50 15.36±3.78 12.59±2.89 山杨 Populus davidiana 9±8 1 100.00±480.00 9.35±2.58 9.12±4.56 白桦 Betula platyphylla 12±10 809.25±250.55 1.13±0.08 6.06±0.17 锐齿槲栎 Quercus aliena var. acuteserrata 4±1 896.00±50.80 4.71±0.95 5.3±0.12 旱榆 Ulmus glaucescens 3±1 1 200.00±175.65 3.25±0.03 5.2±0.30 蒙古栎 Quercus wutaishanica 3±1 758.25±38.50 3.18±0.06 4.05±0.03 红桦 Betula albosinensis 7±2 249.88±40.50 1.26±0.03 3.21±0.01 大果圆柏 Juniperus tibetica 1±1 452.50±58.36 1.56±0.01 2.05±2.05 侧柏 Platycladus orientalis 1±1 270.40±82.85 0.72±0.09 1.26±1.26 黄花柳 Salix caprea 1±1 123.87±25.65 1.02±0.04 1.01±1.01 Table 3. Important value of arborescent stratum
-
5 a 草本层物种丰富,多达76种,其中密生薹草作为先锋物种占绝对优势,重要值为12.97。前10种植物的重要值为40.13,表明该群落中除密生薹草外其他植物的分布较为均匀,每种植物的优势度不高,光照和土壤理化性质对植物的过滤较低,草本植物种间竞争不强。15 a、23 a 和Climax中,密生薹草和高山冷蕨其重要值之和 ≥ 24.00,为3个群落草本层优势种;前10种植物的重要值之和分为别是63.47、64.59、59.16,表明随着火烧迹地植被的正向演替,生态位幅宽的植物逐渐成为优势种,少数植物的生长优势增强,种间竞争加剧,个体分布趋向聚集性分布。
-
5 a灌木层生长时间最短,许多物种刚刚入侵,种间竞争尚不明显,灌木层发育良好,灌木种间重要值的差异不大,前10种灌木的重要值之和仅为63.50,重要值的平均值6.35接近于箭竹重要值(8.50为5 a群落的最大值)。箭竹、扁刺蔷薇、山梅花、唐古特忍冬、毛花忍冬 (Lonicera trichosantha Bureau & Franch.)和绣线菊(Spiraea salicifolia L.)是5 a 灌木群落主要优势种。15 a 和23 a 前10种灌木的重要值之和分别为82.21和 80.63,分别有2种和3种植物的重要值 ≥ 10,前者分别有唐古特忍冬(12.78) 和黑水柳(10.57),后者分别有扁刺蔷薇(11.35)、黑水柳(10.95)和大刺茶藨子(10.42)。说明随着植被的演替,个别灌木的生存策略能更好的利用或适应光照和土壤等因子,优先得以生长发育,具体表现出有较大的冠幅和较庞大的根系。Climax 群落的植物生长时间最长,乔木层通过种间自疏效应,呈现出均匀分布的态势。对于灌木层,穿过乔木层林窗的光照成为灌木赖以生存的最重要条件。因此,Climax 灌木群落的重要值之间的差异不大,最大重要值的唐古特忍冬的重要值为8.81<10.00,灌木层的生长状况和分布表现出均质化特征。
-
23 a和Climax的建群种分别为青海云杉和巴山冷杉。从23 a到 Climax,针叶树种的重要值逐渐增加,分别是23 a的23.29和Climax 的64.01。这表明,巴山冷杉林火烧迹地随着植被的正向演替,乔木层由落叶阔叶林逐渐演变成针叶林,并以巴山冷杉为建群种和优势种。
-
依据植物的重要值(附表1,附表2,附表3),判别乔木层、灌木层和草本层的优势种,命名5 a、15 a、23 a和Climax群落分别为:扁刺蔷薇 + 箭竹 + 密生薹草、唐古特忍冬 + 黑水柳 + 密生薹草、青海云杉-红桦 + 扁刺蔷薇 + 密生薹草群落和云杉-巴山冷杉 + 唐古特忍冬 + 密生薹草群落。
-
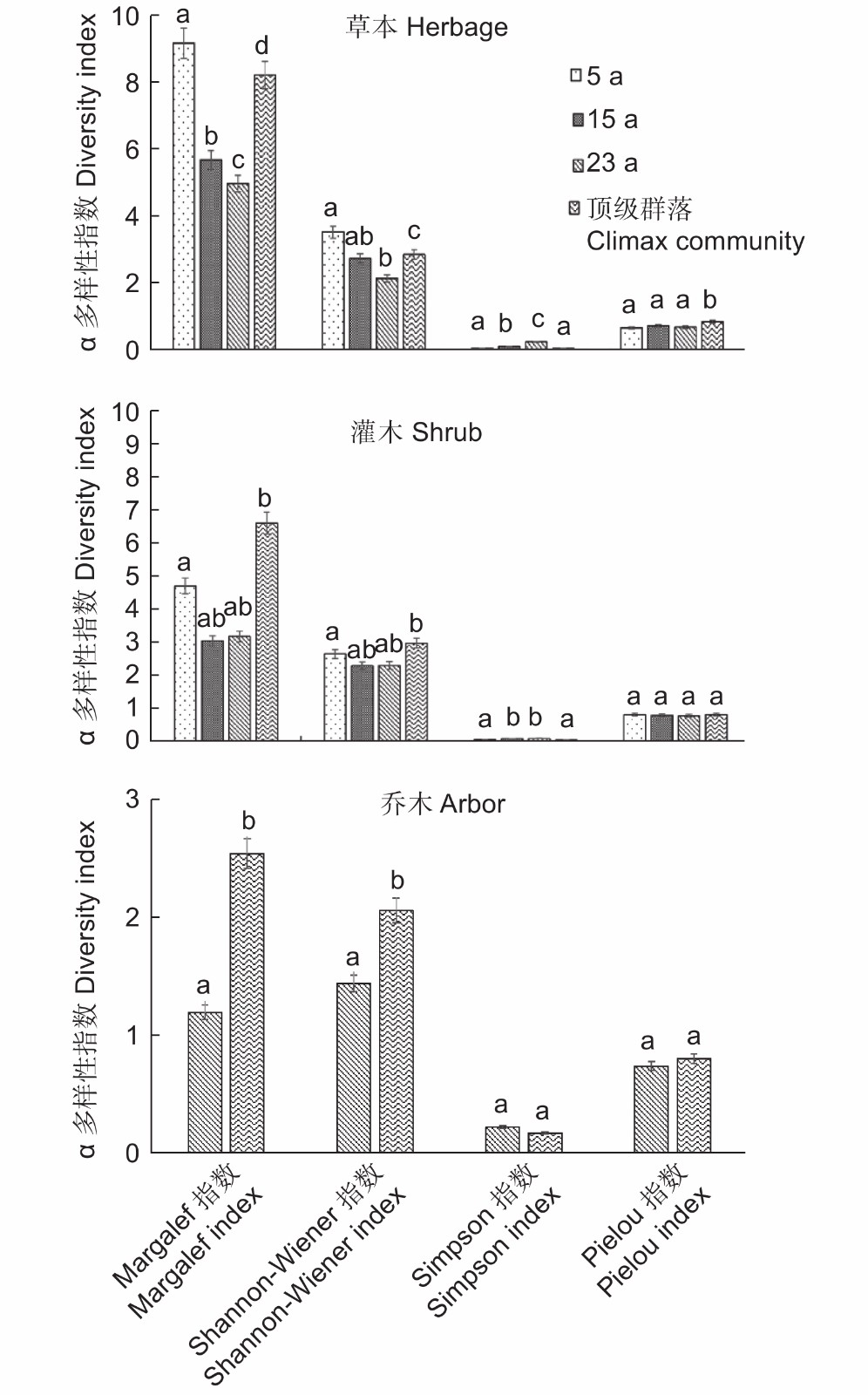
如图1所示,就草本层而言,5 a群落的Margalef指数和Shannon-Wiener指数最大,而23 a群落最小,这与不同演替阶段不同群落的草本物种数变化一致(表2),表明云杉-巴山冷杉林火烧迹地植物群落的次生演替系列中,演替初期草本物种的多样性最大;相反,草本层生物多样性越低的23 a群落,却有着最高的Simpson指数;Climax群落的Pielou指数最大,说明顶级群落草本层物种的分布最均匀。对于灌木层,Margalef指数和Shannon-Wiener指数:Climax>5 a>23 a>15 a;就Simpson指数而言,15 a和23 a高于5 a和Climax;Pielou指数之间没有显著差异,说明在各演替阶段,灌木层在不同的植物群落中都为均匀分布型。乔木层中,Climax的Margalef指数、Shannon-Wiener和Pielou指数都大于23 a群落,而Simpson指数23 a大于Climax群落。这说明,乔木层物种的丰富度和多样性决定于物种的个体总数,优势度和均匀度决定于植物的生长状况和分布方式,5 a群落和Climax群落的Margalef指数和Shannon-Wiener指数较大,相反二者的Pielou指数相对较小。

Figure 1. α diversity indexes of species at different layers in different succession stages
-
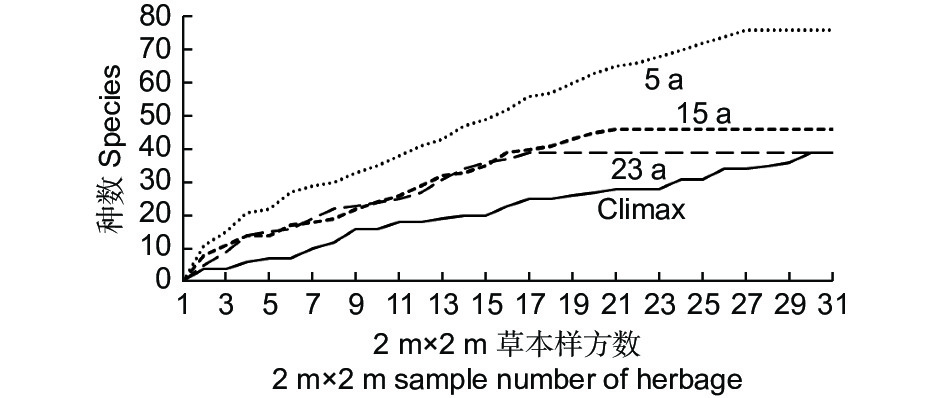
种数-面积曲线表明,随着取样个数的增加,同等样方数的条件下,5 a的物种丰富度始终最大,Climax的物种丰富度最小,15 a和23 a的种面积曲线相差不大。种-面积曲线表明,随着个体数增加,5 a物种丰富度累计速率最大,相应地Climax物种丰富度随个体数增加的累计速率最小(图2)。
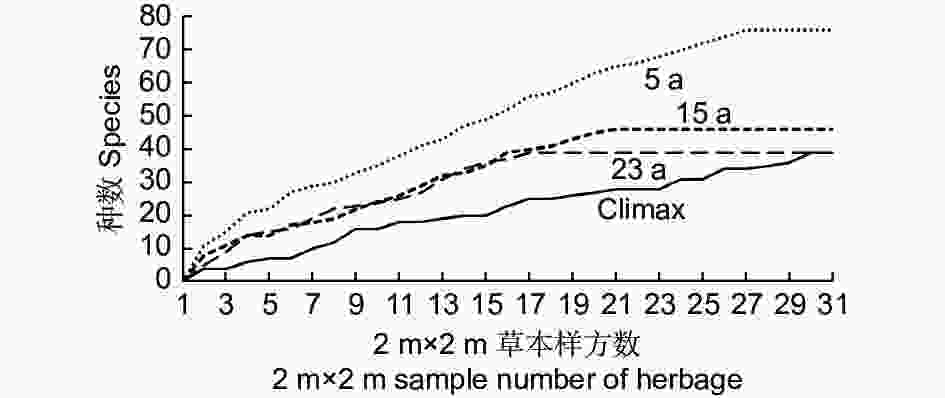
Figure 2. Species – area curve of herbage at different succession stages
同时,从图2反映出不同演替阶段的先锋群落物种的丰富度是有差异的,5 a 和15 a 的种数-面积曲线点与点之间的直线斜率变化较为平缓。相反,23 a 和Climax的种数-面积曲线点与点之间的直线斜率变化较为剧烈。经计算,5 a 群落异质性指数(HI)最小 为0.07,15 a 群落异质指数为0.12,23 a 群落异质指数为0.19,Climax 群落异质指数最大,为0.22。
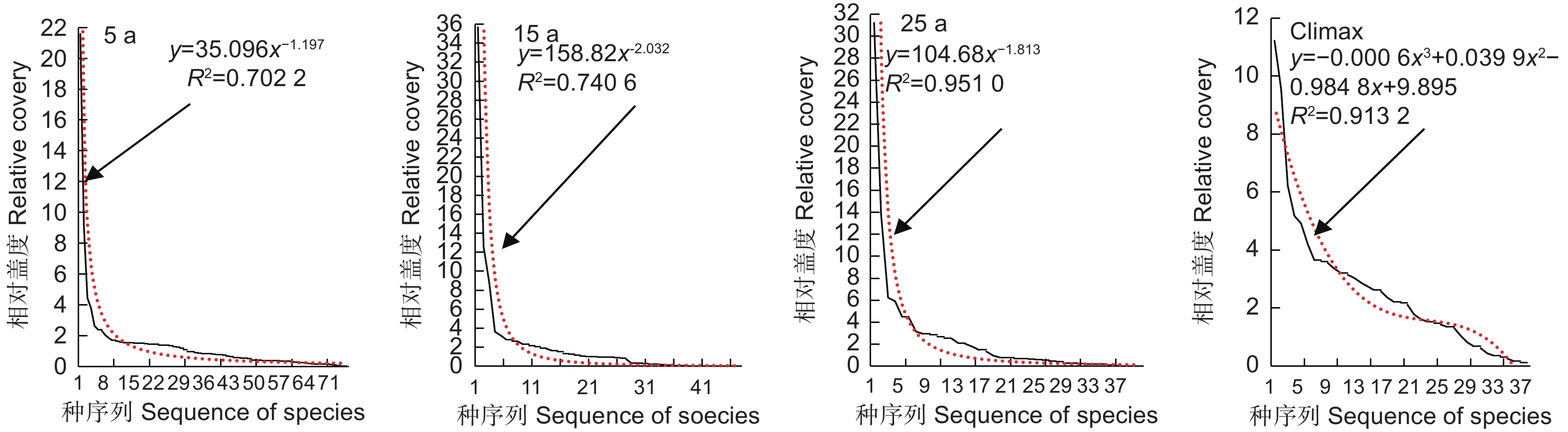
从图3 我们可以看出,5 a、15 a和23 a 3个群落的多度分布均为几何级数分布型,相对盖度分布拟合曲线为幂指数分布;而Climax群落的多度分布呈“S”型分布,多度分布拟合方程为多项式。

Figure 3. Species abundance – sequence diagram
-
从图4 云杉-巴山冷杉林火烧迹地不同演替阶段植物群落间的Wittaker指数形成的树状图可以看出,在火烧迹地发生的演替系列中,对于灌木层,15 a和23 a群落的β多样性最相似;根据β多样性的差异,可把4个演替阶段植物群落直接划分为2类,即可把4个群落的灌木层划分为初建灌木层(5 a)和稳定灌木层(15 a, 24 a, Climax)。对于草本层,随着演替时间越长,不同演替阶段间β多样性值就越高。根据β多样性的差异,可把5 a 和 15 a划分为剧烈变化草本层,意即构建草本群落的种随着小生境的变化而产生较大改变;23 a和Climax群落可划分为稳定草本层,即草本经过生境过滤和对环境的选择性适应,草本群落构建组分处于相对稳定阶段。
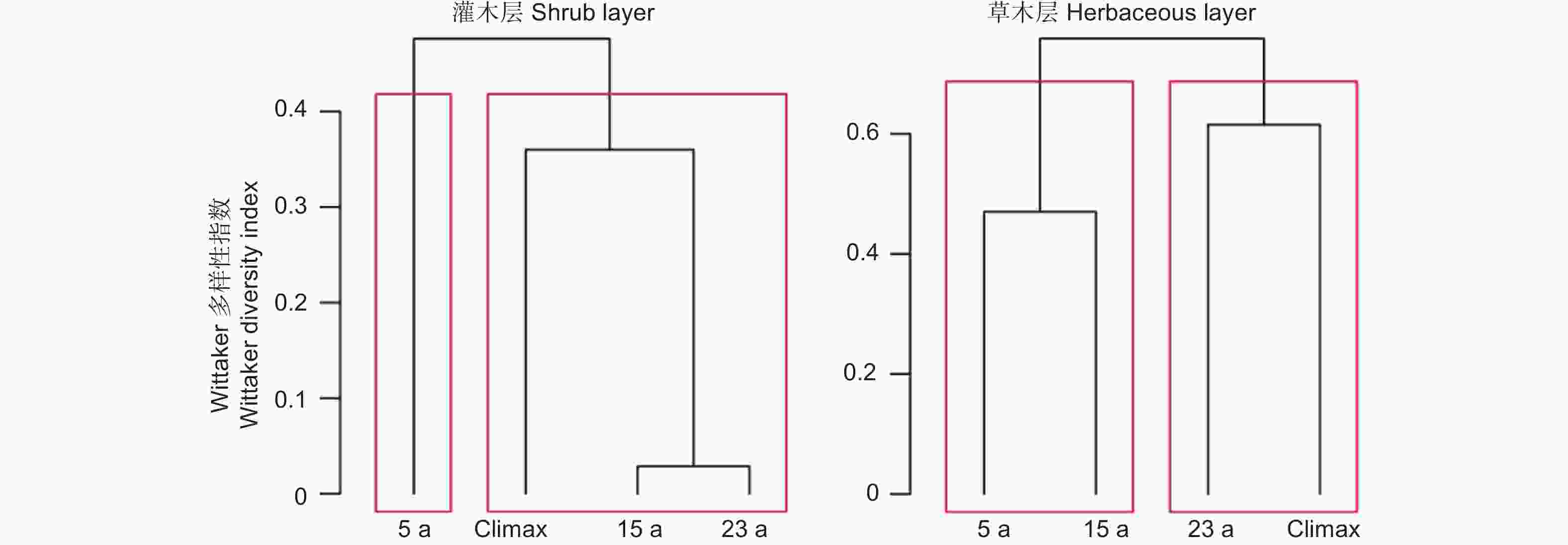
Figure 4. Wittaker index of different plant growth form of Picea - Abies forest burned area at the different succession stage
-
生态学理论研究的主要问题之一就是植物群落的演替[15],植物群落的演替是指植物群落随着时间的推移的过程中,群落不断被另一种群落所代替的有规律的变化。森林重度火后初始,植物尤其是草本物种迅速增多,火烧迹地的植被主要为草本物种,但随着时间的推移,草本物种不仅在数量上明显减少,在物种组成上有很大的变化,盖度也逐渐减少。灌木及乔木物种由于种类较少,在种类及组成上演替初期变化不如草本明显,但是盖度却逐渐增多[16]。本研究中,4个不同演替阶段的植物群落物种丰富度呈递减趋势,Palmquist等的研究表明火干扰可以增加热带稀疏草原物种丰富度,植被恢复初期物种的丰富度增加较快,随后下降并趋于稳定[17]。对于云杉-巴山冷杉林火烧迹地先锋群落草本层物种丰富度随着火后演替时间增加表现出先增加后趋于减少。由于重度林火干扰后,植被恢复时间短,地表裸露,生境恶劣,在光照充足的湿润环境下,具有世代时间短、种子体积小、风传播、光照要求高的r-策略草本物种按照几何指数占用火烧迹地,优先形成草本层,5 a草本层物种及其个体数达到最大,土壤稳定性提高,为根系更为发达的灌木植物创造生长环境,因而灌木后来居上,起到植被恢复的主导作用;随草本层和灌木层的发展,生境得以改善,为乔木生长提供有利的环境,23 a地上植被层次结构逐渐完善,形成明显的乔木层,乔木为群落的主导者,Climax 群落乔木物种数最多,巴山冷杉、岷江冷杉、云杉成为群落的优势种和建群种。随着乔木层的形成,林冠郁闭度逐渐增大,光照限制不利于草本、灌木植物的生长,草本植物的个体数出现急剧下降,这一结果再次验证了余作岳等、王智慧等的研究结果[18-19],即火烧迹地植物群落演替过程中,草本在演替初期阶段,植物的个体数最大,然后随着演替时间增加而逐渐减小;灌木个体数随着演替先增加,伴随着随着乔木层林冠郁闭度增加又减少。草本和灌木在演替过程中都有一个先发展后消退的过程。
对于草地生态系统中植物群落在演替过程中有关物种多样性变化的解释有以下两种理论。第一种理论是,物种多样性随着演替进程的发展而不断增加,也就是物种种类会随演替时间的延长而增加,到演替后期达到最大值[20]。第二种理论是演替的中度干扰理论,指草地生态系统中短命植物和长寿植物共存,物种多样性在演替的中期便会达到最大值[21]。本研究的结果,火烧迹地不同演替阶段草本层和灌木层植物的Margalef丰富度指数、Shannon-Wiener多样性指数的变化规律呈“V型”分布。草本5 a 最大,而灌木Climax最大;Simpson优势度指数在15 a 和23 a达到最大值,遵循中度干扰理论;Pielou均匀度指数在Climax为最大,草本植物在Climax中分布最为均匀,符合演替后期Pielou达到最大值,灌木的Pielou均匀度指数没有差异。在中等或高等强度的林火干扰后,乔木层郁闭度大幅度降低,草本和灌木将得到更多的光资源[22],可用生存空间及资源的大幅度增加,导致许多外来种迁入与定居,同时火烧也促进了土壤养分循环,为植被提供了良好的生长条件,表现出物种多样性的上升[23]。综合分析火烧迹地草本层和灌木层植物多样性的变化特征,说明云杉-巴山冷杉林火烧迹地植物群落演替特征更加接近于森林生态系统,为正向演替。伴随正向演替植物生境进一步改善,环境异质性增加,较耐阴的乔木物种侵入,到 23 a,由于优势种具有更强的竞争优势,乔木树高和冠幅增大,郁闭度也随之增大,草本、灌木进一步减少,草本层多样性、均匀度指数下降,丰富度、优势度指数上升,灌木层物种数减少,优势度指数达到最大,其余指数下降;乔木层各项指数最大,表明群落总丰富度主要受乔木层影响,群落已过渡到了以乔木为主导的阶段且恢复良好,这与王智慧的研究结果一致[19]。
种-面积关系描述的是植物群落中物种数随样地面积的增加而变化的规律[24-26],这甚至已经成为“植物群落生态学中的定律之一”[27]。逻辑斯蒂种—面积关系认为随取样面积的增加, 物种数量具有极限值。物种数先迅速增加, 随后缓慢增加并最终稳定不变, 物种数与面积之间符合逻辑斯蒂曲线[28]。对于云杉属-冷杉属火烧迹地先锋草本层而言,无论5 a、15 a和23 a,还是Climax,种-面积曲线具有共同的特征,即随着样地数的增加,物种数同时增加,当样地数增加到一定值时,草本植物种数不再增加。种-面积关系隐含着复杂的生态过程, 与物种的形成、灭绝和迁移扩散等生态过程密切相关[29-31],生物群落的种-面积关系主要与种-多度分布和物种的空间分布有关[32]。在均质生境中,样方所包含的个体数与取样面积成正比[33],物种数随取样面积的增加而增加[34],而取样面积所包含的物种数主要取决于所有个体在不同物种之间的分配,物种在空间分布上聚集程度越高,小面积中可观察的物种会越少,物种数随面积增加的速率会下降[35]。本研究结果与He和Legendre一致, 即物种在空间分布上聚集程度越高, 小面积中可观察的物种会越少, 物种数随面积增加的速率会下降, 即具有较小的斜率。样地物种数分布相对均匀时,所需最小样方数量较小[36]。从5 a 到Climax,草本群落异质化程度逐渐增加是合乎情理的,因为云杉-巴山冷杉林火烧迹地演替初期的草本群落主要是通过种子扩散决定的,草本种子由于多年的散布较均匀地分布在火烧迹地内各个地点。再者,优势种的生态位预占特性也将使种子种类组成的差异进一步缩小。随着以后的群落演替发展,异质化程度将随着灌木和乔木的侵入及定殖趋于增加。
物种多度分布格局分析对于理解植物群落物种多样性的形成和维持机制具有重要意义[37]。多度可以反映出一个物种在植物群落中占用资源的能力,不同的植物群落具有不同的多度分布格局。物种多度分布格局是多个物种不断地相互作用、相互影响的结果,通过研究多度分布格局可揭示植物群落的组成,进而了解群落中不同物种间的关系和作用机制[38]。由于自然群落结构的复杂性,不同物种多度分布模型表示的生态学意义更能够真实有效地反映群落的内在特征[39]。青藏高原东北边缘云杉-巴山冷杉林火烧迹地先锋植物群落全部物种多度分布随演替阶段发生了明显的变化,导致与其符合的多度-种序列图也随之改变,同时也改变了其群落构建的生态学过程(图3)。5 a、15 a和23 a群落多度分布趋势线拟合呈幂指数分布(R2 > 0.95),拟合效果特佳,这可能是火烧迹地环境条件有利于草本植物生长,减弱了物种间竞争作用,生态位确定性物种在群落构建过程中占据主要地位;但同一群落各物种相似的生态适应性会导致产生较高的生态位重叠,尤其对于物种数量多且差异较小、物种分布比较均匀的群落来说更为显著[40-41]。对于Climax,多度分布趋势线拟合为“S”型,这意味着随着演替的进行,Climax群落变得越来越复杂,“中间类型”物种占据了群落的绝大部分,属于演替晚期群落[42]。
β多样性作为植物多样性的重要测度指标, 通过反映群落物种组成沿某一环境梯度的周转速率[43], 不仅能够揭示不同环境梯度下生境的多样性, 而且能够反映不同时空尺度之间的物种组成差异性[44],从而一定程度上可以揭示植物群落沿这个环境梯度的生态过程[45]。研究植物群落β-多样性的分布格局,才能更好地认识物种与环境的关系,还能够了解不同生态过程在植物群落形成和维持中的相对作用,对群落构建过程进行深入理解[46]。生态位理论认为植物群落β-多样性的差异是环境差异造成的[47],随着环境差异的增加,群落之间物种组成相似性会降低,β-多样性则升高[48-49]。就青藏高原东北边缘的云杉属-冷杉属林火烧迹地灌木层而言,火烧后演替5 a与演替15 a、25 a和Climax之间的β-多样性指数差异最大,植物的相似性最小;对于草本层,5 a和15 a之间、23 a和Climax之间的的β-多样性指数值相对接近,植物群落构成物种相似性高。研究表明随着火后演替时间的增加,生境光照、土壤的理化性质和植物群落自身的特性都会在不同程度上对植物群落β-多样性及其组分产生影响。因此在今后的研究中,应当在已有研究的基础之上结合生物与非生物因素,更加全面地对青藏高原东北边缘的云杉属-冷杉属林火烧迹地植物群落β-多样性及影响因素进行探究。
-
在云杉-巴山冷杉林火烧迹地,不同演替阶段的植物群落物种丰富度呈递减趋势,群落中草本层和灌木层物种的Margalef指数、Shannon-Wiener指数的变化规律呈先减少后增加分布,Simpson指数在中前期达到最大值,遵循中度干扰理论;Pielou指数在Climax达到最大值,灌木的Pielou均匀度指数没有差异。从演替开始到顶级群落,植物群落随着灌木和乔木的侵入及定殖,群落异质化程度逐渐增加。演替前期和中期,生态位确定性物种在群落构建过程中占据主要地位;对于顶级群落,“中间类型”的物种占据了群落的绝大部分。随着火后演替时间越长,不同群落间β-多样性指数越加接近,青藏高原东北边缘云杉-巴山冷杉林火烧迹地植物群落演替为正向演替,群落的演替方向为灌木林——落叶阔叶林——针叶林——云杉-巴山冷杉林。
演替阶段
Succession stages物种
Species株数
Individual高
Height/cm盖度/%
Coverage重要值
Important value5 a 前10种 Top 10 plant species 密生薹草 Carex crebra 1 028±51 42.48±10.32 47.52±3.09 12.97±2.51 总状橐吾 Ligularia botryodes 284±74 21.76±4.55 32.00±11.18 4.85±0.36 婆婆纳 Veronica polita 427±33 5.59±3.96 4.19±2.50 4.30±0.08 草地风毛菊 Saussurea amara 119±87 34.50±8.71 35.60±9.48 3.69±0.25 白叶蒿 Artemisia leucophylla 255±64 40.00±3.08 5.80±5.06 3.23±0.73 车前 Plantago asiatica 26±7 13.00±0.14 46.00±7.08 3.18±024 平车前 Plantago depressa 151±12 10.82±0.47 14.55±4.42 2.42±0.55 中华蹄盖蕨 Athyrium sinense 75±35 34.33±7.78 12.89±0.97 1.91±0.06 糙苏 Phlomis umbrosa 17±10 120.00±19.33 2.13±0.85 1.79±0.12 椭圆叶花锚 Halenia elliptica 121±3 18.50±2.06 7.12±1.26 1.79±0.30 15 a 前10种 Top 10 plant species 密生薹草 Carex crebra 574±88 23.35±5.06 55.20±5.01 17.18±14.08 高山冷蕨 Cystopteris montana 301±117 16.11±4.53 19.72±2.38 7.58±0.36 唐古拉婆婆纳 Veronica vandellioides 219±42 8.88±1.23 20.51±0.54 6.46±1.22 野草莓 Fragaria vesca 289±131 7.00±0.65 15.41±3.29 6.36±0.05 平车前 Plantago depressa 103±26 6.29±2.30 22.39±55.54 5.29±4.45 猪殃殃 Galium spurium 337±125 17.25±5.85 2.00±0.06 5.03±0.01 艾 Artemisia argyi 158±74 24.18±6.33 10.71±5.85 4.62±0.17 总状橐吾 Ligularia botryodes 163±15 15.76±2.32 11.39±9.28 4.48±2.38 腺点柔毛蓼 Polygonum sparsipilosum 124±102 8.50±1.08 9.00±0.85 3.34±0.08 宽叶羌活 Notopterygium franchetii 2±2 84.00±12.31 0.04±0.04 3.13±3.13 23 a 前10种 Top 10 plant species 密生薹草 Carex crebra 881±206 47.00±21.33 49.35±13.62 26.05±5.92 高山冷蕨 Cystopteris montana 472±162 15.42±4.86 21.85±1.28 12.68±2.35 藜 Chenopodium album 87±54 42.00±6.39 9.36±3.47 4.71±0.26 猪殃殃 Galium spurium 108±30 19.85±12.34 9.86±2.06 4.45±0.84 甘青剪股颖 Agrostis hugoniana 41±17 28.50±11.38 6.94±2.89 3.03±0.06 莛子藨 Triosteum pinnatifidum 8±6 58.00±23.41 4.53±0.88 2.94±0.36 箭叶橐吾 Ligularia sagitta 9±4 77.86±10.65 1.25±0.06 2.89±0.15 林地早熟禾 Poa nemoralis 16±3 52.00±6.35 4.05±1.25 2.77±0.12 总状橐吾 Ligularia botryodes 42±18 12.35±0.52 7.12±1.37 2.57±0.06 白叶蒿 Artemisia leucophylla 42±37 38.50±4.36 2.85±0.51 2.50±0.27 顶级群落 Climax community 前10种 Top 10 plant species 密生薹草 Carex crebra 248±112 19.59±4.48 18.67±3.98 14.58±3.06 高山冷蕨 Cystopteris montana 136±57 19.92±3.52 12.13±1.65 9.42±1.77 高山露珠草 Circaea alpina 180±28 6.32±1.38 7.94±0.96 7.74±1.03 长梗金腰 Chrysosplenium axillare 132±53 3.33±0.07 5.50±1.35 5.45±0.41 总状橐吾 Ligularia botryodes 32±8 11.65±5.23 6.99±0.76 4.39±0.33 五福花 Adoxa moschatellina 16±6 34.50±4.40 3.17±0.06 4.14±0.01 东方草莓 Fragaria orientalis 88±15 4.82±0.12 4.10±0.17 3.99±0.21 疏穗野荞麦 Fagopyrum caudatum 8±1 42.38±16.14 0.24±0.01 3.34±0.06 玉竹 Polygonatum odoratum 8±5 34.55±1.18 1.26±1.02 3.19±2.74 茖葱 Allium victorialis 60±8 11.5±0.98 1.80±0.04 2.92±0.05 Table 1. Important value of herbaceous stratum
演替阶段
Succession stages物种
Species株数
Individual高
Height/cm盖度/%
Coverage重要值
Important value5 a 前10种 Top 10 plant species 箭竹 Fargesia spathacea 22±13 83.32±26.56 14.70±4.84 8.50±3.51 扁刺蔷薇 Rosa sweginzowii 12±6 112.38±47.86 30.10±12.63 8.08±2.37 山梅花 Philadelphus incanus 7±2 89.4±21.05 38.91±10.14 7.10±2.62 唐古特忍冬 Lonicera tangutica 1±1 93.00±10.85 46.80±3.85 6.50±1.32 毛花忍冬 Lonicera trichosantha 1±1 122.25±21.74 36.30±4.78 6.31±0.78 绣线菊 Spiraea salicifolia 18±7 11.45±4.85 16.40±0.66 6.30±0.69 托柄菝葜 Smilax discotis 17±11 46.29±32.63 7.90±0.12 5.81±0.45 黑水柳 Salix heishuiensis 9±5 75.29±46.39 20.50±3.21 5.67±0.38 楤木 Aralia elata 7±5 85.88±22.45 17.60±5.35 5.25±0.22 美丽悬钩子 Rubus amabilis 5±3 73.92±8.15 13.30±2.08 3.98±0.08 15 a 前10种 Top 10 plant species 唐古特忍冬Lonicera tangutica 28±11 112.58±10.88 78.05±5.85 12.78±2.86 黑水柳Salix heishuiensis 27±5 79.72±41.35 59.26±13.32 10.57±3.25 灰栒子Cotoneaster acutifolius 1±1 540.00±117.25 12.17±0.82 9.95±1.78 箭竹 Fargesia spathacea Franch 38±21 68.31±5.38 18.23±1.85 9.01±1.32 大刺茶藨子Ribes alpestre var. giganteum 25±6 46.42±13.50 16.32±2.74 7.89±1.82 山梅花hiladelphus incanus 26±11 40.94±25.74 15.43±0.91 7.63±1.06 峨眉蔷薇Rosa omeiensis 15±3 86.08±32.98 14.26±1.76 7.36±0.98 扁刺蔷薇Rosa sweginzowii 9±5 124.07±30.00 13.39±2.05 6.65±0.93 毛花忍冬 Lonicera trichosantha 8±8 143.00±55.06 10.23±1.51 5.53±1.03 小叶蔷薇Rosa willmottiae 3±1 139.15±27.35 5.26±0.16 4.84±0.35 23 a 前10种 Top 10 plant species 扁刺蔷薇Rosa sweginzowii 51±13 108.56±27.56 47.12±8.85 11.35±2.36 黑水柳Salix heishuiensis 27±5 335.00±89.35 12.20±7.98 10.95±4.41 大刺茶藨子Ribes alpestre var. giganteum 23±19 126.66±88.33 19.46±3.55 10.42±2.03 山梅花hiladelphus incanus 19±8 178.95±43.52 16.61±4.17 9.44±1.92 唐古特忍冬Lonicera tangutica 2±1 214.00±70.00 25.16±5.82 9.40±1.53 陕甘花楸Sorbus koehneana 1±1 468.00±58.65 16.88±14.33 9.25±2.35 峨眉蔷薇Rosa omeiensis 21±14 126.75±14.71 5.77±2.36 6.23±1.84 毛花忍冬 Lonicera trichosantha 11±3 348.06±110.55 1.8±0.09 5.52±0.03 箭竹Fargesia spathacea 21±7 56.38±4.85 3.34±0.12 4.89±0.06 小叶蔷薇Rosa willmottiae 5±5 188.05±100.08 2.26±0.52 3.19±3.19 顶级群落 Climax 前10种 Top 10 plant species 唐古特忍冬 Lonicera tangutica 31±9 62.6±38.58 13.87±2.85 8.81±2.81 大刺茶藨子 Ribes alpestre var. giganteum 6±2 106.97±27.41 6.32±1.32 5.74±1.06 山梅花 Philadelphus incanus 4±1 130.86±59.65 5.48±2.63 5.11±1.35 金银忍冬 Lonicera maackii 2±1 218.55±110.33 4.62±4.08 5.1±2.38 六道木 Zabelia biflora 1±1 170.25±89.36 5.72±2.98 5.05±5.05 毛花忍冬 Lonicera trichosantha 11±5 77.38±12.35 2.82±1.02 4.47±0.87 箭竹 Fargesia spathacea Franch 17±7 63.12±41.76 0.44±0.32 4.18±1.63 葱皮忍冬 Lonicera ferdinandi 3±1 173.87±62.31 2.7±0.03 3.79±1.32 楤木 Aralia elata 1±1 151.83±74.35 3.59±0.25 3.68±3.68 美丽悬钩子 Rubus amabilis 14±6 55.85±12.36 0.45±0.04 3.53±0.98 Table 2. Important value of shrub stratum
Plant Community Succession in the Burned Area of Picea asperata- Abies fargesii Forest on the Northeastern Edge of the Qinghai-Tibetan Plateau
- Received Date: 2022-09-06
- Accepted Date: 2022-09-20
- Available Online: 2023-04-20
Abstract:



 DownLoad:
DownLoad: